
酵母菌对重金属的吸附与抗性和解毒重金属的胞内分子机制研究进展
2019-03-08 08:50李丽杰贺银凤
食品科学 2019年3期
李丽杰,贺 敏,贺银凤,*,孙 禹
(1.内蒙古农业大学食品科学与工程学院,内蒙古 呼和浩特 010018;2.内蒙古包头市疾病预防控制中心,内蒙古 包头 014030)
重金属是指密度大于5的金属,不同领域对重金属的规定不同,一般来说,多半是以环境污染领域的定义为准,包括汞、镉、铅、铬、锌、铜、钴、砷等。重金属不仅可以通过水体、空气、土壤等污染环境使生态问题恶化,而且会导致食品中的有毒重金属残留蓄积,引起严重的食品安全问题,在造成巨大经济损失的同时严重威胁人类健康。
对于去除环境及食品中的重金属污染问题,微生物吸附因其具有无污染、成本低等优点已成为广泛认可的重金属脱除方法。目前,越来越多的研究发现酵母菌对重金属具有良好的吸附能力和抗性,其中有些酵母菌可以在食品发酵工业中应用,而这类微生物对重金属的脱除及解毒机制更是最新的研究热点。
目前,关于酵母菌对重金属解毒机制的研究主要集中于如何通过胞外的吸附、沉淀等作用缓解重金属的毒性,但对重金属胁迫下酵母菌细胞内对重金属的抗性及其解毒机制进展的报道较少。本文介绍酵母菌对重金属的抗性及吸附情况,阐述不同种、属对不同类别重金属的去除情况,重点从胞内分子角度分析了酵母菌对重金属的抗性机制,归纳了解毒过程中关键性的基因和蛋白质以及它们的功能作用,旨在为有活性的酵母菌应用于生态环境和被重金属污染的发酵食品提供理论依据。
1 酵母菌对重金属的吸附与抗性情况
酵母属、假丝酵母属、毕赤酵母属中的多种酵母菌对重金属具有很高的抗性,并且有一些是良好的重金属吸附剂。对于酵母属中酿酒酵母(Saccharomyces cerevisiae)吸附重金属的研究较早也比较广泛,主要集中于测定酿酒酵母对不同重金属离子的吸附情况。相关研究表明,酿酒酵母对Cd2+、Pb2+、Cu2+、Co2+、Al3+、U4+、Zn2+、Cr3+、Ni2+等重金属离子具备较强的吸附能力,吸附的基质主要是废水和液体培养基[1-3]。
近几年,对其他菌属酵母菌吸附及抗重金属的研究也逐渐深入,主要是从不同的生态环境中筛选吸附和抗重金属的酵母菌,并且对其吸附及抗性进行研究,包括菌体浓度、重金属浓度及其他离子存在对其吸附及抗性的影响等。重金属镉在自然界中广泛残留,所以酵母菌对镉的吸附和抗性研究最为深入。众多学者发现红酵母(Rhodotorula sp.)Yll、马克思克鲁维酵母(Kluyveromyces marxianus)YS-K1、鲁氏接合酵母(Zygosaccharomyces rouxii)、库德毕赤酵母(Pichia kudriavzevii)、南极红酵母(Rhodotorula mucilaginosa)AN5、土生隐球酵母(Cryptococcus humicola)、热带假丝酵母(Candida tropicalis)PS33等在不同菌体添加量下,分别在3~1 232 mg/L Cd2+溶液中具有很高的镉抗性与积累特性[4-9],同时某些菌种具有很强的镉脱除能力,在一定条件下去除率可以达到90%以上,明显高于酿酒酵母[4-5,8-9]。魏轲研究发现,以上部分菌株除对重金属镉有一定的吸附和抗性外,南极红酵母(Rhodotorula mucilaginosa)AN5在含有极高浓度的CuSO4、CuCl2、Pb(CH3COO)2、CrCl3、MnCl2和HgCl2等重金属的培养基中也可以存活并缓慢生长[6]。而Fernández等也证明分别有55%和80%从南极分离的南极酵母能够抵抗1 mmol/LCr3+和Cu2+[7]。据Andreeva等的报道,土生隐球酵母(Cryptococcus humicola)对浓度为2.502 mmol/L的Mn2+、Co2+、Ni2+、Zn2+、La3+也具有抗性[10];在含有葡萄糖、磷酸盐和2 mmol/L金属盐的培养基中,除镉外其还可以积累钴和铁,去除率分别约为17%和4%[11]。
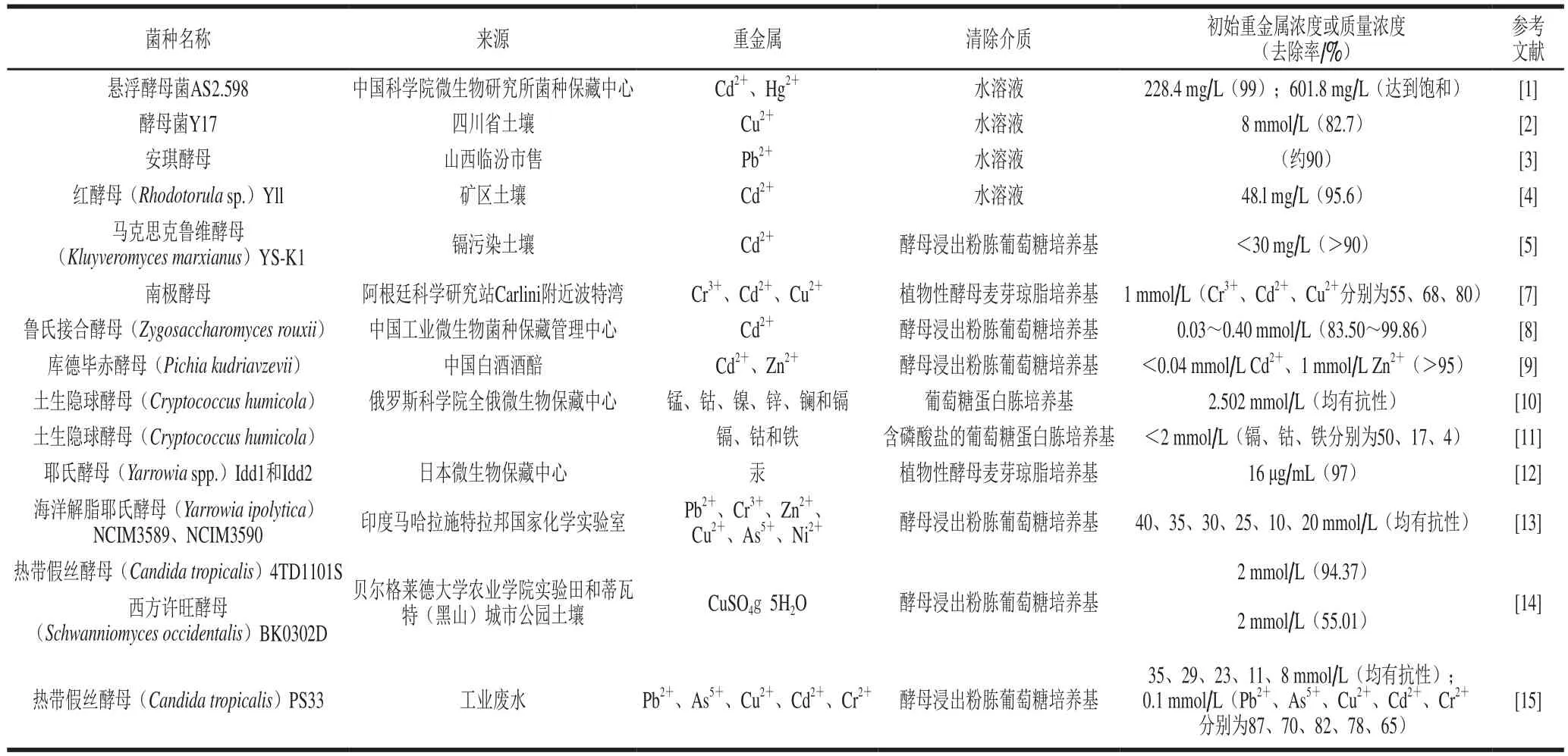
表1 酵母菌对重金属的吸附及抗性情况Table1 Heavy metal biosorption by yeasts and their resistance to heavy metals
另外,Oladunjoye等发现耶氏酵母属(Yarrowia spp.)中的菌株Idd1和Idd2通过生物累积、挥发和微量沉淀可去除培养基中97%以上的汞[12]。Bankar等对两种海洋解脂耶氏酵母菌株的重金属抗性进行测定后推测,两种解脂耶氏酵母对Pb2+、Cr3+、Zn2+、Cu2+、As5+、Ni2+等均具有抗性[13]。Radić等从来自郊区和城市污染的土壤中分离出热带假丝酵母(Candida tropicalis)和西方许旺酵母(Schwanniomyces occidentalis),这两种酵母在2 mmol/L CuSO4g5H2O存在下能大量生长,显示出对铜的抗性[14]。而Ilyas等从工业废水中分离出的热带假丝酵母PS33同样显示出对35 mmol/L Pb2+、29 mmol/L As5+、23 mmol/L Cu2+、8 mmol/L Cr2+具有一定的抗性。培养8 d后,热带假丝酵母PS33能从培养基中除去70% As2+、82% Cu2+、65% Cr3+和87% Pb2+[15](表1)。
本实验室从内蒙古重金属污染区分离筛选出对重金属铅具有不同抗性和吸附能力的酵母菌,并且通过研究发现,菌株对重金属的抗性与吸附性间不存在对应关系。其中低抗铅的2 株金佩梅奇酵母(Metschnikowia chrysoperlae)对Pb2+的去除率高达92.56%以上,而高抗铅的3 株胶红酵母(Rhodotorula mucilaginosa)对Pb2+的去除率最高却仅为23.14%。人们更希望获得高抗及高吸附性的菌株用于对重金属的去除,具有高抗性的酵母菌才能在重金属污染的环境中生长繁殖并保持活性,从而更好地发挥微生物对重金属的吸附及解毒作用。因此本实验室筛选获得的对Pb2+的抗性质量浓度在6 000~7 000 mg/L之间、去除率可达78.94%~91.67%的4 株高抗、高吸附性的异常威客汉姆酵母(Wickerhamomyces anomalus)具备一定的应用潜力,也是进一步研究的重点。
2 酵母菌解毒重金属的胞内分子机制
目前,在对重金属胞内解毒分子机制研究方面,酿酒酵母作为第一个被全基因组测序的真核模式生物,是唯一已经阐明机制的酵母菌。而对于其他酵母菌解毒重金属胞内机制的研究不尽相同。总结起来,酵母菌解毒重金属的机制包括酵母菌含硫代谢通路的重构/谷胱甘肽合成机制、与缓解氧化应激相关的基因和蛋白表达、将重金属从胞内运出或使其封闭在液泡内的转运蛋白的运输作用、与重金属离子结合降低其毒性的金属硫蛋白和植物螯合肽的结合作用等。
2.1 谷胱甘肽合成的解毒机制
硫吸收和谷胱甘肽生物合成对于所有生物体都是必需的。谷胱甘肽是防御氧化应激和重金属解毒的关键因素,其以以下4 种方式进行重金属解毒:1)结合重金属生成复杂的蛋白质,从而介导液泡隔离;2)具有抗氧化作用,可以缓解细胞氧化应激;3)可能与蛋白质上的活性巯基结合(即蛋白质谷胱甘肽化),从而使它们免于与不可逆的重金属结合和/或氧化损伤;4)可以在细胞外与重金属螯合形成复合物,并且这种复合物不容易再进入细胞。
对于酵母菌的抗镉机制最初是由Lee等研究的,他们发现反式激活因子Yap1p可以调节酵母菌对氧化应激的适应性反应,并且在抗镉方面很重要,原因是Yap1p是谷胱甘肽合成过程中转录水平上的调节因子[16]。之后,在21世纪,Vido等利用双向凝胶电泳技术分析了酵母细胞在急性镉胁迫下的蛋白质差异表达情况,发现含硫氨基酸生物合成途径的9 种酶被强烈诱导,这种诱导同样依赖于反式激活因子Yap1p[17]。而Fauchon等的研究与Vido等一致,同样发现酿酒酵母的镉应答机制之一是转录调控因子Met4p与Yap1p,其通过调控酵母菌摄取硫以增强谷胱甘肽的合成,从而协同调节谷胱甘肽产生[18]。
Lafaye等进一步通过蛋白质组学和代谢组学定量了镉处理的酿酒酵母细胞中8 种涉及谷胱甘肽生物合成途径的含硫代谢物(即半胱氨酸、高半胱氨酸、甲硫氨酸、γ-谷氨酰半胱氨酸、胱硫醚、谷胱甘肽的还原和氧化形式、S-腺苷高半胱氨酸),发现与对照组相比,大部分代谢物的细胞内浓度增加,证明了酵母菌中谷胱甘肽的大量产生对于重金属镉的解毒至关重要。在重金属镉存在的条件下,含硫蛋白质的合成显著下降、含硫代谢物重构流向谷胱甘肽通路[19]。Huang Xinhe等进一步证明镉暴露增加了细胞谷胱甘肽水平、降低了氧化型谷胱甘肽水平,并且还降低了细胞的氧化型谷胱甘肽/谷胱甘肽比例[20]。
在酿酒酵母的抗砷方面,Thorsen等通过定量转录组学、蛋白组学和硫代谢物分析探讨了酿酒酵母的抗砷(亚砷酸盐)机制,最重要的结论是:几乎所有硫酸盐同化和谷胱甘肽生物合成途径的组分均在基因和蛋白质水平被诱导。并且确定了核心转录调控因子为Yap1p和Met4p,与其抗镉机制一致[21]。
随着包括基因组学、转录组学、蛋白组学和代谢组学等各层面的组学研究手段的应用,其他能吸附和抗重金属酵母菌的抗性机理逐渐被研究。
在粟酒裂殖酵母(Schizosaccharomyces pombe)中的镉解毒机制同样涉及谷胱甘肽,在镉胁迫下,粟酒裂殖酵母中的硫化物合成也增加,并且硫化物参与具有高镉结合能力的高分子质量植物螯合肽-镉-含硫复合物的产生。Jin等对粟酒裂殖酵母的镉和砷抗性基因进行筛选后发现,在36 个与镉和砷的抗性均相关的基因中涉及3 个生物学过程,最主要的就是含硫化合物代谢过程,特别是硫氨基酸(半胱氨酸和甲硫氨酸)和谷胱甘肽的生物合成[22]。
除了酿酒酵母和粟酒裂殖酵母的重金属抗性与谷胱甘肽合成有关外,Ilyas等从工业废水中分离出抗重金属胶红酵母(Rhodotorula mucilaginosa)。与对照组相比,在100 mg/L CdCl2、NaAsO2、CuSO4、Pb(NO3)2的胁迫下,其谷胱甘肽含量均有所增加[23]。而最近Khan等进一步验证了镉诱导下热带假丝酵母3Aer中谷胱甘肽生物合成增加[24]。Ilyas通过实验证实另一株假丝酵母PS33经0.1 mmol/L砷和铅处理后,细胞中谷胱甘肽水平显著升高[23]。
2.2 与酵母菌解毒重金属相关的基因和蛋白

表2 酵母菌中与重金属解毒相关的基因Table2 Genes related to heavy metal detoxif i cation in yeasts
Jin等[22]通过转录组、蛋白组与代谢组学技术以及生物信息学分析发现,在铜、镉、汞、锌、铬、砷等重金属胁迫下,酵母菌会产生一些对重金属响应一致的基因,称为常见的金属响应(common metal-responsive,CMR)基因。被诱导的CMR基因在与重金属离子转运和稳定、活性氧(reactive oxygen species,ROS)的解毒、碳水化合物代谢、脂肪酸代谢、多胺转运和RNA聚合酶II转录等相关的生物过程中被富集,而被抑制的CMR基因在与多糖生物合成、G蛋白信号传导、蛋白质靶向和转运有关的生物过程中富集[22]。另外,在不同重金属胁迫下,酵母菌不同菌属基因表达的变化不同。表2中例举的基因主要包括缓解重金属胁迫引起细胞应激反应基因、为了修复重金属胁迫引起细胞损伤的DNA损伤和修复基因、降低胞内ROS含量的抗氧化活性基因、维持细胞壁完整性及编码各种转运蛋白的基因等。
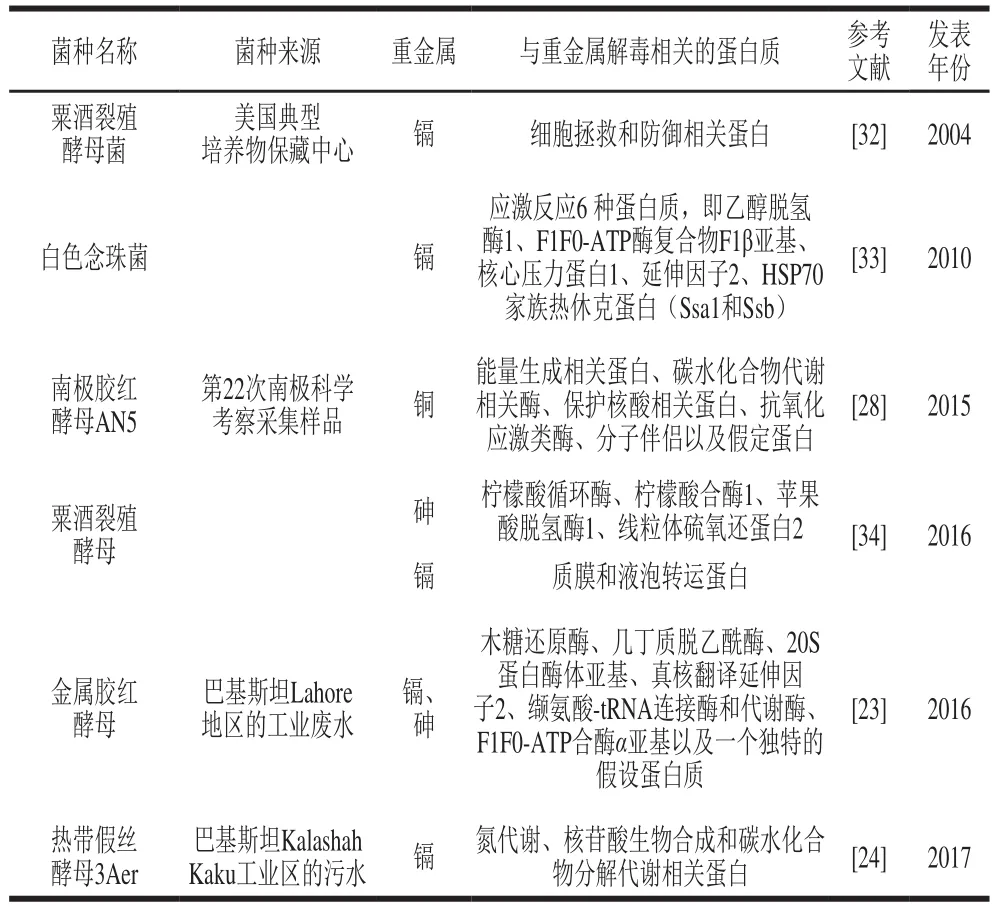
表3 酵母菌中与重金属解毒相关的蛋白Table3 Proteins related to heavy metal detoxif i cation in yeasts
表3中列举的蛋白功能同样主要集中在细胞防御和拯救、蛋白修饰、转录、信号传导、甘油运输、电子传递、应激反应和核糖体生物合成、抗氧化活性、细胞壁合成、细胞内转运等方面。
随着蛋白组学和转录组学等技术的推进,除了对酿酒酵母重金属解毒机制研究得比较深入外,对其他酵母菌的抗重金属机制的研究也逐渐展开。其中比较深入的是对粟酒裂殖酵母与重金属抗性相关基因和蛋白的研究。Bae等系统研究了粟酒裂殖酵母在镉胁迫下的细胞反应,并与酿酒酵母进行对比后发现,镉暴露后粟酒裂殖酵母80%的上调蛋白质不同于酿酒酵母,在粟酒裂殖酵母中未观察到诱导半胱氨酸合酶表达。作为镉解毒的替代机制,粟酒裂殖酵母显著产生更高水平的无机硫化物,以固定细胞镉作为CdS纳米晶体覆盖谷胱甘肽和/或植物螯合物[32]。Guo Lan等[34]对粟酒裂殖酵母的镉和砷抗性基因进一步进行筛选后发现,辅因子生物特别是泛醌和西罗血红素、5-磷酸吡哆醛的生物合成与镉和砷的抗性均相关。
2.3 转运蛋白介导的细胞内重金属的排出和液泡隔离机制
在酵母菌细胞膜、线粒体、囊泡、液泡、内质网、高尔基体和细胞核上都有重金属转运蛋白的存在,可以运输多种重金属。这些蛋白质中有的是特异性的、有的是非特异性的,有的介导重金属的转入、有的介导重金属的转出,而基本探明的与酵母菌胞内解毒重金属相关的转运蛋白质主要有3 类。
2.3.1 位于细胞膜上转运蛋白的解毒机制
位于细胞膜上的转运蛋白通过运出途径将重金属从胞质中运出,降低重金属在细胞中的毒性。Wysocki等证明酿酒酵母中存在促进As3+外排的砷抗性-3(Acr3)p(也称为Arr3p)和促进Cd2+外排的Pca1p等蛋白,As3+通过Acr3p输出可能是酿酒酵母中最重要的砷解毒机制[35]。Acr3p是Acr3家族的转运蛋白的原型成员,其属于胆汁/亚砷酸盐/核黄素转运蛋白超家族。Pca1p是包括各种重金属如银、镉、钴、铜、铅和锌的转运蛋白,是酿酒酵母中镉输出的主要途径,对镉抗性起着关键作用[36]。水甘油通道蛋白(Fps1p)是另一种转运蛋白,是双向通道蛋白,是介导As3+和Sb3+向酵母细胞流入和流出的主要途径。当细胞内As(OH)3浓度高于细胞外时,Fps1p可能与Acr3p一起促进As3+从细胞内排出。为了防止通过Acr3p排出的As3+通过Fps1p重新进入,细胞通过排出谷胱甘肽以降低As(OH)3的细胞外浓度,细胞外As3+和谷胱甘肽形成复合物,且不能进入细胞[26]。Cui Zhifeng[37]、Nagy[38]等证明Yor1p转运蛋白也涉及重金属解毒,Yor1p属于ATP结合盒转运蛋白(ATP-binding cassette transporter protein,ABC)家族,可能介导双谷胱甘肽镉(bis-glutathionatocadmium,Cd(GS)2)形式的镉外排。
2.3.2 位于液泡膜上转运蛋白的解毒机制
位于液泡膜上的转运蛋白可以将植物螯合肽-重金属络合物或谷胱甘肽-重金属络合物转运至液泡,通过封闭隔离使重金属从胞质中去除。许多研究表明,在酿酒酵母中,Ycf1p代表了一个主要液泡隔离途径。Li Zesheng等通过含有YCF1基因缺失的酿酒酵母菌株DTY167及同基因野生型菌株DTY165之间的直接比较,证明YCF1是增加对外源谷胱甘肽S-共轭前体的毒性作用抗性所必需的[39]。Ycf1p在体外促进As(GS)3、Cd(GS)2和Hg(GS)2以谷胱甘肽络合物的形式向液泡转运。Paumi等证明两个Ycf1p同源物Bpt1p和Vmr1p在镉的解毒中也发挥了部分的作用[40]。Khan等进一步验证了YCF1基因在热带假丝酵母3Aer抗镉机制中的作用[24]。另外,袁红莉推断在酿酒酵母中像Zrc1p这样的阳离子扩散促进剂运输蛋白的运输作用和金属硫蛋白的结合作用可能都被包括在内,但主要的解毒机制是经过ABC将谷胱甘肽/金属硫蛋白复合物转运进入液泡[6]。MacDiarmid等提出阳离子扩散促进剂家族中的Zrc1p、Cot1p两种转运蛋白可以将Cd2+、Co2+、Zn2+等二价阳离子螯合到液泡中[41]。
粟酒裂殖酵母中镉的抗性涉及HMT1基因编码抗Cd2+所必需的ABC,它定位于液泡膜上[42]。含有HMT1的粟酒裂殖酵母液泡膜能够摄取植物螯合肽-Cd2+复合物。Guo Lan等在对粟酒裂殖酵母的镉和砷抗性进行研究时也证实了液泡膜跨膜运输蛋白Hmt1p是一种抗镉所需的蛋白质[34]。
2.3.3 位于线粒体、内质网、高尔基体等细胞器上转运蛋白的解毒机制
位于线粒体、内质网、高尔基体等细胞器上的蛋白质在重金属解毒中可能发挥作用,但是具体作用机制不明确。MacDiarmid等提出阳离子扩散促进剂家族中的Zrc1p、Cot1p两种转运蛋白可以将线粒体内的锌和镉排出,降低重金属对线粒体的毒害作用[41]。另外两个转运蛋白Atm1p和Atm3p可以从线粒体转运重金属离子铁硫蛋白结合物至细胞质,进行某些含金属离子蛋白辅助因子的组装,并以某种未知的方式提高宿主的重金属抗性,其转运的重金属离子主要为钼[43],但是其具体作用机制目前仍不清楚。此外,Lauer[44]、Guo Lan[34]等先后证明存在于内质网和高尔基体的P型ATP酶家族成员Pmr1p可能是通过分泌途径直接参与镉、锰解毒的蛋白质;渗透进细胞的过量阳离子可能是通过Pmr1p的作用迅速泵送,通过分泌囊泡从细胞排出(图1)。
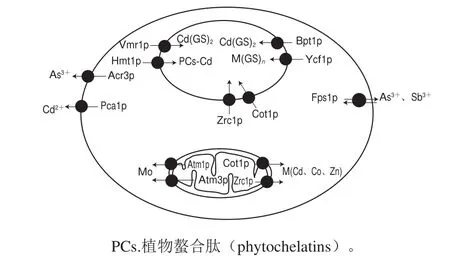
图1 酵母菌转运蛋白对重金属的解毒机制[35]Fig.1 Detoxif i cation mechanism of heavy metals in yeast through transport proteins[35]
2.4 金属硫蛋白和植物螯合肽对重金属的螯合作用
细胞中存在重金属时,会诱导低分子质量的富含半胱氨酸的蛋白质和肽如金属硫蛋白和植物螯合肽表达并与其结合,由毒性较强的离子态转变为毒性较弱的结合态,从而使重金属的毒性降低[45]。
2.4.1 金属硫蛋白对重金属的螯合作用
金属硫蛋白是一类含有较多半胱氨酸残基、分子质量较小的金属结合蛋白,广泛存在于多种生物中,是与重金属抗性和富集能力相关的蛋白中的一类。
在酿酒酵母中有两种类型金属硫蛋白,即Cup1p和Crs5p。很多研究证明,在酿酒酵母中Cup1p的主要生理作用是镉和铜的解毒,而Crs5p可以结合铜、镉特别是锌[46]。酵母金属硫蛋白主要参与铜、锌和镉的解毒机制。另外,粟酒裂殖酵母重金属硫蛋白Zym1p可以螯合锌,而缺乏Zym1p的细胞对锌和镉敏感[47]。
Radić等同时证明热带假丝酵母4TD1101S在铜胁迫下胞内金属硫蛋白产量有一定的增加,推测金属硫蛋白在热带假丝酵母对铜的抗性中有一定的作用[14]。而Yang等发现编码植物螯合素EC的基因与α-凝集素基因融合的工程化酿酒酵母表现出高效的镉吸附能力,同时镉抗性显著提高[48]。
近年来,许多研究表明利用基因工程中异源表达技术提高酵母菌重金属抗性是可行的。在酿酒酵母中表达东南景天(Sedum alfredii)的金属硫蛋白基因SaMT2后,其镉抗性和积累能力显著增强[49]。Ruta等将7 种拟南芥金属硫蛋白(AtMT1a、AtMT1c、AtMT2a、AtMT2b、AtMT3、AtMT4a和AtMT4b)和4 种天蓝遏蓝菜金属硫蛋白(NcMT1、NcMT2a、NcMT2b和NcMT3)在酿酒酵母细胞中表达后,导致酿酒酵母对Cu2+、Zn2+或Cd2+等重金属抗性增加[50]。
徐炳政[51]、王颖[52]等通过动物实验先后探讨了两种酵母源金属硫蛋白亚型(MT-1和MT-2)对急性铅中毒小鼠、慢性铅中毒小鼠、慢性汞中毒小鼠的排铅、排汞、肝脏保护作用和过氧化损伤修复作用。研究发现,两种酵母源金属硫蛋白对3 种小鼠均具有显著的排铅、排汞效果以及良好的氧化损伤保护作用,且作用效果类似于动物源金属硫蛋白。
2.4.2 植物螯合肽对重金属的螯合作用
植物螯合肽是在所有真核生物和一些原核生物中发现的肽,其由谷氨酸(Glu)、半胱氨酸(Cys)、甘氨酸(Gly)3 种氨基酸组成,一般化学式为(γ-Glu-Cys)n-Gly(n=2~11)。这些肽由谷胱甘肽通过植物螯合肽合成酶合成。植物螯合肽是构成裂殖酵母和中、重金属解毒的主要途径,尤其是镉和砷解毒。植物螯合肽可能通过螯合重金属来保护细胞,从而减少重金属毒性。Iinouhe等发现3 种不同的野生型菌株八胞裂殖酵母(Schizosaccharomyces octosporus)、粉状毕赤酵母(Pichia farinosa)和戴尔有孢圆酵母(Torulaspora debrueckii)均对镉有部分抗性,其中八胞裂殖酵母细胞质中镉和植物螯合肽结合为复合物,而粉状毕赤酵母和戴尔有孢圆酵母细胞质中的镉和蛋白结合,类似于酿酒酵母中镉和金属硫蛋白的结合物。这些细胞质中的复合物分子在上述3 种酵母的镉抗性中发挥作用[53]。而Guo Lan等对粟酒裂殖酵母的镉和砷抗性基因进行筛选,发现与镉和砷的抗性均相关的36 个基因中涉及的第3个生物学过程即植物螯合肽生物合成[34]。
3 结 语
虽然生物吸附法作为新型的重金属脱除方法已在去除环境中重金属方面得到广泛应用,但对于具有高吸附、高抗性且广泛应用于食品中的酵母菌的研究尚处于探索阶段,对这类酵母菌重金属解毒的胞内分子机制研究尚不系统和完善。虽然通过新型的各种组学技术和分子机制研究确定了许多与重金属抗性、重金属解毒相关的转录本和蛋白质,然而这些蛋白在胞内受何种转录因子激活以及这些蛋白激活后如何作用于靶点的确切机制并不完全清楚,并且对涉及重金属解毒的各种转运蛋白的翻译后调控了解较少。因此,对于酵母菌抗性、吸附及解毒重金属的机理还应进行更为全面和深入的研究。在后续研究中需从以下方面深入探索:1)进一步筛选对重金属具有良好吸附特性的酵母菌株,建立优势菌种库;2)通过对酵母菌进行组分分离、掩蔽基团及添加能量抑制剂等方法,探究酵母菌对重金属的作用机制主要是胞外的被动吸附还是胞内的主动累积;3)分析酵母菌中的各种转运蛋白在转运、缓解重金属离子带来的氧化应激方面起到的关键作用,以及转运蛋白同重金属结合后如何转运进行外排和细胞器积累及其作用的分子机制;4)对筛选到的对重金属有较强去除作用的优秀菌株的安全性进行探讨和评价,并探究其在宿主体内缓解重金属中毒的机制,以期将其应用于食品、动物和人体中,这在解决环境污染、食品安全危机方面有重要的意义。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
现代养生·上半月(2021年10期)2021-09-24
军事文摘·科学少年(2021年1期)2021-02-04
中国食品学报(2019年10期)2019-11-12
中国药剂学杂志(网络版)(2019年4期)2019-07-17
当代水产(2019年3期)2019-05-14
农产品加工(2019年5期)2019-04-12
天然产物研究与开发(2018年9期)2018-10-08
化肥设计(2017年5期)2017-11-03
天然产物研究与开发(2016年6期)2016-06-05
