
不同光照条件对2个品种(系)坛紫菜光合作用、呼吸作用的影响
2018-12-19 09:05王怡娟应苗苗周庆澔陈思航许曹鲁
江苏农业科学 2018年22期
权 伟, 王怡娟, 应苗苗, 周庆澔, 陈思航, 许曹鲁
(1.温州科技职业学院/浙江省温州市农林渔生态系统增汇减排重点实验室,浙江温州325006;2.浙江省温州市洞头区水产科技推广站,浙江温州325700)
坛紫菜(Porphyrahaitanensis)广泛栽培于我国的浙、闽、粤3省沿海地区,其产量占全国紫菜产量的 75% 左右[1],光合作用与呼吸作用中溶解氧(dissolved oxygen,DO)、溶解无机碳(dissolved inorganic carbon,DIC)的供应及利用是坛紫菜养殖过程中的基本理论问题之一。DO通常是指通过大气交换或经生物、化学反应溶解于海水中的氧气,它是海洋生命活动不可缺少的重要物质,也是衡量海水环境质量的标准之一[2]。DIC是海洋碳系统中最主要的存在形态之一,也是海洋碳循环和全球碳循环研究中极其重要的部分[3-4]。坛紫菜在养殖过程中随着潮汐变化会经历反复干出和沉水循环,沉水阶段通过光合作用利用水体中的DIC,释放DO,通过呼吸作用消耗水体中的DO,释放DIC。
包括坛紫菜在内的大型海藻是近岸海洋生态的主要初级生产者,其强大的光合固碳能力显著增加了海洋碳汇强度[5-7]。大型海藻养殖能够降低水体无机碳浓度,促进大气CO2向海水转移,进而降低大气CO2浓度。同时,伴随养殖大型海藻的收获,形成了一个“可移出的碳汇”。所以,大力发展近海海藻养殖具有重要的碳汇生态意义及社会意义。
目前对坛紫菜光合生理的研究主要集中在生态环境因子(营养盐、CO2浓度升高、紫外辐射、温度、pH值)对光合作用的影响[8-13],坛紫菜在不同的光照、黑暗持续时间及不同光照度条件下,产氧、固碳速率会呈现怎样的变化很少涉及,开展相关研究可为坛紫菜生理生态、固碳机理的研究及其人工栽培提供依据。
1 材料与方法
1.1 材料
养殖试验坛紫菜(三水)及海水采集于2014年12月,取自素有“浙江省紫菜之乡”美誉的浙江省温州市洞头区。洞头自20世纪60年代开始试养坛紫菜,至今已有50多年养殖历史,截至2017年,洞头紫菜养殖面积2 000多hm2,堪称“海上田园”。洞头本地菜为洞头本地传统养殖品系的俗称,浙东1号从宁波大学引进。采集的坛紫菜用原海水在实验室内暂养约12 h,养殖试验海水基本理化参数见表1。
1.2 方法
养殖试验于2014年12月13—14日在温州科技职业学院园林与水利工程学院组培室进行。将完整的坛紫菜放入盛有250 mL海水的透明广口玻璃瓶内,玻璃瓶经高温高压灭菌,从培养瓶放入光照或黑暗环境下开始计时。
不同光照时间养殖试验的光照度为2 800 lx,日光灯为光源,温度23.1 ℃,黑暗养殖时温度为20.5 ℃,分别于处理后1、2、3.5、5、6、8、10、12 h时用英国Aquaread Ap-2000水质分析仪测定DO含量;光照、黑暗养殖试验分别设置了3个时间梯度处理,L2(光照2 h)、L6(光照6 h)、L12(光照12 h),D2(黑暗2 h)、D6(黑暗6 h)、D12(黑暗12 h),处理时间截止后立即捞出坛紫菜,用Whatman GF/F 0.45 μm玻璃纤维滤膜抽滤,滤液DIC含量用岛津TOC-LCPH总有机碳分析仪测定;不同光照度试验另外同时设置了1500、2000、3800lx3个梯度的光照度,光照6 h后测定DO、DIC含量。所有处理同时设置空白对照,处理和对照均为3个重复。养殖试验结束后,将养殖后捞出的坛紫菜放至80 ℃烘箱内,烘24 h至恒质量,称干质量DW。

表1 养殖试验海水基本理化参数背景值
光合产氧速率、呼吸耗氧速率指单位质量(干质量)坛紫菜在单位时间内引起水体DO含量的变化。用下式计算:
RO[mg/(kg·h)]=(CT-C0)×V/(DW·T)。
式中:CT是试验结束时有坛紫菜的光合作用瓶DO的含量,mg/L;C0是空白对照瓶中(无坛紫菜)DO的含量,mg/L;V是养殖用水体积,L;DW是试验坛紫菜的干质量,kg;T是试验处理时间,h。
光合固碳速率、呼吸作用释放DIC速率指单位质量(干质量)坛紫菜在单位时间内引起处理水体DIC含量的变化。用下式计算:
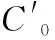
光合熵(photosynthetic quotient,PQ)指光合作用净产氧量与光合作用固碳量的比值(摩尔比),通过下式计算:PQ=产氧量/碳吸收量=Δ[O2]/Δ[DIC]。
呼吸熵(respiration quotient,RQ)指呼吸作用释放DIC量与呼吸作用耗氧量的比值(摩尔比),通过下式计算:RQ=碳释放量/耗氧量=Δ[DIC]/Δ[O2]。
1.3 统计分析
采用Excel 2003对文中数据进行计算及作图处理。所有统计分析用SPSS 17.0统计软件,不同品种(系)坛紫菜各指标的差异性分析用配对样本t检验方法,不同处理时间、不同光照度间各指标的差异性分析用单因素方差分析方法。
2 结果与分析
2.1 不同处理下2个品种(系)坛紫菜的产氧、耗氧速率
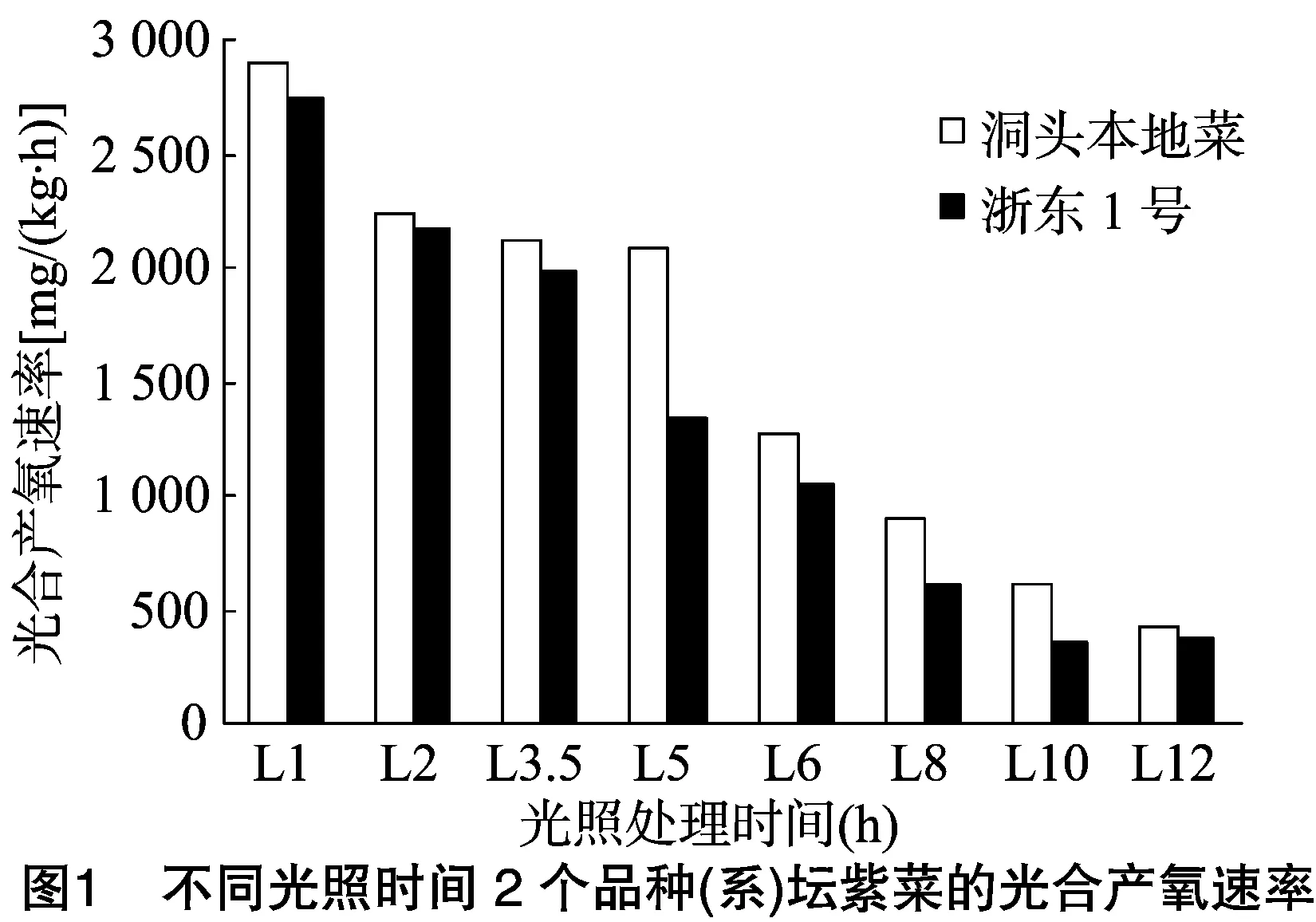
2.1.1不同光照时间对光合产氧速率的影响 由图1可知,洞头本地菜、浙东1号光合产氧速率在12 h内随着光照时间的延长均呈下降趋势,不同处理时间光合产氧速率差异极显著(P<0.01)。洞头本地菜光合产氧速率由2 897.52 mg/(kg·h)降至422.61 mg/(kg·h),浙东1号由2 746.93 mg/(kg·h)降至368.62 mg/(kg·h)。相同光照时间下,洞头本地菜光合产氧速率均高于浙东1号。
2.1.2 不同光照度对光合产氧速率的影响 由图2可知,洞头本地菜、浙东1号光合产氧速率随着光照度的增加先升高后降低,均在光照度2 800 lx条件下最高,分别达1 270.49、1 043.97 mg/(kg·h),在3 800 lx条件下最低,分别降至517.48、284.23 mg/(kg·h),不同光照度间光合产氧速率差异极显著(P<0.01)。相同光照度条件下,洞头本地菜光合产氧速率均高于浙东1号,差异显著(P<0.05)。

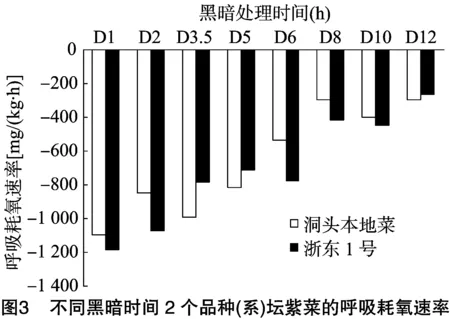
2.1.3 不同黑暗时间对呼吸耗氧速率的影响 由图3可知,洞头本地菜、浙东1号呼吸耗氧速率(用其绝对值比较)在 12 h 内随着黑暗时间的延长均呈下降趋势,不同处理时间呼吸耗氧速率差异极显著(P<0.01)。洞头本地菜呼吸耗氧速率由1 092.55 mg/(kg·h)降至296.66 mg/(kg·h),浙东1号由1 182.35mg/(kg·h)降至263.80 mg/(kg·h)。相同黑暗时间下,洞头本地菜与浙东1号之间差异不显著。
2.2 不同处理下2个品种(系)坛紫菜的固碳、释碳速率
2.2.1 不同光照时间对光合固碳速率的影响 由图4可知,洞头本地菜、浙东1号光合固碳速率随着光照时间的延长均呈先上升后下降的趋势,不同光照时间光合固碳速率差异极显著(P<0.01)。在L6时最高,洞头本地菜光合固碳速率达1 042.62 mg/(kg·h),浙东1号达976.35mg/(kg·h)。相同光照时间处理下,洞头本地菜光合固碳速率均高于浙东1号,但差异不显著。

2.2.2 不同光照度对光合固碳速率的影响 由图5可知,洞头本地菜、浙东1号光合固碳速率随着光照度的增加均呈先上升后下降的趋势,不同光照度间光合固碳速率差异极显著(P<0.01)。在2 800 lx时最高,洞头本地菜光合固碳速率达1 042.62 mg/(kg·h),浙东1号达976.35mg/(kg·h)。相同光照度条件下,洞头本地菜光合固碳速率均高于浙东1号,但差异不显著 (P>0.05)。

2.2.3 不同黑暗时间对呼吸作用DIC释放速率的影响 由图6可知,洞头本地菜、浙东1号呼吸释碳速率(用其绝对值比较)随着黑暗时间的延长逐渐降低,不同黑暗时间呼吸作用释放DIC速率差异极显著(P<0.01)。在D2时最高,洞头本地菜呼吸作用释放DIC速率达455.32 mg/(kg·h),浙东1号达404.97 mg/(kg·h)。相同黑暗时间处理下,洞头本地菜呼吸释碳速率均高于浙东1号 (P>0.05)。

2.3 不同处理下2个品种(系)坛紫菜的光合熵、呼吸熵
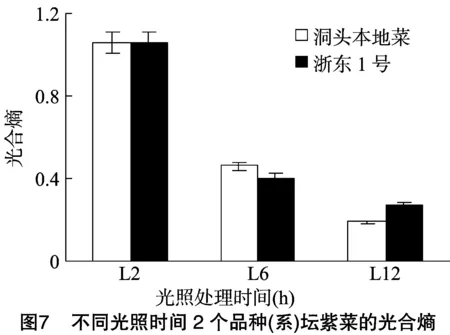
2.3.1 不同光照时间对光合熵的影响 由图7可知,洞头本地菜、浙东1号光合熵随着光照时间的延长逐渐降低,均在L2时达到最高,浙东1号最高,为1.06,不同光照时间光合熵差异极显著(P<0.01)。L2、L12相同光照时间处理下洞头本地菜光合熵均低于浙东1号,L6处理下洞头本地菜光合熵高于浙东1号 (P>0.05)。
2.3.2 不同光照度对光合熵的影响 由图8可知,洞头本地菜、浙东1号光合熵随着光照度的增加,光合熵先增加后降低,不同光照度间光合熵差异极显著(P<0.01)。2个品种(系)坛紫菜光合熵在2 000 lx时最高,洞头本地菜达0.54,浙东1号达0.44。3 800 lx时最低,光合熵均为0.19。相同光照度条件下,洞头本地菜光合熵均高于浙东1号 (P>0.05)。

2.3.3 不同黑暗时间对呼吸熵的影响 由图9可知,洞头本地菜、浙东1号呼吸熵随着光照时间的延长呈降低趋势,均在D2时最高,洞头本地菜达1.43,浙东1号达1.01,不同黑暗时间呼吸熵差异极显著(P<0.01)。在相同黑暗时间处理下,洞头本地菜呼吸熵均高于浙东1号 (P>0.05)。
3 讨论
3.1 坛紫菜在光照条件下的光合作用特性
海水中的DIC是海洋中自养生物合成有机物质的碳素来源[14],海水中DIC的主要成分包括海水中溶解的CO2、HCO3-以及CO32-,其中HCO3-占DIC的85%以上,CO32-占9%左右,其余为溶解的H2CO3和CO2[4]。大型海藻通过利用海水中溶解的无机碳进行光合作用,不但利用海水中游离CO2作为光合碳源,而且还可以利用海水中存在的大量HCO3-[15-16]。水体中游离的CO2分子可以自由透过细胞质膜,HCO3-经质膜外碳酸酐酶(CA)催化水解成游离CO2后以CO2分子形式扩散进入细胞膜[17]。大型海藻养殖能够显著改变海水中无机碳的含量,明显提高海水的DO水平[18]。

光照是光合作用的能源,因此光照对藻类的生长影响很大。坛紫菜等红藻的光合色素和光能转化,除了一般植物具有的叶绿素a外,还有重要的捕光色素——藻胆蛋白[19]。叶绿素在光合作用中起吸收和传递光量子的作用,其含量的大小可以反映植物叶片光合作用功能的强弱。光照影响紫菜体内色素含量[20],鼠尾藻异形叶的叶绿素a含量与最大净光合速率(Pn,max)呈正相关[21],但未发现条斑紫菜光合色素含量与光合效率的相关性[22]。
在本研究中,随着光照时间的延长,光合固碳速率呈先上升后下降的趋势(在L6时最高),这可能与大型海藻对DIC的利用机制有关。在光照条件养殖初期,坛紫菜的光合作用首先吸收了能够被直接利用的CO2,随着水体CO2含量的降低,快速激发了坛紫菜利用HCO3-的浓缩机能,水体中存在的大量HCO3-被吸收利用,使得光照初期光合固碳速率升高。坛紫菜利用HCO3-作为光合作用时的碳源、胞外CA催化的HCO3-脱水作用、其低的米氏常数(Km)值及低的无机碳补偿点表明坛紫菜具有浓缩碳的作用[23]。随着光合作用的进行,水中的无机碳含量降低,从而导致水体CO2分压降低,大气向水体扩散CO2,但是水体与大气交换有一个滞后作用,培养液中的碳源已难以满足其光合作用的需求,致使光照后期光合固碳速率下降。充足的碳源能提高坛紫菜光合作用强度,海水搅动较大的海域能够及时补充光合作用消耗的DIC,可以减弱水体DIC浓度降低对光合作用产生的抑制,促进坛紫菜的生长。本研究表明,随着光照时间的延长,2个品种(系)坛紫菜光合产氧速率均呈下降趋势,可能与水体中溶解无机碳浓度不断下降及大气向水体扩散CO2的补给不及时有关。王云霞研究发现,紫菜叶状体培养过程中随着发育现象的出现,各组织的光合放氧速率均呈现降低的趋势[24],这与本研究结论相一致。
在本研究中,在不同的光照度下,2个品种(系)坛紫菜光合产氧速率、光合固碳速率均在光照度2 800 lx条件下最高。光照度大于2 800 lx后,固碳速率随着光照度升高而降低的原因可能是藻体受到损伤抑制了光合作用。研究光照度对坛紫菜光合呼吸作用的影响时,还必须结合考虑培养时的温度、营养和碳源,在营养不足、碳源缺乏的情况下,片面地提高光照度可能造成生理作用受抑制。坛紫菜丝状体生长在 3 000 lx 时达到最高效率[13],严兴洪等发现,坛紫菜叶状体细胞的适宜发育光照度为2 000~3 000 lx,过高或过低的光照度均会抑制苗假根形成和生长[25]。条斑紫菜叶状体光系统(PS)Ⅰ活性随温度和光照度的升高而呈现先升高后降低的趋势,光照度为2 320 lx时具有最高的PSⅠ活性[26],上述研究结论的最适光照度与本研究结论较为接近。
不同品种(系)大型海藻的光合呼吸生理作用有一定的差异性,随着光照度从20 μmol/(m2·s)提高到 1 200 μmol/(m2·s),不同品种(系)条斑紫菜光合效率的下降幅度有一定差异[22];黄瑶的研究表明,在未充CO2的试验条件下,鼠尾藻比石莼和海黍子总无机碳 DIC吸收速率高,最高达到423.36 μmol/(g·h),添加营养盐对大型藻类的总无机碳吸收速率 RDIC影响差异不显著,添加营养盐能够提高龙须菜的光合产氧速率,充CO2对石莼、鼠尾藻和海黍子RDIC的影响差异显著[18];蔡继晗等在浙江省苍南县沿浦湾海域养殖试验表明,洞头本地菜单养殖周期产量高于浙东1号,产量高表明其净光合效率较高[27]。本研究表明,相同光照时间、光照度处理下,洞头本地菜光合产氧速率、光合固碳速率均高于浙东1号,这与蔡继晗等研究结论[27]相吻合。
光合熵(PQ)是反映大型藻类光合生理特性的重要指标,光合熵较为敏感,可以作为衡量大型藻类固碳能力的动态参数,不同类型大型藻类PQ与其无机碳利用机制有关,试验大型藻类的光合熵在0.36~1.18[18]。本试验中,随着光照时间的延长,2个品种(系)坛紫菜光合熵逐渐降低,这可能是由于随着光照时间的延长,水体中DIC逐渐降低,坛紫菜光合利用DIC的难度增强,须要消耗更多的能量获取DIC,使得光呼吸作用增强,同时水体中逐渐升高的DO含量为坛紫菜的光呼吸作用提供了充足的氧气,消耗了更多的DO,致使产氧量下降、PQ降低。本研究表明,在不同光照度下,2个品种(系)坛紫菜光合熵在2 000 lx时最高,此时产氧效率较高,有利于水体DO含量的升高。2个品种(系)坛紫菜光合熵在较高光照度3 800 lx时最低,高光照度可能抑制了坛紫菜的光合作用,激发了光呼吸作用,致使产氧效率降低。相同光照度条件下,洞头本地菜光合熵高于浙东1号,表明洞头本地菜产氧效率优于浙东1号,洞头本地菜固碳效率弱于浙东1号。
3.2 坛紫菜在黑暗条件下的呼吸作用特性
呼吸作用是植物体内非常重要的生理过程,为植物的生命活动提供大部分能量,但容易受到外界环境的影响。藻体的光合作用与呼吸作用均受到高盐度海水的显著影响,随着盐度的增加,光合放氧率逐渐降低,呼吸耗氧率也逐渐降低[28]。
本试验中,随着黑暗时间的延长,2个品种(系)的坛紫菜呼吸耗氧速率呈下降趋势。这可能是由于随着光照时间的延长、水体中DO含量的降低,有氧呼吸受到抑制,通过酶的催化作用而发生无氧呼吸,坛紫菜共同发生有氧呼吸与无氧呼吸,但由于无氧呼吸释放的ATP较少,同时由于DO含量的降低对有氧呼吸的抑制,综合导致暗呼吸作用强度将低,呼吸耗氧速率逐渐下降。
RQ可以作为判断呼吸底物的类型和供氧情况的指标之一[29]。本研究表明,随着黑暗时间的延长,2个品系的坛紫菜呼吸熵呈下降趋势。D2时,2个品种(系)的坛紫菜呼吸熵均>1,表明此时呼吸作用主要的呼吸底物为糖类,同时进行了有氧呼吸与无氧呼吸,随着呼吸作用时间延长,易被呼吸作用利用的糖类减少。D6时,2个品种(系)的坛紫菜呼吸熵均<1,表明呼吸底物由糖类转换为脂肪,无氧呼吸虽能提高呼吸熵,但由于强度较弱未能明显提高呼吸熵。
本研究中,相同黑暗时间处理下,洞头本地菜呼吸释碳速率、呼吸熵均高于浙东1号,可能是由于洞头本地菜为洞头本地长期养殖的品系,能更好地适应当地的环境条件,呼吸作用更强烈。也可能是洞头本地菜暗呼吸作用时存在较强的DO浓缩机制,可以高效利用水体中低含量的DO,同时洞头本地菜无氧呼吸作用稍强的缘故,须开展进一步的研究探讨。
4 结论
洞头本地菜较高的光合作用、呼吸作用强度,表现了对光环境较强的适应性。2个品种(系)坛紫菜光合产氧速率与光合固碳速率均在2 800 lx时最高,表明该光照度下产氧强度最大。光合熵在2 000 lx时最高,此时固定单位量碳的产氧效率最高。随着时间的延长,2个品种(系)坛紫菜光合产氧速率、呼吸耗氧速率、呼吸作用释放DIC速率、光合熵、呼吸熵基本呈下降趋势,这可能与本研究的试验方法有关,随着试验时间的延长,光合作用、呼吸作用的反应底物含量降低,影响了其作用强度。
猜你喜欢
戏曲研究(2022年2期)2022-10-24
传感器世界(2022年7期)2022-10-12
科学导报(2020年61期)2020-09-29
工程与建设(2019年2期)2019-09-02
东方教育(2017年9期)2017-07-19
农业工程技术·温室园艺(2016年9期)2017-04-23
应用海洋学学报(2015年4期)2015-11-24
———评郭庆财博士《南宋浙东学派文学思想研究》一书
山西师大学报(社会科学版)(2014年3期)2014-01-24
终身教育研究(2012年2期)2012-03-25
