
细胞壁降解酶在辣椒尾孢霉致病过程中的作用
2019-07-02 10:37孙俊
中国瓜菜 2019年2期
关键词:辣椒


摘 要: 以辣椒尾孢霉(Cercospora capsici Heald. et Wolf.)为供试菌,采用分光光度计测定辣椒尾孢霉产生的细胞壁降解酶种类;通过电导率测定及透射电镜观察以探索细胞壁降解酶的致病作用。结果表明,辣椒尾孢霉在活体内外均可产生多聚半乳糖醛酸酶(PG)、果胶甲基半乳糖醛酸酶(PMG)、多聚半乳糖醛酸反式消除酶(PGTE)、果胶甲基反式消除酶(PMTE) 和羧甲基纤维素酶(Cx) 等细胞壁降解酶,而β-葡萄糖苷酶只在活体外产生,且该菌在活体内和活体外产生的细胞壁降解酶活性差异较大。透射电镜观察结果表明,细胞壁降解酶对叶片的超微结构有破坏作用,病菌细胞壁降解酶在致病过程中能够损伤寄主的细胞膜,从而影响其透性;就细胞膜损伤率而言,混合酶的作用显著高于单一酶,果胶酶对细胞膜的损伤明显强于纤维素酶。
关键词: 辣椒; 辣椒尾孢霉; 细胞壁降解酶; 致病机制
The role of cell wall degrading enzymes in the pathogenic process of pepper brown leaf spot disease caused by Cercospora capsici
SUN Jun
(Liaoning Institute of Economic Forestry, Dalian 116031, Liaoning, China)
Abstract: The species of cell wall degradation enzyme (CWDE) produced by Cercospora capsici was determined by spectrophotometer. The cell wall degradation enzyme involved in pathogenicity was observed by measuring electrical conductivity and transmission electron microscopy. It showed that Cercospora capsici can produce polygalacturonase(PG), polymethyl- galacturonase(PMG), polygalacturonic acid trans- eliminase (PGTE ), pectin methyl trans-eliminase (PMTE) and carboxymethyl cellulase (Cx) both in vitro and in vivo, while β-glucosidase was only found in vitro.The cell wall degradation enzyme (CWDE) produced by Cercospora capsici were different between in vivo and in vitro.The results of transmission electron microscopy showed that the cell wall degradation enzyme damaged to the ultrastructure of the leaves.The host cell membrane can be damaged during the pathogenic process, thus affecting its permeability. In terms of cell membrane injury rate, the effect of mixed enzyme was significantly higher than that of single enzyme, and pectinase was significantly stronger than cellulase.
Key words:Pepper;Cercospora capsici; Cell wall degrading enzymes; Pathogenic mechanism
辣椒褐斑病为辽宁地区蔬菜生产上的新病害,致病菌为辣椒尾孢霉(Cercospora capsici Heald. et Wolf.),主要危害叶片,严重时病斑相连成不规则的大斑,后期病组织干枯坏死,呈挣裂状穿孔,致使叶片支离破碎,严重时病叶变黄脱落[1]。目前,對于辣椒褐斑病的报道仅限于症状描述及病原菌的鉴定,其致病机制尚不明确。植物病原菌在与寄主相互识别的过程中可产生一系列细胞壁降解酶,降解植物组织以提供营养并促进其对寄主组织的入侵[2-3]。因此,细胞壁降解酶是植物病原真菌的重要致病因子[4-5]。了解细胞壁降解酶在尾孢霉致病机制中的作用,对进一步有针对性地控制病害具有重要意义。植物病原菌产生的细胞壁降解酶对寄主植物的伤害往往是果胶酶和纤维素酶共同作用完成的。陈晓林等[6]发现,苹果树腐烂病菌Valsa ceratosperma在活体组织内、外均能分泌一系列的细胞壁降解酶——多聚半乳糖醛酸酶(PG)、果胶甲基半乳糖醛酸酶(PMG)、纤维素酶、β-葡萄糖苷酶和木聚糖酶。赵景楠等[7]研究了草莓褐色叶斑病菌产生的细胞壁降解酶的种类、培养条件及致病机制,证明其在该病原菌的致病过程中起重要作用。车喜庆等[8]为了明确人参不同部位对人参核盘菌的敏感性,在温室分别对人参芦头部和须根部接种病菌,观察不同部位发病情况,并对人参不同部位细胞壁降解酶活性及寄主防御酶活性进行测定。
由于病菌产生的细胞壁降解酶是病菌侵入寄主的主要手段之一,因此明确辣椒褐斑病菌产生的细胞壁降解酶种类及活性,对深入研究其致病机制具有重要意义。鉴于此,笔者研究了辣椒褐斑病菌产生的细胞壁酶种类及活性,进一步阐明辣椒褐斑病菌的致病机制,旨在为阐明病菌生长与环境条件的关系以及进一步探讨病害的发病规律及防治方法提供理论基础。
1 材料与方法
1.1 细胞壁降解酶的提取与纯化
试验于2014年7月在沈阳农业大学植物病理真菌实验室和仪器分析中心进行。按Papavizas和Ayers的方法将分离纯化的辣椒尾孢菌(Cercospora capsici)在Marcus[9]或Richard细胞壁降解酶诱导培养基上培养。病菌培养9 d及15 d 后,过滤除去菌丝和孢子,在4 ℃下10 000 r·min-1离心15 min,上清液即为酶粗提液。向酶粗提液中加入硫酸铵到60%饱和度(25 ℃),在4 ℃下先静置5 h后再以10 000 r·min-1离心15min,弃上清液,用50 mmol·L-1醋酸-醋酸钠缓冲液(pH 5.0)分别溶解沉淀。沉淀在同样的缓冲液中于4 ℃下透析2 d,每12 h换1次透析液,纯化的酶在-20 ℃下保存备用。分离纯化的辣椒尾孢菌菌株在PSA上培养25 d后,打取菌片接种到辣椒叶片上,置于人工气候箱中高湿培养,取样并提纯[10],以未接种的辣椒叶片作对照。
1.2 细胞壁降解酶活性的测定
参照张红、吴洁云等 [11-12]的方法采用紫外-可见光分光光度计法测定细胞壁降解酶的活性。
1.3 辣椒褐斑病菌细胞壁降解酶致病作用
1.3.1 对辣椒叶片作用的观察 选取4~5叶期辣椒叶片(‘沈椒4号),针刺叶片造成伤口,分别将浸有纯化的病菌细胞壁降解酶和经高温(121 ℃)处理后的病菌细胞壁降解酶的脱脂棉覆盖在伤口上,置于28~30 ℃下培养并保湿,24 h后观察叶片的病变情况并拍照,以蒸馏水处理作对照。
1.3.2 对辣椒叶片细胞膜损伤的测定 将辣椒叶片用直径5 mm的打孔器制成若干块,以去离子水冲洗晾干,置于分别装有5 mL酶液的PC管中,减压抽气使叶片沉入酶液中,处理后测定叶片组织浸出液的电导率[11-12]。
1.3.3 对辣椒叶片细胞超微结构破坏的观察 切取辣椒叶片2 mm组织块若干,分别浸入装有酶液的PC管中,取样,脱水。用JEM-1200EX(JEOL日本电子)透射电子显微镜观察组织结构的变化。对照用灭菌水处理36 h [11-12]。
2 结果与分析
2.1 病菌在活体内外产生细胞壁降解酶的种类及活性
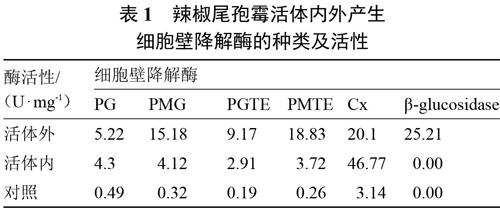
辣椒尾孢菌在活体内外均能产PG、PMG、PGTE、PMTE和Cx,而β-葡萄糖苷酶只在活体外产生,在活体内不产生。该病菌在活体外产生细胞壁降解酶的活性依次为β-葡萄糖苷酶、Cx、PMTE、PMG、PGTE和PG;在活体内产生细胞壁降解酶的活性依次为Cx、PG、PMG、PMTE和PGTE。产生的纤维素酶活性高于果胶酶。除Cx外,其余5种细胞壁降解酶活体外的产生量均高于活体内。结果还表明,接种病菌后,病组织内各种细胞壁降解酶的活性明显高于未接种的健康对照植株(表1)。
表1 辣椒尾孢霉活体内外产生
细胞壁降解酶的种类及活性
[酶活性/
(U·mg-1) 细胞壁降解酶 PG PMG PGTE PMTE Cx β-glucosidase 活体外 5.22 15.18 9.17 18.83 20.1 25.21 活体内 4.3 4.12 2.91 3.72 46.77 0.00 对照 0.49 0.32 0.19 0.26 3.14 0.00 ]
2.2 辣椒褐斑病菌细胞壁降解酶的致病作用
由图1可以看出,细胞壁降解酶处理的辣椒叶片出现了不同程度的水渍状病斑和黄色斑块,而酶液经高温(121 ℃)处理后则未能侵害叶片。病菌接种后的叶片褐化严重,病斑近圆形,而酶液处理叶片以黄化为主。果胶酶和混合酶对叶片的破坏作用较大。
2.3 对辣椒叶片细胞膜损伤的测定
由表2可知,辣椒褐斑病菌产生的细胞壁降解酶对辣椒叶片细胞膜有明显的损伤作用。混合酶的作用极显著高于单一酶,细胞膜损伤率最大,达到51.33%;果胶酶对细胞膜的损伤极显著强于纤维素酶。(注:纤维素酶Cx活性为21.10 U·mg-1;果胶酶PMG活性为15.18 U·mg-1)
2.4 对辣椒叶片细胞超微结构破坏的观察
如图2-A所示,对照叶片细胞超微结构中细胞壁结构完整,叶绿体膜结构完整,片层结构清晰、排列有序。纤维素酶处理36 h后,叶片细胞叶绿体脱落,叶绿体膜消解,线粒体膜模糊(图2-B),用果胶酶处理的叶片,超微结构发生了明显变化,细胞壁结构受损、消解,细胞膜破裂,内含物外泄,叶绿体膜破裂,基粒片层模糊,线粒体膜受损,内含物外泄,线粒体嵴模糊、消失(图2-C)。比较果胶酶与纤维素酶对叶片细胞的破坏作用,果胶酶消解细胞内含物的作用更强。混合酶处理叶片后,细胞壁变形扭曲、变薄,线粒体嵴消失,线粒体膜模糊,细胞内细胞器消失,叶绿体膜消解,基粒片层结构崩解(图2-D)。
3 讨论与结论
辣椒尾孢菌在活体内、外均能产PG、PMG、PGTE、PMTE和Cx,而β-葡萄糖苷酶只在活體外产生,在活体内不产生,且该菌在活体内和活体外产生的细胞壁降解酶活性差异较大,这可能是由于活体内和活体外环境因素不同,导致酶活性差异较大。活体外酶活性分析时,应多考虑温度、pH值、底物浓度等对酶活性的影响,但活体内产生的酶接近自然发病情况,具有实际研究意义。研究发现,PGTE和PMTE培养9 d时活性最高,而PG 、PMG、Cx和β-葡萄糖苷酶培养15 d时活性最高,因此PGTE和PMTE在培养9 d测定,而PG、PMG、Cx和β-葡萄糖苷酶在培养15 d测定;PMG活体外培养基选用的是Richard培养液,其他酶选用改良的Marcus培养液[13]。
细胞壁降解酶处理的辣椒叶片出现不同程度的水渍状病斑和黄色斑块,而酶液经高温(121 ℃)处理后则失去对叶片的侵害作用,酶液处理的叶片与病菌接种的叶片症状相似。由此推测,细胞壁降解酶是该病原菌致病因子之一。果胶酶、纤维素酶以及两者的混合酶液,可使叶片出现明显病斑,其中以果胶酶和混合酶对叶片的破坏作用较大。在叶片没有伤口的情况下,酶液处理后产生的症状较轻,但在叶片有伤口的情况下用酶液处理,则产生明显黄化和水渍状病斑,这与金勤等[14]的研究结果一致。同样杨媚等[15]研究认为,通过酶处理叶片后发现,细胞壁降解酶确实对水稻叶片组织造成损伤,并且随着酶浓度升高,损伤程度逐渐加重。
随着电镜技术的发展,很多学者更深入地研究了病原菌致病因子对寄主组织细胞的破坏。李宝聚等[16]通过透射电镜和扫描电镜观察,指出果胶酶、纤维素酶可以造成植物细胞质壁分离,还可造成叶绿体、线粒体内部空泡化,液泡、内质网受损。研究证明,叶绿体膜、片层结构主要被纤维素酶降解,线粒体膜主要被果胶酶降解。辣椒褐斑病菌产生的细胞壁降解酶对辣椒叶片细胞膜有明显损伤作用。通过透射电镜观察,细胞壁降解酶对叶片的超微结构有破坏作用。酶液能造成辣椒叶片细胞质壁分离、细胞壁消解、细胞质膜断裂、叶绿体膜破裂、基粒片层模糊、线粒体膜受损、内含物外泄甚至消失等现象,该结果与张红[11]对立枯丝核菌细胞壁降解酶致病作用的研究结果一致。
提高寄主细胞壁抗降解酶能力可能是寄主抗褐斑病的有效途径之一。由于辣椒尾孢菌可以产生一系列细胞壁降解酶,而且这几种细胞壁降解酶是该菌的主要致病因子之一,因此增强寄主细胞壁抗降解酶能力是在生产中提高寄主抗病性的可行措施之一,可在今后的抗病育种过程中進一步深入研究。
参考文献
[1] 孙俊,刘志恒,黄欣阳,等.辣椒褐斑病菌生物学特性研究[J].植物保护,2009,35(5):109-113.
[2] HEMATY K,CHERK C,SOMERVILLE S.Host-pathogen warfare at the plant cell wall[J].Current Opinion in Plant Biology,2009,12(4):406-413.
[3] CANTU D,VICENTE A R,LABAVITCH J M,et al.Strangers in the matrix:Plant cell walls and pathogen susceptibility[J].Trends in Plant Science,2008,13(11):610-617.
[4] YANG Y Q,YANG M,LI M H,et al.Cloning and functional analysis of an endo-PG-encoding gene Rrspg1 of Rhizoctonia solani,the causal agent of rice sheath blight[J].Canadian Journal of Plant Pathology,2012(3):43-47.
[5] 李明海,杨迎青,杨媚,等.井冈霉素对水稻纹枯病菌细胞壁降解酶活性和可溶性蛋白产生的影响[J].华中农业大学学报,2010,29(3):272-276.
[6] 陈晓林,牛程旺,李保华,等.苹果树腐烂病菌产生细胞壁降解酶的种类及其活性分析[J].华北农学报,2012,27(2):207-212.
[7] 赵景楠,马喆,耿文龙,等.新病害草莓褐色叶斑病病原菌细胞壁降解酶研究[J].中国农学通报2014,30(31):261-266.
[8] 车喜庆,傅俊范,李自博,等.人参核盘菌侵染对参根不同部位细胞壁降解酶及防御酶活性的影响[J].河南农业科学,2017,46(10):66-70.
[9] MARCUS L,BARASH I,SNEH B,et al.Purification and characterization of pectolytic enzymes produced by virulent and hypovirulent isolates of Rhizoctonia solani Kuhn[J].Physiological and Molecular Plant Pathology,1986,29(3):325-336.
[10] 李宝聚.温室黄瓜、番茄主要病害生态控制的应用基础研究[D].沈阳:沈阳农业大学,2005.
[11] 张红.立枯丝核菌胞壁降解酶及其在致病中的作用[D].江苏扬州:扬州大学,2004.
[12] 吴洁云.灰葡萄孢胞壁降解酶、角质酶及其对番茄植株的致病作用[D].江苏扬州:扬州大学,2007.
[13] 张懿,台莲梅,左豫虎.马铃薯早疫病病菌产生细胞壁降解酶的条件研究[J].中国植保导刊,2016,36(6):12-16.
[14] 金勤,周国英,刘君昂,等.细胞壁降解酶在油茶炭疽病菌致病过程中的作用研究[J].植物保护,2017,43(3):97-102.
[15] 杨媚,杨迎青,郑丽,等.水稻纹枯病菌细胞壁降解酶组分分析、活性测定及其致病作用[J].中国水稻科学,2012,26(5):600-606.
[16] 李宝聚,周长力,赵奎华.黄瓜黑星病菌致病机理的研究Ⅲ细胞壁降解酶和毒素对寄主超微结构的影响及协同作用[J].植物病理学报,2001,31(1):63-64.
猜你喜欢
领导文萃(2022年13期)2022-07-13
快乐作文(1.2年级)(2020年8期)2020-09-10
作文评点报·作文素材初中版(2016年29期)2016-08-18
37°女人(2016年5期)2016-05-06
风流一代·经典文摘(2011年1期)2011-07-15
现代妇女(2009年12期)2010-02-25
金点子生意(2009年8期)2009-09-29
感悟(2009年7期)2009-08-17
今日文摘(2009年1期)2009-08-11
故事家·风吟(2009年2期)2009-02-13
