
不同替代程度对葎草控制喜旱莲子草光合作用的影响
2019-06-11 05:31代宇雨王力超王一帆张震
安徽农业科学 2019年2期
关键词:光合作用
代宇雨 王力超 王一帆 张震


摘要为了更好地阐释替代控制机制,开展了不同替代程度对葎草控制喜旱莲子草光合作用的影响。将葎草与喜旱莲子草按照4种比例进行种植(3∶3、2∶4、4∶2和0∶6),于2017年8月中旬对喜旱莲子草的光合作用参数进行测定,结果表明,4:2处理喜旱莲子草的净光合速率为单峰现象,净光合速率1 d内均较低,其余处理均为双峰现象,且0:6处理午休现象明显。于2017年6、8和10月晴朗天气对4种处理喜旱莲子草的净光合速率进行测定,3个月中,4∶2处理喜旱莲子草的净光合速率均为单峰现象,且最大峰值出现在10:00,随后逐渐降低。6月,4∶2处理喜旱莲子草的日均净光合速率和最大净光合速率均最大,8和10月4∶2处理的喜旱莲子草日均净光合速率和最大净光合速率较6月迅速降低,显著低于其他处理。对4种处理3个月的喜旱莲子草叶绿素进行测定,除6月2∶4处理喜旱莲子草的叶绿素含量显著低于其他处理外,8和10月各处理间叶绿素含量差异均不显著。
关键词替代控制; 入侵植物; 光合作用
中图分类号S476文献标识码A
文章编号0517-6611(2019)02-0123-04
doi:10.3969/j.issn.0517-6611.2019.02.036
绿色植物通过光合作用使太阳能转化为化学能,同时将大气中的二氧化碳固定下来,形成初级生产量,为生态系统内的能量流动与物质循环提供基础[1]。植物捕捉和利用光能是决定植物生长潜力和适应力的重要因素,光能竞争影响入侵植物的入侵能力[1-2]。随着全球二氧化碳浓度的变化,入侵植物的入侵能力随着改变,这主要表现在入侵植物的光合作用能力[3]。入侵植物对于光的反应有较高的可塑性,但也有研究发现与本地植物相比,入侵植物具有更低的最大光合速率[4]。
光合作用的主要过程包括类囊体电子传递、碳还原周期及气孔导度调控[5]。由于外界环境的变化,导致气孔改变,气孔运动则改变了蒸腾速率,而蒸腾速率的变化又改变了叶片温度和水势等因子的变化[6]。
研究发现,菊芋与三裂叶豚草互作可以降低豚草叶绿素含量,同时降低其光合速率、蒸腾速率等指标[7]。杨晴等[8]研究发现,在黎与黄顶菊的竞争中,黄顶菊的耐阴能力显著低于本地物种黎。因此,本地植物与入侵植物在光能竞争方面的生理生态研究,能够为有效防控入侵植物提供理论依据。
喜旱莲子草(Alternanthera philoxeroides (Mart.) Griseb.)为苋科多年生宿根植物,原产巴西,是一种全球性的恶性杂草[9]。研究发现,葎草(Humulus scandens(Lour.) Merr.)浸提物對喜旱莲子草根茎的生长具有明显的抑制作用,同时野外调查发现葎草与喜旱莲子草共生区后者的盖度明显降低[10]。笔者将葎草与喜旱莲子草不同密度混种,研究葎草替代控制过程中对喜旱莲子草光合作用的影响。
1材料与方法
1.1盆栽试验
试验于安徽农业大学生态学实验室完成,在上口径30 cm、下口径25 cm、高40 cm的桶中种植长势一致的喜旱莲子草和葎草幼苗共6株,分别以3∶3、2∶4、4∶2和0∶6(葎草∶喜旱莲子草)的比例进行种植,每个处理3个重复,分别于2017年6、8和10月中旬晴朗天气进行光合作用参数测定。随机选取每个处理每个重复3片喜旱莲子草与葎草的功能叶进行测定。
1.2光合作用气体交换参数测定
选择晴朗天气,从06:00—16:00 用Li-6400 光合测定仪每隔2 h测定一次,连续测定3 d取平均值,测定参数包括净光合速率(Pn)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、气孔导度(Gs)、光合有效辐射(PAR)、大气温度(Ta)、叶片温度(Tl)、大气相对湿度(RH)和大气二氧化碳浓度(Ca)等。
气孔限制值(Li)采用Berry等[11]的方法,水分利用效率(WUE)采用式(1)计算。
WUE=Pn/Tr(1)
2结果与分析
2.1不同处理对光合特征的影响
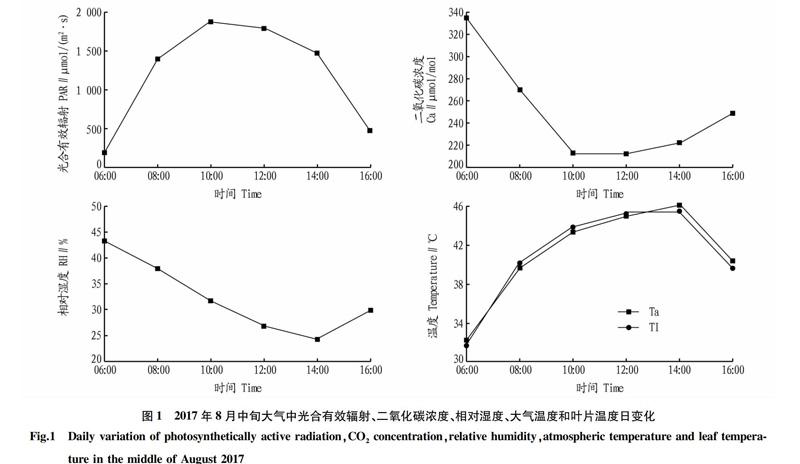
2.1.1气体交换参数日变化。于2017年8月中旬对盆栽周围大气中的PAR、Ca、RH、Ta和Tl的日变化进行测定,结果见图1。由图1可知,随着日变化,PAR逐渐增加,在10:00时达到峰值1 881 μmol/(m2·s),随后逐渐降低。Ta与Tl的变化趋势与PAR相似,仅最高温度出现在14:00,比最大PAR滞后4 h。Ca与RH的变化趋势相似,06:00时Ca浓度最高,随后迅速降低,在10:00时降至最低,随后逐渐升高。RH在14:00时降至最低,随后逐渐升高。
2.1.2光合作用日变化。由图2可知,8月中旬,除4∶2处理外,其余处理喜旱莲子草的Pn均出现双峰现象,2∶4与3∶3处理第1峰值出现在08∶00,0∶6和4∶2处理的第1峰值延后2 h,第2峰值均出现在14:00,谷底出现在12∶00左右,0∶6处理午休现象明显,但4∶2处理仅在10∶00达到峰值,之后Pn明显下降,4∶2处理Pn 1 d内均较低。
4种处理中,除3∶3处理外,其余处理喜旱莲子草Gs均为双峰现象,2∶4处理第1峰值出现在08∶00,4∶2与0∶6处理第1峰值出现在10∶00,第2峰值均出现在14∶00,谷底出现在12∶00左右。Gs随着光强和气温的变化而变化,PAR和Ta均较低,因此Gs也较低,但12∶00时,随着气温与光强达到较高值时,植物出现午休现象,气孔关闭,Gs降低。
Tr与Gs的变化趋势相似,0∶6和2∶4处理均出现双峰现象,分别在10∶00和14∶00,并在12∶00时出现谷值,但3∶3和4∶2处理的Tr在14:00时出现最大值。光强和温度的升高影响了气孔的开放程度,进而增加了内外蒸气压差而影响Tr,由于午间Gs较低,因而Tr也相对较低[12]。
WUE表示植物消耗单位质量的水分所固定的二氧化碳的量,大小取决于二氧化碳净同化速率和蒸腾速率[13]。0∶6与2∶4处理喜旱莲子草的WUE在06∶00时最高,随后逐渐降低,12∶00时降至最低,随后逐渐升高,3∶3与4∶2处理WUE的最高点与最低点较其他处理均延迟2 h。
2.2喜旱莲子草净光合速率季节动态
2.2.1不同时期净光合速率日变化。由图3可知,2017年6月,除4∶2处理外,其余处理均为双峰现象,第1峰值出现在08∶00,第2峰值出现在12∶00,但3∶3处理第2峰值延迟2 h,4∶2处理单峰峰值出现在10∶00。8月,4∶2处理的净光合速率较6月显著降低,除0∶6处理为双峰现象外,其余均为单峰现象,3∶3与2∶4处理的峰值出现在08∶00,其余处理峰值出现在10∶00。10月,4∶2处理喜旱莲子草的Pn日变化趋势与8月相似,10∶00出现峰值,但2∶4处理峰值较8月延后2 h,0∶6处理峰值则较8月提前2 h。
2.2.2日均净光合速率和最大净光合速率的季节变化。由图4可知,不同处理喜旱莲子草的日均Pn和最大Pn在不同季节也不同,这与喜旱莲子草和葎草的生物学特性有关。4个处理的日均Pn和最大Pn变化趋势相似,6月,4∶2处理喜旱莲子草的日均Pn和最大Pn均最大,但随后迅速降低,变化幅度大,8月和10月4∶2处理喜旱莲子草日均Pn和最大Pn显著低于其他处理。
2.3喜旱莲子草叶绿素含量的变化由表1可知,
6月,3∶3处理喜旱莲子草叶绿素a和叶绿素b含量均显著高于其他处理,2∶4处理叶绿素含量则显著低于其他处理。但8月和10月各处理的叶绿素含量差异均不显著。但3∶3处理叶绿素含量较其他处理高,0∶6处理叶绿素含量较其他处理低。4∶2和2∶4处理叶绿素a含量8月高于6月,而0∶6和3∶3处理叶绿素a含量8月低于6月,10月叶绿素a含量均高于6月;叶绿素b含量随着时间逐渐增加。叶绿素a与叶绿素b的比值各处理间差异不显著,但6月与8月混种的喜旱莲子草叶绿素a与叶绿素b的比值较单种低。
3结论与讨论
光是生物生存的能量来源,自然条件下,光强随着时间和空间的变化而改变[12]。除4:2处理外,其他处理均为双峰现象,这说明8月的光强和气温处于较高状态,但4∶2处理中,由于葎草较大的植株与叶片,限制了冠层下喜旱莲子草对光的捕获。植物捕获和利用光能的能力是决定植物生长潜力和适应能力的重要因素,决定入侵植物的入侵能力[2]。植被对于光截获很大程度上受植被内部光分布特征与叶倾角的影响,冠层的形态结构影响植物群落光分布及光能利用率,冠层内水平叶片则能够更加有效地利用光辐射资源,因此,葎草相较于喜旱莲子草有更强的光捕获能力[14-15]。气孔导度能够反映植物气孔张开程度,是衡量二氧化碳和水分进出难易程度的指标,气孔导度的大小反映了植物光合速率、呼吸
速率及蒸腾速率的强弱,是影响光合能力的初始因素[16-17]。与葎草混种处理中,喜旱莲子草的气孔导度显著降低,这可能是由于混种降低了喜旱莲子草对光能的捕获,进而降低了其叶片的羧化能力,Rubisco酶活性降低,从而导致气孔密度降低,最终导致气孔导度降低[18]。研究发现,水分利用率与干物质积累之间存在近似线性关系,干物质的积累则取决于植物对碳的固定[19]。植物种间竞争引起的生理参数的变化可归因于植物对现有资源有效利用能力的差异,尤其是影响叶肉细胞对水分及二氧化碳的有效利用能力[20]。
在混种中,6月喜旱莲子草的光合速率普遍较低,8月由于光强较大,午休现象增强,10月温度降低,使得午休现象被解除。但随着替代时间的延长,4∶2处理喜旱莲子草的日均净光合速率及最大净光合速率显著降低,同时随着替代程度的增加喜旱莲子草的净光合速率也降低。随着光强的逐渐增大,幼嫩的叶片厚度也逐渐增加,光合能力逐渐增加,但成熟的叶片不会改变厚度[21]。但光合速率受多种因素的影响,温度、土壤水分有效性、外界环境空气与内叶环境之间的饱和蒸气压差、植物激素、碳汇强度、污染和养分等[22]。8月由于外界温度过高,导致植物水分蒸发较快,水分成为影响植物光合作用的重要因素。研究发现,在干旱条件下,植物叶绿体膨胀,颗粒层减少,导致光合器官的超微结构受到很大损伤,这严重影响了植物的光合能力[23]。在资源限制的情况下,植物通过调整对资源水平生理反应以及与邻近植物形态的相互作用来增强它们的竞争能力和适应能力[24]。
叶绿素含量与光能吸收、电子传递和转化以及对二氧化碳同化能力具有紧密的关系,高的叶绿素含量反映高的同化效率[25]。混种处理的喜旱莲子草叶绿素含量较高,但除10月外,叶绿素a与叶绿素b的比值均较单种低。研究发现,较高的总叶绿素含量與较低的叶绿素a与叶绿素b的比值表明植物在弱光条件下具有高的光合能力[26]。但叶绿素含量与光合速率间差异的原因仍不清楚,需要继续探讨。
参考文献
[1] FENG Y L,WANG J F,SANG W G.Biomass allocation,morphology and photosynthesis of invasive and noninvasive exotic species grown at four irradiance levels[J].Acta oecologica,2007,31(1):40-47.
[2] STANDISH R J,ROBERTSON A W,WILLIAMS P A.The impact of an invasive weed Tradescantia fluminensis on native forest regeneration[J].Journal of applied ecology,2001,38(6):1253-1263.
[3] WELTZIN J F,BELOTE R T,SANDERS N J.Biological invaders in a greenhouse world:Will elevated CO2 fuel plant invasions?[J].Frontiers in ecology & the environment,2003,1(3):146-153.
[4] NIINEMETS ,VALLADARES F,CEULEMANS R.Leaflevel phenotypic variability and plasticity of invasive Rhododendron ponticum and noninvasive Ilex aquifolium cooccurring at two contrasting European sites[J].Plant cell & environment,2003,26(6):941-956.
[5] ALLEN D J,ORT D R.Impacts of chilling temperatures on photosynthesis in warmclimate plants[J].Trends in plant science,2001,6(1):36-42.
[6] FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annual review of plant physiology,2003,33(3):317-345.
[7] 李建东,孙备,王国骄,等.菊芋对三裂叶豚草叶片光合特性的竞争机理[J].沈阳农业大学学报,2006,37(4):569-572.
[8] 杨晴,李婧实,郭艾英,等.遮阴和藜竞争对黄顶菊光合荧光和生长特性的影响[J].应用生态学报,2014,25(9):2536-2542.
[9] HOLM L,DOLL J,HOLM E,et al.World weeds:Natural histories and distribution[J].Taxon,1997,47(1):209-210.
[10] 张震,徐丽,王育鹏,等.葎草水浸提物对外来入侵植物喜旱莲子草营养生长的影响[J].西北植物学报,2012,32(9):1844-1849.
[11] BERRY J A,DOWNTON W J S.Environmental regulation of photosynthesis[M]//GOVINDJEE.Development,carbon metabolism and plant productivity.New York:Academic Press,1982.
[12] 金则新,李钧敏,马金娥.濒危植物夏蜡梅光合生理生态特性[J].浙江大学学报(理学版),2011,38(6):682-688.
[13] MARSHALL H L,GEIDER R J,FLYNN K J.A mechanistic model of photoinhibition[J].The new phytologist,2000,145(2):347-359.
[14] 黄高宝.作物群体受光结构与作物生产力研究[J].生态学杂志,1999,18(1):59-65.
[15] 董树亭,胡昌浩,岳寿松,等.夏玉米群体光合速率特性及其与冠层结构、生态条件的关系[J].植物生态学与地植物学学报,1992,16(4):372-378.
[16] 宋世佳.施肥模式对棉花生长生理特征及肥料利用率的影响[D].保定:河北农业大学,2011.
[17] QUICK W P,CHAVES M M,WENDLER R,et al.The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions[J].Plant,cell & environment,1992,15(1):25-35.
[18] VINCENT G.Leaf photosynthetic capacity and nitrogen content adjustment to canopy openness in tropical forest tree seedlings[J].Journal of tropical ecology,2001,17(4):495-509.
[19] JENSEN A M,LF M,GARDINER E S.Effects of above and belowground competition from shrubs on photosynthesis,transpiration and growth in Quercus robur L.seedlings[J].Environmental & experimental botany,2011,71(3):367-375.
[20] WANIC M,JASTRZEBSKA M,KOSTRZEWSKA M K,et al.Competition between spring barley(Hordeum vulgare L.) and italian ryegrass(Lolium multiflorum Lam.) under different water supply conditions[J].Acta agrobotanica,2013,66(3):73-80.
[21] OGUCHI R,HIKOSAKA K,HIROSE T.Does the photosynthetic lightacclimation need change in leaf anatomy?[J].Plant,cell & environment,2003,26(4):505-512.
[22] GAYLORD M L,KOLB T E,WALLIN K F,et al.Seasonal dynamics of tree growth,physiology,and resin defenses in a northern Arizona ponderosa pine forest[J].Canadian journal of forest research,2007,37:1173-1183.
[23] CHEN J,DUAN B L,WANG M L,et al.Intraand intersexual competition of Populus cathayana under different watering regimes[J].Functional ecology,2014,28(1):124-136.
[24] ANTEN N P R,CASADOGARCIA R,NAGASHIMA H.Effects of mechanical stress and plant density on mechanical characteristics,growth,and lifetime reproduction of tobacco plants[J].The American naturalist,2005,166(6):650-660.
[25] 張靖梓,柏新富,侯玉平,等.山东半岛沿海防护林入侵植物美洲商陆及其伴生种生长竞争力的比较[J].林业科学,2016,52(3):23-29.
[26] 黄承建,赵思毅,王龙昌,等.马铃薯/玉米套作对马铃薯品种光合特性及产量的影响[J].作物学报,2013,39(2):330-342.
猜你喜欢
科学(2022年4期)2022-10-25
发明与创新(2019年43期)2019-11-16
学苑创造·A版(2018年5期)2018-05-28
现代园艺(2018年3期)2018-02-10
求学·理科版(2016年4期)2016-09-02
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
