
菌根共生与植物入侵
2019-06-19 01:07黄大明尚嘉琛李淑芬AnnePringle
中国科技纵横 2019年8期
关键词:真菌
黄大明 尚嘉琛 李淑芬 Anne Pringle
摘 要:我们对植物入侵能力的影响因素还没有深刻的了解,这涉及到许多机制,不同机制的相对重要性取决于具体的入侵条件。本文讨论菌根共生这一因素。这些共生关系是在大多数陆地生态系统中植物和土壤真菌普遍存在的相互关系。研究植物入侵中菌根共生的概念框架,最重要的几方面包括:(a)菌根状态;(b)入侵植物的生长反应;(c)植物结合不同种真菌的能力;(d)植物作为当地真菌的宿主的能力与反馈动态;(e)真菌生物地理学与分布;(f)真菌的引入与传播;(g)产生新菌根的生态影响。这些因素对植物入侵具有重要影响,因此菌根共生在植物入侵的研究中应受到更多关注。
关键词:丛枝菌根;保护生物学;外生菌根;真菌
中图分类号:S451 文献标识码:A 文章编号:1671-2064(2019)08-0009-07
0 引言
全世界范围内有许多植物被引入新的栖息地,其中一些植物在新的栖息环境中长势茂盛,甚至超出了引进时的预期[55]。植物在新的栖息地中疯长被定义为入侵。一些入侵植物可能会影响当地群落、威胁濒危物种生存[108,32],或对生态系统状态产生影响[79]。当一种入侵植物成为人们的关注目标时,人们便开始设计管理办法以控制其生长。然而,为了设计有效的管理方法,理解入侵过程涉及的机制十分重要。
关于植物入侵的大量文献表明,单一机制无法解释入侵过程。一种植物的入侵能力受到其生态位、竞争能力、繁殖潜力以及这些特性在新栖息地中的演化影响[53,14]。物种的入侵能力同样还受到新栖息地的影响,包括各种新的生物相互作用[65]。食草动物和致病菌是研究这些新生物相互作用的重点,并形成了天敌释放假说的基础[66,97]。然而鲜有工作关注共生作用改变成功侵入的潜在可能但见Richardson等[84]与Mitchell等[65]的综述)。尽管如此,共生关系仍是形成物种生态学的核心力量[38],而且共生关系至少应与寄生关系在外来物种中同样常见[90]。因此,一个物种的引入可导致其它物种的随后侵入[58]。
与真菌形成共生菌根可能有利于植物繁衍。理解共生菌根是否以及如何影响植物入侵,对入侵植物的生态学管理以及当地栖息地的保护十分关键。本文关注大多数植物中常见的菌根共生关系,并讨论这一共生关系如何影响植物的入侵。
菌根是植物的根和真菌组成的共生体。这种关系历史悠久,有化石记录的共生关系可以追溯到460万年前[81]。世界上幾乎95%的植物科有菌根,尽管大部分物种的菌根状态未得到具体的研究证明,但80%鉴定过的陆生植物有菌根[92,98,105]。可参与形成共生体的真菌种类繁多,涵盖了所有真菌的4个门。共生菌根通常被分为丛枝菌根(AM)和外生菌根(EM),其种类由真菌种类和植物的根的结构共同决定[92]。在丛枝菌根和外生菌根之外,还有浆果莓类菌根、水晶兰类菌根、杜鹃花类菌根和兰类菌根等等。菌根通常被认为是一种共生关系,植物和真菌进行物质交换,植物用光合作用得到的碳交换从土壤中获得的氮和磷。菌根带来的其他好处,例如抗病虫害[15]和抗旱能力[3],也被归结于互利共生。与其它共生关系一样,共生成员所得到的好处与具体的共生体状态相关,实际上在某些情况下已成为寄生关系[45,47]。
将植物和真菌形成共生菌根影响植物入侵过程归纳为以下七个方面:
(1)入侵植物的菌根状态。它是否形成了菌根?一些植物不能形成根菌共生体,而且许多入侵植物不与根菌共生。
(2)入侵植物的生长反应。如果植物不能正常形成菌根,那么植物与真菌的共生体是否是必须的,植物是否需要真菌才能发芽和生长?或者说这种共生是否是兼容的?如果是兼容的,共生的植物与本土植物物种相比是否更不容易被感染(疾病)?哪种环境最适宜?
(3)入侵植物结合不同种真菌的能力。植物是否需要与特定的真菌形成菌根共生?或者它是否能与种类多样的不同真菌形成结合体?如果植物需要在新的栖息环境中与当地没有的真菌种类共生,则菌根共生将成为入侵的约束条件。
(4)入侵过程中植物对共生真菌的帮助与在植物入侵中的反馈动态。入侵植物是否是所有真菌或特定真菌的优良宿主?植物和真菌之间相互作用的质量,包括真菌对植物宿主的响应与植物对共生真菌感染的响应,决定了植物-真菌的反馈动态,这能够极大地影响植物入侵的动态过程。
(5)真菌生物地理学与分布。如果每种菌根真菌在地球上均匀分布,那么菌根共生体将不再是植物入侵的约束条件。当然真菌有其分布范围,而且更详尽的真菌的生物地理学认识,对于研究具体情况下需要特定种类真菌的入侵物种十分重要。菌根真菌的分布也与之相关,因为真菌可以独立于引入的植物物种进入新的栖息地。
(6)菌根真菌的引入。最近的研究关注了人类活动导致的菌根真菌的传播;人们在很多情况下将植物完整的根系统与植株一起移植,因此菌根共生体也被打包移植。当菌根真菌随被引入的植物一起被移植到新的栖息环境时,植物更容易存活和传播。被移植到新栖息环境的真菌通常是否是独立于被引入的植物而存活的问题仍悬而未决。
(7)产生新菌根的生态学影响。新真菌的到来与新的植物与真菌的结合是否会影响当地群落及其功能?
在讨论这些问题之前,我们提供了一个植物入侵中关于菌根的概念框架。尽管这个框架被用做关于菌根约束植物入侵的讨论,但框架的部分内容也可以用来理解任何共生体对任何物种入侵的约束。
1 研究植物入侵中菌根作用的概念框架
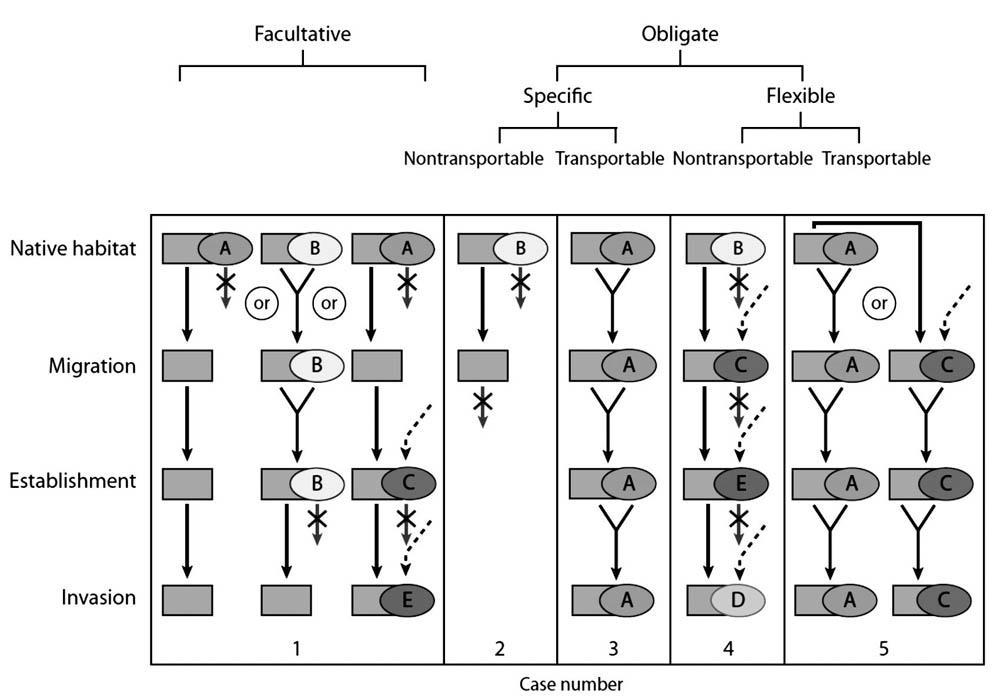
在这一框架中植物根据三个标准分类:(a)是否需要菌根真菌以发芽和生长,专性或兼性;(b)需要特定的真菌或能与多种真菌结合,特定的或多样化的;(c)是与菌根真菌一并被移植还是可以独立移植,菌根结合体是否可以移植(图1)。
每个标准可以结合起来从而对一种植物给出具体的预测,如例1,非菌根或兼性菌根植物(无论该种植物与真菌形成菌根是特定或多样化的,菌根结合体是否可以移植)可以成功入侵(图1)。根据概念框架的预测,共生条件不会阻碍非菌根或兼性生物的入侵。相反,在例2中,专性、形成特定菌根共生体的植物除非与完整的菌根共生体一并移植(例3),否则不能在新的栖息地上生长或传播。专性需要菌根结合的植物如果能与不同种类的真菌结合(例4)或许可以与当地的真菌形成菌根从而成功入侵。最后,在例5中,专性、可以和不同真菌形成菌根的植物与完整的菌根一并移植,很容易成为成功的入侵物种。
根据以上标准可得出以下3个假说:(1)入侵植物常为非菌根植物或兼性共生体。(2)专性依赖菌根共生体的入侵植物,很可能所需菌根真菌的种类多样,并与当地的真菌形成共生体。Richardson等[84]已讨论了与此相关的实验工作;通常的入侵植物对传粉者或其他微生物共生者的需求是泛化的,可以与多样化的共生体结合。Rejmanek[82]也做出了这一预测。(3)专性依赖特定真菌的入侵植物很可能与其完整共生体一并移植,或传播到了合适的共生者已经存在或提前引入的栖息地。
这一框架以及相应举例证明了共生关系和有机体入侵的基本要点:有机体不依赖于共生关系或为兼性共生者有利于有机体入侵,而有机体依赖于专性共生时将阻碍有机体入侵。该物种可形成的共生体多样并与当地生物群互利共生时(这种情况下共生可能促进入侵),或物种与原有共生体一并移植,或移植到已有共生体的地区时,可以减小专性共生对入侵过程的影响。预测引入物种是否会造成生物入侵是重要的思想[30,50,56]。根据这一框架,共生关系对入侵过程的影响可通过生物的兼性、多样化或可转移的共生关系来预测。
本文接下来利用这一框架探讨以上7个主要方面:
(1)入侵植物的菌根状态。有限的数据表明,如框架所预测的,许多入侵植物不需与菌根真菌共生,或利用多样化策略、在环境情况允许时形成菌根,或与不同种类的菌根真菌形成菌根(例如丛枝菌根和外生菌根)。为证明这一结论,我们利用NatureServe[72,73]数据库得到了82种在“美国入侵物种影响排名”(I-Rank)中列为“主要”的维管植物列表。列表中包括82个物种,每个物种的菌根状态根据Wang和Qiu[105]的工作来核对。不出意外,大多数入侵物种的菌根状态仍是未知的。只有33个物种的数据可查到,其中15种与丛枝菌根真菌结合,18种为非菌根植物、兼性菌根植物或可与多样化的真菌形成菌根共生体。
为了解更具有入侵性的植物与不具有入侵性的植物在菌根状态上是否有区别,我对74种I-Rank列为“非主要”的外来物种做了同样的分析。不幸的是其中61种植物在Wang和Qiu[105]的工作中没有相应信息,只有13个物种的数据可查到,其中7种与丛枝菌根真菌结合,其余6种为非菌根的或采用混合策略。尽管数据表明入侵效果小的植物比入侵效果大的植物更可能为专性菌根植物(45%的高风险入侵植物形成专性菌根,在低风险入侵植物中比例为53%),但没有足够的数据证明这一趋势。鉴定植物的菌根状态十分简单,由于菌根真菌能够影响植物入侵过程[104],因此收集入侵植物的菌根结合体数据将会成为大有裨益的研究方向。
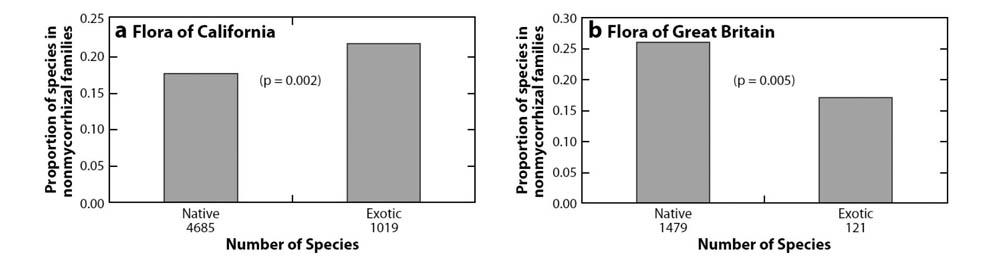
我们也发现有证据表明本地植物种比外来植物种结合菌根真菌的比例更高。我们找到加州的外来植物种与加州的本地植物种的列表[40]。与本地植物相比(图2),加州的归化植物来自常不与菌根真菌结合的科的比例更高[28,74,96,105]。在美国西部的本地植物群中也有类似情况(J.D. Bever,未出版)。在已知的同一科中的菌根状态多样化之外[64,105],这一现象也表明较低的菌根真菌依赖是归化植物成功引入的重要因素之一。然而,这一现象并不适用于英国,与本地植物相比,归化植物来自通常与丛枝菌根真菌共生的科的比例更高[26]。这一差异表明菌根状态在不同地区对植物入侵起到了不同的作用,这一主题我们将在后续部分讨论。
(2)入侵植物的生长反应。植物与菌根真菌共生的反应差异很大。某些情况下植物需要与真菌共生才能生长、繁殖,在离开真菌的条件下不能存活。根据之前的讨论,这些植物是专性共生的。例如许多兰科植物离开合适的真菌甚至无法发育到幼苗期;它们的种子很小,没有充足的碳源储备以供自身生长[92]。许多外生菌根树木在野外也表现为专性依赖于特定的菌根真菌。尽管外生菌根的树苗在理想的实验室条件下能够不依赖菌根良好生长,但在野外其存活率和长势在合适的真菌存在下得以显著提高[62]。然而,大多数植物,尤其是形成丛枝菌根的植物是兼性共生的。它们的确在野外一定会形成菌根共生体,但其生长和繁殖不依赖于菌根。即便无菌根的接种条件下,植物也能生長。然而,它们对接种菌根有生长反应[43]。观察到微小的刺激反应并不罕见,例如加入菌根真菌使植物生物量增加了5-20%。许多情况下促生长效应更显著,当然也可能产生抑制效应。反应程度极大地依赖于环境因素,包括土壤中的营养条件、其它环境参数(例如光)、植物的基因型以及真菌菌株等[45,49,77]。尽管大多数研究在实验室里完成,但在野外也观测到类似结果[77]。
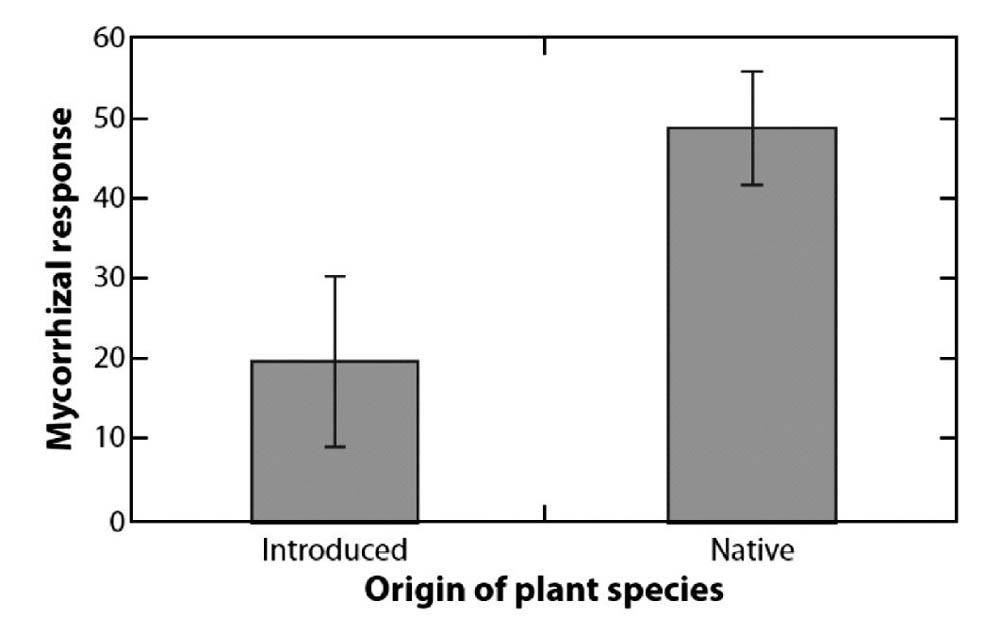
生态学家对在新栖息地和原有栖息地之间的成本和收益平衡仍然缺乏了解,记录由菌根真菌在原有和被引入的区域取得的收益的资料很少。然而,仍有实例与入侵植物种对菌根真菌依赖性小的趋势相符。例如对堪萨斯的96种植物的菌根反应的再分析表明,当地的植物对菌根真菌的接种响应比引入植物更显著,在植物科属和长期控制下也是如此(图3)。进一步研究这些植物中的某一对时,Hetrick&Wilson[39]证明接种响应的差异并非由菌根真菌的来源产生,表明所得结果是由菌根响应中一种普遍的差异导致的。在南加州的植物中也发现引入植物相对于本地植物响应较小的现象(J.D.Bever,未发表)。
原产欧洲与引入北美的St.Johns wort(贯叶连翘)的菌根依赖性对比表明,在植物入侵过程中选择压力使植物对菌根真菌的依赖下降[89]。与欧洲的贯叶连翘种群相比,北美种群对丛枝菌根接种的响应显著减小,相应地根系更细。尽管这一现象与加州地区入侵植物种中非菌根植物比例更高的结果一致,却不与英国的结果相符(图1),而且体现了菌根真菌在北美入侵过程相对于欧洲更加重要的不对称性,因而需要进一步的工作验证这一现象。
(3)入侵植物结合不同真菌的能力。许多菌根植物可以与多种真菌形成菌根结合体,当植物可以与当地的真菌种类结合时,入侵将更有可能成功。例如在实验室条件下,大多数结合丛枝菌根真菌的植物似乎可与大多数丛枝菌根真菌结合。而实际上,一些菌根植物只能与一种或几种丛枝菌根真菌形成高度专一性的共生体[36],这些专一性的结合是否普遍存在还不知道。
入侵性的Centaurea maculosa(矢车菊)是一例表面上与丛枝菌根真菌专性共生却能与多种真菌形成共生体的植物;因为C. maculosa能与当地的丛枝菌根真菌共生。共生关系似乎不能成为其入侵北美西部的约束条件。在原产地和入侵地,丛枝菌根真菌侵染C. maculosa的比例都很高[59]。丛枝菌根真菌共生体可通过水平传播,于种子在土壤中发芽时重新建立共生体;C. maculosa的种子传播时,会暴露于新种类的真菌。C. maculosa可以同似乎是当地种类的丛枝菌根真菌共生,而且能利用这些真菌的优势与当地的草类竞争[59]。这一情况下,引入的草类植物与当地真菌的共生有助于植物的入侵。其他可以同多种丛枝菌根真菌共生的植物包括北卡罗来纳草原从兼性(Anthoxanthum odoratum,黄花茅)到专性(Allium vineale,鸦蒜)依赖丛枝菌根真菌的引入植物[9]。这些植物均与多种本地丛枝菌根真菌共生[9,11,77]。
相反,形成外生菌根的植物在共生体的专一性上差异更大。尽管部分植物能与多种真菌共生(据记载超过2000种真菌可与Pseudotsuga menziesii(花旗松)共生;Molina et al[68]),但其它物种更为专一,只能与一个或几个属/种的真菌结合。例如桤木属的物种高度专一地与少数几个物种共生(Molina[67])。这种情况下共生关系将成为入侵的障碍,除非植物与共生真菌一并移植,或被移植到已有所需真菌的地方(见下文第5点)。
(4)入侵过程中植物对菌根真菌的帮助以及反馈动态。入侵植物与当地菌根真菌的相互作用,以及植物和真菌群落间的动态反馈,影响着植物入侵的过程和效果。一种植物的引入可能改变菌根真菌群落的密度和/或组成,对该植物的传播可能产生正反馈或负反馈作用[8,12,10]。例如引入的非菌根植物药用蒜芥抑制了当地丛枝菌根真菌和外生菌根真菌的生长,进而提高了它与当地物种的竞争能力[17,94,110]。类似地,在加州草原,丛枝菌根真菌的种群密度在归化植物群落的支配下受到抑制,进而抑制了高度依赖丛枝菌根的本地植物的生长[104]。在这两个例子中,丛枝菌根真菌密度的变化产生的正反馈促使群落向多稳态发展。丛枝菌根真菌密度抑制将导致群落更易受其他归化物种入侵,进而引发入侵崩溃[90]。
菌根真菌群落的组成变化可能会抑制植物入侵。例如归化的兼性宿主Plantago lanceolata(长叶车前),在当地Panicum sphaerocarpon(球形黍)聚集的真菌存在下长势良好,但会受到自身聚集的丛枝菌根真菌的抑制[7]。在这种情况下,此负反馈动力阻止了P. lanceolata完全替代当地的植物品种。
菌根真菌群落的组成变化也可能加剧入侵。这种可能性得到了几种引入加拿大的植物获得菌根真菌群落正反馈的支持[48]。这种效应在C. maculosa的入侵中或许也是一个重要因素,因为在C. maculosa根系中的丛枝菌根真菌组成与本地植物根系中的组成不同[69],当二者共培养时,邻近C. maculosa的本地植物根系的真菌组成发生了改变[70]。在美国西部和非洲大草原上的入侵草类中也观察到了类似变化[34,100]。
(5)真菌生物地理学与分布。所有真菌的分布和生物地理学由形态特征、生态特征和婚配系统决定,形态特征限制其传播过程,生态学特征决定栖息环境是否适合生存,婚配系统决定了种群的建立和耐受性。过去人们认为微生物可以在世界范围内分布:微生物能够在全球传播,环境因素限制了在某个特定地点的种群建立[5]。然而当生物学家利用分子生物学手段描绘和研究微生物物种的分布时,细菌和真菌具有按地理组织的种群结构,而且遵循距离隔离模式[61,95]。遵循这类生物地理学分布的真菌包括Pleurotus spp.(侧耳属)[103]和Schizophyllum commune(裂褶菌)[42]等腐生真菌和Rhizopogon spp.(须腹菌属)[31,52]、Russula brevipes(短柄红菇)[6]和Tricholoma scalpturatum(雕文口蘑)[18]等外生菌根真菌。进一步地,对全球分布的利用形态学定义的腐生物种进行多基因测序,其基因系谱往往呈现多样化,种系差异大而形态相似的物种的亲缘关系也比较远,例如Neurospora(脉孢菌)[24],Saccharomyces paradoxus(奇异酵母)[51]。
实际上,过去根据形态学定义的外生菌根物种也被发现寄宿方式多样、亲缘关系远,且这些物种呈现出洲际差异性,可能是扩散限制的结果。例如一项全球范围内对假松露属Pisolithus(豆马勃属)的调查发现,之前被认为是一个物种的P. tinctorius(彩色豆马勃),实际上是11个亲缘种形成的复合种[60]。以上现象均具有受限的地域分布,例如P. tinctorius仅分布在北半球,P. marmoratus仅分布在大洋洲。类似地,外生菌根的捕蝇蕈的多基因位点的分子生物学证据表明,这一过去被认为是一种广泛分布的物种实际上由8个亲缘种组成,在北美(clade I)和欧亚大陆(clade II)分别形成2個受地域限制分布的进化分枝[27]。
与独立生活的真菌相反,菌根真菌的生物地理学和种群结构受到了宿主专一性和宿主植物地域分布结构的影响[33,107]。对Tricholoma matsutake(松茸)复合种的植物地理学研究表明,其现在的生物地理学分布是它沿着其松柏类的宿主,向西穿过白令陆桥形成的[19]。P. marmoratus在大洋洲的地理分布限制在桉属植物的范围,即使其他外生菌根植物与桉属植物同时存在也是如此。这一模式被认为是由于劳亚古陆和冈瓦纳古陆的分离产生的地理隔离导致的[25,60]。宿主专一性也是生态多样性导致物种形成的潜在机理[29]。在Paxillus(桩菇属)中关系密切的两种外生菌根真菌(P. involutus and P. filamentosus)具有重叠的地理分布,但结合的宿主大不相同,P. filamentosus几乎仅以桤木属植物为宿主,而P. involutus的宿主种类广泛[44]。宿主专一性或许参与形成了这两个物种的差异。
对特定环境条件或栖息地类型的适应很可能对决定菌根真菌基因型和物种的现有分布模式至关重要。土壤化学,包括营养物质浓度[4,54,75]和蛇纹岩发育土壤中的重金属浓度[86]或例如采矿等人类活动[20]都影响着菌根真菌的群落组成。在捕蝇蕈复合种中,在不易受扩散限制的较小的地理距离上,仍有证据表明其分布模式具有种间的栖息地偏好[27]。
综上所述,在长的时间尺度上的地质事件,例如冈瓦纳古陆与劳亚古陆的形成与扩散限制,宿主偏好以及栖息地专一性都是影响菌根真菌生物地理学分布模式的因素。然而,现有与这些生物的多样性及生态学意义相关的研究数量很少,全球范围内利用多基因序列數据研究丛枝菌根真菌物种的研究几乎没有。作为古老、无性繁殖,具有大孢子的广布类群,丛枝菌根真菌与外生菌根真菌差异巨大,作为极有价值的参照,可帮助深入理解以上因素对菌根真菌的生物地理学分布与种群结构的多样化影响。我们的最终目标是全面了解植物分布、土壤因素和地理起源在菌根真菌分布中的决定性作用。对决定菌根真菌分布因素的相对重要性有了更清晰的认识,我们可以更准确地预测植物进入新栖息地找到合适、有益的菌根共生体的可能性。
(6)菌根真菌的引入。当真菌独立引入新栖息地,或植物连同完整的根系被一并移植时,即使是高度专一的共生关系也不会影响植物的传播。尽管最近有综述关注菌根接种体携带真菌引入新的栖息地[88]以及全球范围内外生菌根真菌的引进[101],但真菌很少被当作引入或入侵物种讨论[23]。
真菌个体通常隐藏在土壤或其它基质中。蘑菇的个体十分明显,可成为地面的风景,例如剧毒的外生菌根真菌Amanita phalloides(毒伞蕈)在世界各地均有分布[78]。尽管该物种原产于欧洲,现已被引入澳大利亚、新西兰、非洲、美洲等地;在加州甚至入侵了沿海森林[76]。广义的外生菌根A. muscaria也很明显、易辨识;这些就是童话中经典的带白色圆点的红蘑菇。这一复合种原产北半球,现已传播到澳大利亚、新西兰、非洲、夏威夷、南美等地[13,25,37,91,101,102]。在新西兰和澳大利亚,A. muscaria入侵了当地的假山毛榉林[41,46,93,101,111]。
专性依赖于特定真菌或某一类菌根共生体的引入植物,可在它的共生伙伴被独立引入之后具备入侵能力。尽管A. phalloides和A. muscaria似乎都是广泛分布的物种,但能与这些真菌共生、获得特定利益的植物更易在真菌引入后才入侵。由丛枝菌根真菌主导的外生菌根植物宿主的传播也可以从A. phalloides和A. muscaria的引入中受益。这些真菌入侵当地森林,使引入植物可以随之进入当地森林。已有至少200种外生菌根真菌被引入世界各地[101];它们会影响引入植物在当地的传播。
这一逻辑应当被优先考虑。真菌的引入通常由土壤的搬运完成,而土壤经常被运输于非洲各地、帮助造林(图4)。这些土壤显然会携带许多种真菌到本地区外的大洲,这些植物的生长和传播会受限,直到这些真菌被引入。例如Pinus patula(展叶松)在外生菌根共生体单独引入之前,无法在赞比亚生长。在新栖息地中,真菌能够帮助植物入侵[85]。南非地区的松树也有类似的情况[85]。
此外,当植物连同完整的根系被一并移植时,即使是高度专一的共生关系也不会影响植物的传播。赤杨树似乎只能与少数几种真菌形成菌根结合体(Molina[67]),但赤杨树已被成功引入了世界上很多地区,因此我们认为这些树木是与菌根共生体一并移植的。或许赤杨树对真菌共生体的选择性比过去所观察到的更加广泛。
(7)群落与生态学影响。菌根真菌不仅影响植物个体的表现和植物群落的组成;它们也是生态系统的组成部分。真菌对土壤团聚、营养物质循环、初级生产以及碳储存等诸多过程都有已知或潜在的影响(Rillig[83])。但是大多数情况下菌根对某一过程速率的贡献比例人们尚不清楚[99]。菌根可以通过以下四个不同的途径(Rillig[83])影响生态系统过程速率,每一途径都要通过与入侵植物的相互作用完成:菌丝的直接影响(例如土壤团聚),生态系统内对宿主植物的影响(例如营养物质循环),对土壤微生物群落的影响,以及对植物群落组成的影响(例如初级生产)。不同类型的菌根结合体(包括非菌根植物,丛枝菌根和外生菌根)被认为通常沿不同的轴向排列,例如根据矿物质获取的模式被划分为水平、纵向或连续的模式,菌根对植物的帮助也被认为与此相符[80]。外生菌根和丛枝菌根具有不同的营养循环特点,外生菌根(和欧石南属)真菌可以从有机物中获得养分,而丛枝菌根偏好矿化的养分[1]。因此区分同种菌根的入侵和不同种菌根入侵的生态学影响十分有用。
第一种情况是同种菌根类型的入侵,例如丛枝菌根植物或真菌入侵了丛枝菌根结合主导的生态系统。植物宿主倾向于在根际聚集一部分当地丛枝菌根群落(如,Bever等[9])。实际上,Mummey和Rillig等[70]发现蒙大拿州一片被C. maculosa入侵的草原具有不同于未被入侵的邻近地区的丛枝菌根真菌群落。入侵植物也可以改变邻近本地植物根系中的丛枝菌根真菌组成[34,70]。有两个原因可能使真菌组成的改变对植物的反馈响应尤为重要。如第4点所述,与本地植物相比,真菌群落的改变可能使入侵植物直接获益(植物-真菌正反馈作用)。第二,真菌组成的改变也可能在生态系统水平上产生间接的效果。发生改变的真菌群落可能有助于改变营养物质丰度、其它物化性质或土壤微生物群落,其相互作用可能会促进进一步的入侵。例如发生改变的真菌群落由于功能或数量增加可使土壤中可吸收的养分急剧下降,实际上会改变本地植物群落成员间的竞争强度。另一个例子是,改变了的丛枝菌根真菌群落可能影响土壤的团聚结构、稳定性以及碳储存,从而通过改变持水能力、离子交换能力或不同的养分淋失水平,间接影响本地植物。改变后的丛枝菌根真菌群落也会影响其它土壤微生物的丰度和组成(例如磷助溶剂,致病菌等),对营养物质循环产生功能性影响。
某种菌根类型的宿主向另一种菌根类型主导的生态系统入侵也会导致以上所述的生态效应,但影响可能会更为显著,因为菌根类型与不同的植物和生态系统过程紧密相关[1,21,80]。Alliaria(葱芥属)在北美枫树林的入侵便是非菌根植物入侵丛枝菌根主导的森林的例子[17,94]。这一入侵会导致丛枝菌根真菌对树苗发芽的帮助减少,减弱丛枝菌根真菌调控的生态系统功能。这种植物也会影响外生菌根真菌的丰度[110]。因此暂时的结论是(如第4点所述),非菌根类杂草可以使生态系统向非菌根状态发展,有益于未来非菌根(或菌根依赖低)植物入侵。其它例子包括外生菌根树木向丛枝菌根真菌主导的草原的入侵。由于外生菌根真菌也能够营腐生生活,可能获取了更多的有机营养物质导致有机养分在循环中的短缺[87]。另一种重要情况包含了新的真菌类群本地真菌群落的潜在入侵风险,即商用的真菌接种物被用于农业或园艺中;这一实践很广泛却缺乏相应的研究[88]。正如Maherali和Klironomos[57]所展示的那样,入侵真菌对真菌群落的组成和功能的影响很可能取决于入侵者与目标群落的系统发育关系。实际上Antunes等[2]的工作表明,Glomus intraradices(根内球囊霉)这种丛枝菌根真菌的引入,对已经含有该种真菌的丛枝菌根真菌群落影响很小。但有了更易得的跟踪个体真菌在环境中活动的工具[22],鉴于许多菌根真菌可商品化生产并被传播到世界各地,商用真菌接种体的使用仍需进一步研究。
本综述集中讨论了菌根共生体和植物入侵的生态学影响。然而,也有许多主要的经济影响与这些现象相关,本文提供了一些实例,但并未详细讨论。Mura等[71]认为欧洲的佩里戈尔黑松露(Tuber melanosporum,法国黑孢块菌),一种外生菌根真菌共生体,将会被十分相似的中国松露Tuber indicum(印度块菌)所取代;在经济效应之外,也会对烹饪造成可怕的影响。在马达加斯加,当地生长在原始森林中的外生菌根真菌产量无法满足商业需求。相反,在占据中部高原大部地区70%总土地面积的桉树林中,食用蘑菇的产销量巨大。这些与新的宿主共生的蘑菇并不是澳大利亚桉树林的本地物种,可能为马达加斯加原有或其它地区引入的物种。不论这些真菌的来源如何,新的共生关系为马达加斯加人提供了重要的食物和经济来源。
2 结语
综上所述,共生根菌分布广泛,且在陆地生态系统中,真菌根菌通常能与多种植物结合。本文讨论了共生根菌对植物入侵过程可能产生限制或帮助的几个要点,尤其是入侵植物是否需要根菌共生体、植物与真菌结合的专一程度,以及合适的真菌是否与植物一并移植或先行独立引入新的栖息地。在这些真菌成功引入的基础上,入侵植物被其自身的根菌共生体影响(同时也影响着周围的菌根共生体),进而影响入侵过程并对当地群落产生影响。
参考文献
[1] Aerts R. 2002. The role of various types of mycorrhizal fungi in nutrient cycling and plant competition. In Mycorrhizal Ecology, ed. MGA van der Heijden, IR Sanders, pp. 117-33. Berlin: Springer Verlag.
[2] Antunes PM, Koch AM, Dunfield KE, Hart MM, Downing A, et al. Influence of commercial inoculation withGlomus intraradices on the structure and functioning of an AM fungal community from an agriculturalsite. Plant Soil. 317:257-66.
[3] Aug'e RM. 2001.Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 11:3-42.
[4] Avis PG, McLaughlin DJ, Dentinger BC, Reich PB. 2003. Long-term increase in nitrogen supply altersabove- and below-ground ectomycorrhizal communities and increases the dominance of Russula spp. Ina temperate oak savanna. New Phytol. 160:239-53.
[5] Baas-Becking LGM. 1934. Geobiologie of Inleiding Tot de Milieukunde. The Hague: WP van Stockum & Zoon.
[6] Bergemann SE, Miller SL. 2002. Size, distribution, and persistence of genets in local populations of the latestage ectomycorrhizal basidiomycete Russula brevipes. New Phytol. 156:313-20.
[7] Bever JD. 2002. Negative feedback within a mutualism: Host-specific growth of mycorrhizal fungi reducesplant benefit. Proc. R. Soc. London Ser. B 269:2595-601.
[8] Bever JD. 2003. Soil community feedback and the coexistence of competitors: conceptual frameworks andempirical tests. New Phytol. 157:465-73.
[9] Bever JD, Morton JB, Antonovics J, Schultz PA. 1996. Host-dependent sporulation and species diversity ofarbuscular mycorrhizal fungi in a mown grassland. J. Ecol. 84:71-82.
[10] Bever JD, Pringle A, Schultz P. 2002. Dynamics within the plant-arbuscular mycorrhizal fungal mutualism:testing the nature of community feedback. In Mycorrhizal Ecology, ed. MGA van der Heijden, IR Sanders,pp. 267-94. Berlin: Springer Verlag.
[11]-[112]文獻略.
猜你喜欢
学苑创造·A版(2022年11期)2022-05-30
科学(2020年2期)2020-08-24
小学科学(学生版)(2018年8期)2018-08-21
现代园艺(2017年23期)2018-01-18
天然产物研究与开发(2016年11期)2016-06-15
微生物与感染(2015年5期)2015-12-08
大豆科技(2015年1期)2015-03-23
营销界(2015年23期)2015-02-28
食品工业科技(2014年9期)2014-03-11
华东理工大学学报(自然科学版)(2014年1期)2014-02-27
