
N末端改造提高GH11家族木聚糖酶热稳定性的研究进展
2019-03-08 08:50蒋玥凤李秀婷
食品科学 2019年3期
侯 洁,蒋玥凤,熊 科,杨 然,李秀婷,*
(1.北京工商大学 北京食品营养与人类健康高精尖创新中心,北京 100048;2.北京工商大学食品学院,北京 100048;3.北京工商大学 北京市食品添加剂工程技术研究中心,北京 100048)
木聚糖是半纤维素的主要组成成分,广泛存在于植物细胞壁中,是自然界最丰富的生物质资源之一[1]。木聚糖酶(EC 3.2.1.8)是一组能够将木聚糖降解为低聚木糖和木糖的复合水解酶,主要包括β-1,4-D-外切木聚糖酶、β-1,4-D-内切木聚糖酶和β-木糖苷酶;其中,β-1,4-D-内切木聚糖酶作用于主链内部的木糖苷键,起主要降解作用[2-3]。木聚糖酶可分为GH5、7、8、10、11和GH43等家族[4],根据其结构组成以及理化性质的不同,主要分为GH10和GH11家族[5]。
近年来,木聚糖酶广泛应用于造纸、饲料、食品等行业[6]。由于许多工业生产过程需要在高温条件下进行,热稳定性差成为大多数天然木聚糖酶在工业应用上的瓶颈。与GH10族木聚糖酶相比,GH11家族木聚糖酶具有催化效率高、底物专一性好等优点,引起了研究者的广泛关注。为了使酶学性质优良的GH11家族木聚糖酶在高温条件下的应用不受限制,越来越多的研究人员致力于GH11家族木聚糖酶热稳定性的改善。大量研究表明N末端区域序列对于GH11家族木聚糖酶的耐热性具有重要作用[7-9];因此,许多研究者通过N端改造的方法成功提升了GH11家族木聚糖酶的热稳定性。本文从GH11家族木聚糖酶分子的结构及性质特点、影响其热稳定性的关键因素、N末端改造提升木聚糖酶耐热性的技术和方法,以及耐热木聚糖酶的应用领域方面进行综述,以期为GH11家族木聚糖酶的耐热性研究提供一定的参考。
1 GH11家族木聚糖酶
GH11家族木聚糖酶由2 个反向平行的β-折叠和1 个α-螺旋构成,空间结构如图1所示,呈“右手半握状”,β-折叠的一侧扭转与另一侧形成裂缝,活性中心位于裂缝的凹槽处[10]。木聚糖酶的催化机制与其他糖苷水解酶类似,为双替换机制[3]。其中一个氨基酸作为酸碱催化剂使底物质子化,另一个氨基酸作为亲核体对底物进行亲核攻击,产生的糖片段与活性中心分离,促进酶-糖基共价中间体的形成,而后水分子上的羟基结合至正碳离子,质子结合至亲核基团,反应结束[11]。由于GH11家族木聚糖酶底物结合位点位于较深的裂缝中,与GH10家族相比,与底物结合更紧密,从而具有较强的底物专一性,水解产物中低聚木糖的含量较多,单糖较少,且具有广泛的酸碱适应性[12-13]。
GH11家族的木聚糖酶的分子质量一般小于30 kDa,多为单一结构域。虽然GH11家族的木聚糖酶具有优良的酶学特性,但是大多属于中温木聚糖酶,热稳定性较差。表1为部分GH11家族木聚糖酶的酶学性质,最适反应温度均低于50 ℃,温度高于55 ℃时不稳定。
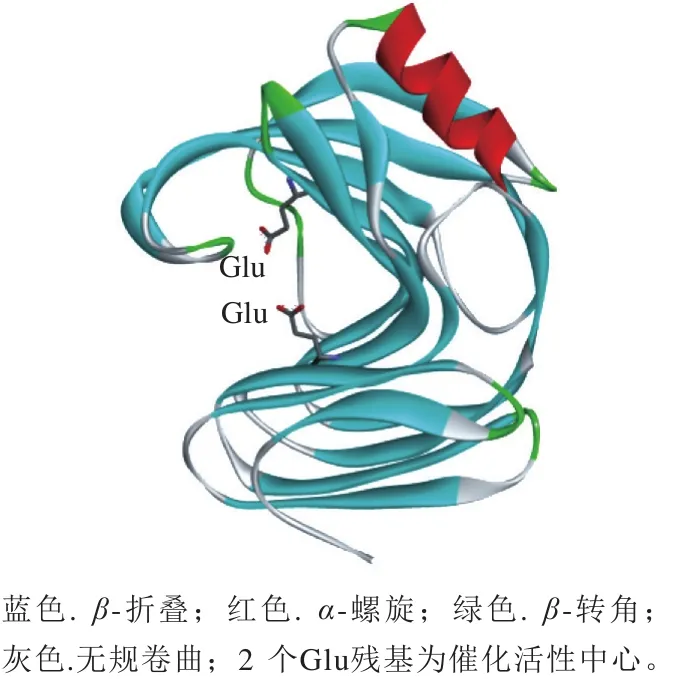
图1 GH11家族木聚糖酶空间结构Fig.1 Spatial structure of GH11 xylanases

表1 GH11族木聚糖酶的酶学性质Table1 Enzymatic properties of GH11 xylanases
2 影响GH11家族木聚糖酶热稳定性的因素
生物信息学分析手段以及基因工程技术的发展,使得木聚糖酶热稳定性相关机制的研究更加透彻。在研究的过程中发现许多因素都会对木聚糖酶的耐热性造成重要影响,其中比较重要的因素包括氢键和盐桥的数量与位置、氨基酸的组成与比例、二硫键和疏水相互作用等。
2.1 氢键和盐桥
氢原子与一些电负性强的原子形成偶极矩较大的基团,导致电子云偏向带负电的原子,当裸露的氢核遇到其他电负性强的原子时,产生静电相互作用,即氢键。氢键是稳定蛋白质二级结构的主要作用力。Purmonen[21]、Vieira[22]等的研究发现耐热木聚糖酶氢键的数量一般要高于中温木聚糖酶。另外,蛋白质分子与水之间也可以形成氢键。大多数蛋白质分子趋向于主肽链之间形成的氢键数量最多,同时应尽可能保证侧链与介质水之间的相互作用,这种相互作用有利于蛋白质结构的稳定[23]。
盐桥是指带电荷氨基酸侧链的正负基团相互接近时,形成的静电相互作用。盐桥的形成对保持蛋白质分子的构象产生积极影响,有助于改善蛋白质分子的稳定性[24]。Xue Huping等[25]对Neocallimastigales rumen来源的木聚糖酶进行突变后,突变体XynR8的热变性温度Tm值与野生型相比升高约9 ℃。根据三维建模结果推测,氨基酸突变后盐桥的形成可能是其耐热性提高的重要原因。
2.2 氨基酸的组成和比例
通过比较GH11家族中温木聚糖酶和耐热木聚糖酶的序列和结构信息,发现蛋白质中所含氨基酸残基的种类对耐热性具有一定的作用。Sriprang等[26]在对黑曲霉BCC14405木聚糖酶XylB的改造过程中,用精氨酸残基替换蛋白质分子表面的丝氨酸残基或苏氨酸残基后,所形成的突变株ST4和ST5在50 ℃的半衰期比野生型提高了18~20 倍。精氨酸为碱性氨基酸,位于分子表面时,可以通过与介质水之间发生偶极-电荷作用形成水化层,增加蛋白分子的稳定性。
脯氨酸的N原子位于吡咯环上,使前一个氨基酸与其形成肽键(CaüN)时不能自由转动,有利于增强蛋白的刚性。同时,吡咯环也具有疏水性,以上特性导致脯氨酸的构形熵小于其他氨基酸,所连接的β-折叠股比较稳定[27-28]。采用脯氨酸替换中温木聚糖酶中某些分子自由度高的氨基酸,有望提升其耐热性。但替换单一氨基酸的效果可能并不明显,Yang Wenhan等[29]在替换脯氨酸的同时引入一个二硫键,突变体在60 ℃的半衰期提升了22 倍。
芳香族氨基酸含有苯环结构,包括酪氨酸、苯丙氨酸、色氨酸。芳香族氨基酸的相互作用对维持蛋白质的稳定具有重要贡献。Georis等[30]在中温木聚糖酶Xyl1中引入芳香族氨基酸酪氨酸,使其最适反应温度提高了10 ℃,结构分析显示Y11及Y16氨基酸分别存在于β-折叠B1和B2链上,这两个芳香族氨基酸的相互作用提高了木聚糖酶N末端的稳定性。组氨酸的咪唑环与芳香族氨基酸的苯环的相互作用也有助于酶分子的稳定。
此外,氨基酸的比例也对GH11家族木聚糖酶的热稳定性具有一定影响。Hakulinen等[31]发现在耐热木聚糖酶中,苏氨酸/丝氨酸的比值较高。
2.3 二硫键
二硫键是蛋白分子空间结构上位置相邻的两个半胱氨酸残基的巯基基团发生氧化反应形成的共价键。二硫键与蛋白质的折叠过程密切相关,其键能远大于其他次级键,在维持高级结构的稳定性方面具有十分重要的作用。研究表明,二硫键通过降低蛋白质解折叠状态的熵值,从而稳定酶分子的构象[32]。在GH11家族天然的木聚糖酶中,发现少数耐热酶含有一个二硫键,存在于β-折叠股B9和α-螺旋之间[33]。虽然,在GH11家族野生型木聚糖酶的N末端一般不存在二硫键,但是在与其结构相似的GH12家族葡聚糖酶中,大多N末端区域存在二硫键。并且,经过定向进化随机突变获得的超耐热木聚糖酶Evxyn11TS的N末端也形成了一个二硫键,使木聚糖酶分子的结构更加紧凑。因此,通过在GH11家族木聚糖酶合适的位置引入二硫键,有利于改善蛋白酶的热稳定性。
2.4 疏水作用
疏水作用是指蛋白在水介质中形成折叠时,大部分疏水残基倾向于被埋藏在分子的内部。由于木聚糖酶分子内部为典型的疏水结构,疏水相互作用的增强可以促进疏水核心的有效包埋。通过分析比较耐热酶与中温酶的三维结构,发现耐热酶趋向于通过包装效应形成更密实的空腔,从而使分子结构更加紧凑和稳定。Kim等[34]发现了更有利于蛋白质稳定性的疏水结构簇构象,通过定点突变(W58F、G64A、K95L、H156V、M158L和V168I)将其整合到环状芽孢杆菌木聚糖酶分子内,使木聚糖酶的局部结构更加稳定,突变体半衰期提高了78 倍,Tm值提高了8.8 ℃。You Chun等[35]构建突变体G201C和C60A-G201C,Tm与野生型相比分别提高约10 ℃和12 ℃;同源建模结果显示,突变酶中氨基酸残基C50、C60与C201在空间相近,但是硫醇滴定结果排除了二硫键的存在;进一步分析表明氨基酸残基C50和C201在空间结构上形成疏水基团相互作用。由此可知,半胱氨酸除了形成二硫键使蛋白酶分子结构更加稳定,也可以通过疏水相互作用来提升蛋白酶分子的热稳定性。
3 N末端区域的改造策略
N末端区域是GH11家族木聚糖酶分子解折叠的起始区域,与GH11家族木聚糖酶的耐热性密切相关[36]。Lee等[37]对耐热木聚糖酶XynA的N末端区域进行删除突变,突变体的热稳定性丧失但酶活性保持不变,从而确定了N末端是其热稳定区。也有研究表明,GH11家族的耐热木聚糖酶与中温木聚糖酶相比大多存在较长的N端,这部分序列趋向于形成1 个β-折叠片段,而中温木聚糖酶N末端区域的相应位置主要为一段无规卷曲序列[38]。在二级结构中,无规卷曲的柔性较大,与α-螺旋和β-折叠相比稳定性较差。因此,通过对中温木聚糖酶的N端序列进行分析和改造,有助于提升GH11家族木聚糖酶热稳定性,更好地发挥其应用价值,N末端区域改造的主要策略示意图见图2。
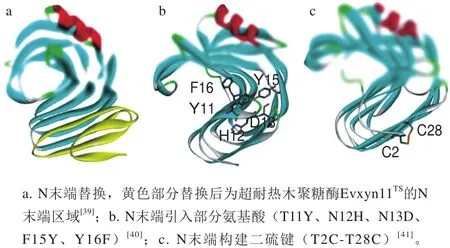
图2 N末端区域改造策略示意图Fig.2 Schematic illustration of N-terminal modif i cation
3.1 N末端替换
研究表明,利用耐热木聚糖酶的N末端替换中温木聚糖酶的N末端,可以一定程度提高中温木聚糖酶的热稳定性。
首先需要选择合适的耐热木聚糖酶N末端片段。通过对耐热木聚糖酶与中温木聚糖酶的序列同源性进行比对和分析,判断所研究的中温木聚糖酶热稳定性差的根本原因是否与其N末端序列相关,选择全序列同源性较高且N末端同源性较低的耐热木聚糖酶作为替换模板,使得整体序列的差异性较小,有利于融合突变体的成功表达。Evxyn11TS和TfxA属于目前发现的较少数GH11家族的耐热木聚糖酶,由于其N末端均被证实对热稳定性有突出贡献,经常被研究者作为替换模板。其中,木聚糖酶Evxyn11TS是通过定向进化获得的重组酶,且在7 个突变位点中,6 个存在于N端,其Tm值为101 ℃,在85 ℃保温15 min后,剩余酶活力仍为85%[42]。TfxA属于天然未经改造的重组酶,在75 ℃下保温18 h仍有96%的剩余酶活力[37]。
其次还需要确定替换片段的长度,根据序列比对分析及分子动力学模拟等方法,确定合适的替换位置,能够在提升热稳定性的同时,保持中温木聚糖酶的本身优良性质。Yin Xin等[43]将Aspergillus oryzae来源的中温木聚糖酶AoXyn11N末端的57 个氨基酸替换成耐热酶Evxyn11TS的相应序列,重组酶NhXyn1157最适反应温度提高了25 ℃,热稳定性由40 ℃提升至60 ℃。Zhang Huimin等[39]也采用Evxyn11TSN末端的38 个氨基酸替换了中温木聚糖酶Auxyn11AN末端的33 个氨基酸,重组酶AEXynM的Tm提升了34 ℃,进一步的研究表明,氨基酸Cys5、Pro9、和His14对重组酶热稳定性的提升起主要作用。Sun Jianyi[44]、杨浩萌[45]等均选择耐热木聚糖酶TfxA的N末端,分别替换中温酶AnxA及XYNB的N末端,最适反应温度均提高了10 ℃。
分析耐热N末端突变体的结构特征及氨基酸相互作用,与突变前木聚糖酶分子进行同源建模比较,找到影响耐热性提升的关键氨基酸位点,可以为其他改造策略中突变位点的选择提供更广阔的思路。
3.2 N末端引入部分氨基酸
木聚糖酶的来源广泛,不同来源的GH11家族木聚糖酶,序列具有一定的差异性,因此通过突变大量氨基酸提升热稳定性的方法存在一定限制。N末端整体替换的策略涉及较多的氨基酸改变,可能会对突变体的酶学性质造成消极影响。Song Letian等[46]所构建的N末端延伸了17个氨基酸的突变体Ntfus,热稳定性和热变性温度均稍有下降,而催化效率有所提高。Zhang Shan等[40]通过对耐热突变体STxAB的N末端区域进行分段研究,确定了5 个突变位点(T11Y、N12H、N13D、F15Y、Y16F)对该酶耐热性的提升起到主要作用;在此基础上,又结合定点突变N32G和S33P,进一步提升了其热稳定性[47]。由此可知,N末端区域引入部分氨基酸,可以对GH11家族木聚糖酶的热稳定性起到良好的改善作用。
为了减小对酶分子空间结构的影响,考虑采用N末端引入部分氨基酸而非N端全部替换的方式进行耐热性改造。目前木聚糖酶的空间结构和催化机理已知,可以使用生物信息学手段进行辅助设计,选择合适的突变位点。杨浩萌等[48]在橄榄绿链霉菌Streptomyces olivaceoviridis木聚糖酶XYNB的N末端区域分别引入点突变N13D和S40E,在70 ℃处理5 min后,与野生型相比,突变体的热稳定性分别提高了24.76%和14.46%。柏文琴等[49]通过理性设计在GH11家族碱性木聚糖酶Xyn11A-LC的N末端区域同时引入芳香族氨基酸T9Y和D14F,突变体的Tm值由55.3 ℃提升至67.9 ℃。Watanabe等[50]结合多序列比对结果,在GH11家族中温木聚糖酶XylC的N末端区域选择9 个氨基酸位点进行突变,变性温度显著提升20 ℃,且仍保持较高的酶活力。通过对突变体进行晶体结构分析,发现N末端区域结构发生变化,推测N末端区域的变化是突变体耐热性改善的原因。
热稳定性的大幅度提升往往需要多个位点的同时突变,而突变氨基酸数目和位置影响酶分子的空间结构。近年来,有研究表明N末端区域与底物结合方式及催化效率相关[51-52]。因此,应用定点突变技术在N末端区域改造木聚糖酶热稳定性时,需要深入分析并充分了解目标木聚糖酶的序列特点,避免酶活力的损失。
3.3 N末端构建二硫键
大量研究表明,N末端二硫键的存在是GH11家族木聚糖酶Evxyn11TS耐热的重要原因,将相应位置的半胱氨酸突变为苏氨酸,突变体的最适反应温度和热稳定性均有明显下降[53]。以Evxyn11TS为模板进行N末端替换,对耐热突变体AoXyn11A[54]和SyXyn11[55]采用定点突变的方法去除二硫键后,耐热性均降低。许多研究者也已经通过在GH11家族中温木聚糖酶的N末端区域引入二硫键改善其热稳定性。Fenel等[41]通过构建二硫键T2C-T28获得的突变体,在65 ℃下的半衰期由40 s延长至20 min。N末端二硫键Q1C-Q24C的引入,使GH11家族木聚糖酶TLX的最适温度和热变性温度均提升了10 ℃[56]。因此,在N末端区域引入二硫键,对热稳定性的提升具有重要作用。
形成二硫键的两个半胱氨酸残基,一般空间距离很近,而序列距离可能很远。通常条件下,二硫键的形成受周围氨基酸环境的影响和限制。若要成功地在GH11家族木聚糖酶分子中构建二硫键,首先需要根据三维建模结果,结合二硫键预测软件(Disulfide by Design),找到可以突变形成二硫键的氨基酸对,与已知含二硫键的耐热木聚糖酶序列进行比对分析,从而确定引入二硫键的最优位置构建突变体,然后再验证二硫键是否形成。通过比较断开二硫键前后突变体在聚丙烯酰胺凝胶电泳的迁移率,可以初步判断二硫键是否构建成功[57]。采用Ellman试剂测定加入还原剂前后的巯基含量,同样可以检测二硫键的存在并估算其数量。一般而言,若要确认二硫键形成的具体位置,还需要经过酶解-质谱法进行验证和分析。
鉴于在GH11家族少量的耐热木聚糖酶中,发现分子内部α-螺旋与β-折叠之间存在二硫键,有研究者在与其序列相似度高的木聚糖酶分子的相应位置引入二硫键,成功构建了热稳定性提升的突变体[58-59],证实了二硫键对酶分子结构稳定性的重要作用。与分子内引入二硫键相比,在N末端构建二硫键更有利于蛋白质新生肽链的延伸,可减少错误折叠的可能性。
4 耐热木聚糖酶的应用
木聚糖酶作为一种绿色高效的生物催化剂,广泛应用于食品、饲料、造纸、医药、能源等领域。不同领域对于木聚糖酶的需求不同,对应酶的酶学性质具有差异。GH11家族木聚糖酶由于催化效率高、底物专一性强,具有良好的应用潜力。然而,在实际生产过程中,经常会遇到高温环境。例如在食品工业中,较高的温度可以提升底物的溶解性并防止杂菌污染[60];造纸工业中需在高温下对纸浆进行漂白处理[61];在饲料工业中同样存在短暂高温制粒过程[62]。因此,耐热木聚糖酶具有巨大的应用价值和广阔的市场前景。
4.1 食品工业
阿拉伯木聚糖是小麦面粉中非淀粉多糖的主要成分,其中水不溶性阿拉伯木聚糖含量与面团流变性和面制品口感密切相关。在面粉中添加适量的木聚糖酶可以使不溶性阿拉伯木聚糖增溶,提高面团的柔软度和拉力,从而改善面制品品质。在面包等焙烤食品中加入木聚糖酶,可以显著增大面包体积,改善面包心质地,降低面包的老化速率,延长货架期[63-64]。在果蔬汁的生产过程中,木聚糖酶能够使果汁澄清,同时释放寡糖和单糖,并提升口感[65]。酿酒工业中,木聚糖酶可以通过破坏原料细胞结构,降低啤酒的黏度,增强麦芽汁的滤过性能,达到提高产率的目的[66]。
木聚糖的水解产物中含有低聚木糖,能够作为益生元提高双歧杆菌活性、改善肠道环境、帮助消化、增强人体免疫力;同时也可以代替蔗糖作为低能量甜味剂,避免人体血糖水平大幅度波动,适于生产糖尿病人所需的低糖食品。
4.2 饲料工业
在饲料工业领域中,木聚糖酶能够破碎植物细胞壁,作用于饲料中的非淀粉多糖,消除其抗营养作用,从而提高蛋白质、脂肪、淀粉等养分的消化吸收。研究表明,在小麦日粮中添加木聚糖酶能够提高家禽的养分消化率和日粮代谢能值[67]。木聚糖酶通过降解饲料中的黏性木聚糖成分,可以降低食糜黏性,提高消化酶对底物的作用效率,因此广泛应用于饲料添加剂中。Hernández等[68]将木聚糖酶与酿酒酵母混合作为饲料添加剂,有效降低了反刍动物肠道温室气体的排放量,对环境保护具有重要意义。根据动物消化道pH值较低的特点,饲料添加剂中选取酸性木聚糖酶更加合适[69]。
5 结 语
随着GH11家族木聚糖酶耐热机制研究的逐步深入,N末端区域被认为是影响GH11家族木聚糖酶热稳定性的关键区域。许多研究人员通过N末端整体替换、定点突变,以及在此区域引入二硫键等方式大幅度提升了GH11家族木聚糖酶的热稳定性。不同来源的木聚糖酶蛋白序列具有一定差异;因此在选择N末端改造策略时,首先要对待改造的木聚糖酶基因进行系统的生物信息学分析,通过结合计算机辅助设计等方法,找到对热稳定性有重要作用的关键区域或氨基酸位点,而后采用合适的策略进行耐热性改造。此外,GH11家族木聚糖酶分子的α-螺旋区域同样对其结构稳定性至关重要。近年来,也有研究者将N末端改造与分子内部改造相结合进行复合突变,进一步提升了GH11家族木聚糖酶的热稳定性[55]。
木聚糖酶热稳定性的研究有助于降低工业生产成本和市场应用的推广。虽然目前已经在分子水平上对木聚糖酶的结构信息和耐热机制进行了一定的了解,也通过蛋白质工程成功构建了热稳定性提升的突变体,但是与超耐热的蛋白酶分子相比仍有差距。因此,GH11家族木聚糖酶耐热性的提升还存在一定发展空间,有望通过更加先进的结构技术进一步深入和系统地研究,从而取得突破性进展。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
分析化学(2019年4期)2019-05-13
陶瓷学报(2019年5期)2019-01-12
中国饲料(2016年17期)2016-12-01
中国塑料(2016年7期)2016-04-16
中国塑料(2015年6期)2015-11-13
中国塑料(2015年6期)2015-11-13
中国塑料(2015年8期)2015-10-14
安徽农学通报(2015年4期)2015-07-30
质量与标准化(2015年11期)2015-07-21
