
福建三明东牙溪流域南方红豆杉自然种群结构与动态研究
2019-07-12 07:53张莉
安徽农学通报 2019年11期
张莉
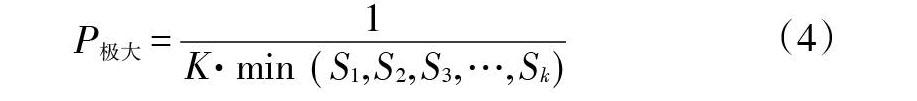
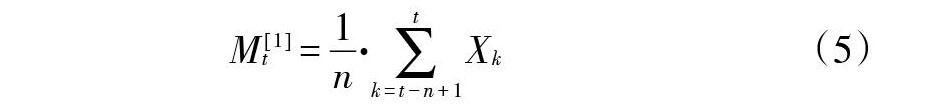
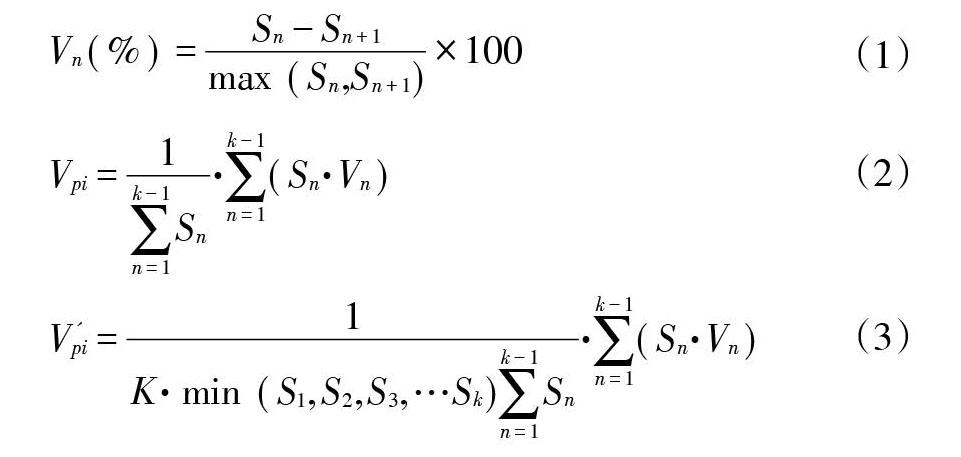
摘 要:以福建三明东牙溪流域南方红豆杉为对象,研究了南方红豆自然种群结构,编制了种群静态生命表,分析了存活曲线、死亡率曲线和寿命期望曲线,并運用时间序列模型预测种群数量动态。结果表明:南方红豆杉种群幼苗幼树较多,达72.24%,只有17.90%的幼苗能长成小树,个体数到第Ⅳ级时达到最低;个体平均寿命期望在第Ⅳ~Ⅶ级间最大,种群结构稳定;维持保护好现有种群,东牙溪流域南方红豆杉种群依然有恢复壮大潜能。
关键词:南方红豆杉;种群结构;生命表;存活曲线;时间序列
中图分类号 S791.49文献标识码 A文章编号 1007-7731(2019)11-0057-5
Abstract:Based on Taxus chinensis var. Mairei in the Dongyaxi Basin of Sanming,Fujian. The natural population structure of the T. chinensis var. Mairei was studied. The static life table of the population was compiled,the survival curve,the mortality curve and the life expectancy curve were analyzed,the time series model was used to predict the population dynamics. The results showed,that there were more seedlings and saplings in the T. chinensis var. Mairei population,reading 72.24%,only 17.90% of the seedlings could grow into small trees,and the number of individuals reached the lowest when they reached the size class Ⅳ;the average life expectancy of individuals reached the maximum in the size class Ⅳ to the size class Ⅶ,the population structure is stable;maintaining and protecting the existing population,the population of the T. chinensis var. Mairei in the Dongyaxi Basin still has the potential to recover.
Key words:Taxus chinensis var. Mairei;Population structure;Life table;Survival curve;Time series
植物种群结构体现了种群大小和内部不同个体的分布状况,反映了种群内个体间的组配情况和种群与环境的相互关系[1],生命表、存活曲线和时间序列模型都是研究植物种群结构及种群动态变化的重要工具[2]。通过研究珍稀濒危植物天然种群的生命表、存活曲线及时间序列模型,可以分析种群结构现状、演替动态及其与环境的竞争关系,对制定珍稀濒危植物保护利用策略具有十分重要的意义[3-5]。
南方红豆杉(Taxus chinensis var. mairei)属红豆杉科、红豆杉属,是国家一级保护珍贵植物[6,7]。为耐荫树种,寿命长,生长缓慢,主要生长在海拔1500m以下的山谷溪流边[8],常以小种群或散生木形式散布在天然阔叶林、毛竹林内。在福建山区,常有南方红豆杉大树生长在村落周围的景观风水林内,有时甚至还是主要的建群树种。目前,对于南方红豆杉种群结构的研究,主要集中在连续森林中的种群[9,10]。研究表明,南方红豆杉幼苗早期阶段死亡率较高,但当达到一定龄级后,存活率则相对稳定。对于村落周围的南方红豆杉种群,刘佳庆[11]等对比研究了皖南仙寓山、浙江天目山和福建梅花山3地村落周围的天然种群结构及其动态变化。而目前对于小流域范围内南方红豆杉天然种群结构和动态的研究尚未见报道。
福建省三明市东牙溪是闽江上游沙溪一级支流,源于三明市三元区中村乡莲花顶,其下游建有东牙溪水库,系三明市区主要饮用水源,供水量占市区饮用水的95%以上。东牙溪流域面积18791hm2,其中有林地17005hm2,毛竹林12632hm2,分别占流域面积、有林地面积的67.22%和74.28%。目前,流域内仍然广泛散布的南方红豆杉大树,以及山上遗存有大量尚未完全腐烂的南方红豆杉树蔸,均表明南方红豆杉曾经在东牙溪流域大量分布。由于人类长期经营活动和过度采伐利用,致使当地南方红豆杉数量大量减少。为此,笔者开展了东牙溪流域南方红豆杉天然种群结构及其动态变化研究,对于在饮用水源地科学制定珍稀濒危树种的保护策略,提高饮用水源地的生态安全具有重要意义。
1 研究地点及其自然概况
东牙溪流域位于三明市三元区中村乡,地理坐标为东经117°36′2″~117°45′21″,北纬26°3′7″~26°15′50″。属亚热带湿润气候,光照充足,四季分明,年平均气温19.5℃,最低气温-6.5℃,最高气温40.6℃,多年平均降雨量1563mm。东牙溪流域属中低山地貌,切割深,高差大,相对落差在700~900m,有普禅山(1510m)、莲花峰(1502m)、张公岩(1471m)、蓬仙岩(1319m)等10余座海拔在1000m以上高山。土壤主要为花岗岩发育的红壤和黄红壤,土层厚度多在80cm以上。现有红豆杉主要以小种群或散生木形式散布在村落周围的景观风水林、毛竹林内,远离村庄的天然阔叶林中也有部分分布。
2 研究方法
2.1 野外调查方法 2017年9—12月,借助当地村民和平时了解到的情况,在对东牙溪流域南方红豆杉种群进行充分普查的基础上,選择在南方红豆杉集中分布处设置30m×30m的样地。调查样地内的乔木树种及其株数,对南方红豆杉进行每木检尺(起测胸径≥5.0cm),记录其树高、胸径、冠幅等指标,对胸径<5.0cm的幼苗、幼树,按树高1.0m为级差,分别统计株数。共计设置调查样地30个。
2.2 种群结构特征
2.2.1 径级、高度级划分 对于难以获取年龄的树种,许多学者在分析种群结构特征时,采用径级结构代替龄级结构,取得了良好的效果[12-14]。南方红豆杉是国家一级保护树种,不能随意采伐,且不少野生植株胸径超过30cm,难以确定准确年龄,因而采用径级结构代替年龄结构来分析其种群结构特征。参考李先琨等[1]的划分标准,结合南方红豆杉的生长特性,将南方红豆杉种群个体划分为6个径级:Ⅰ级,幼苗(S1),个体高度H≤100.0cm;Ⅱ级,幼树(S2),H>100.0cm,胸径D<5.0cm;Ⅲ级,小树(S3),5.0≤D<10.0cm;Ⅳ级,中年树(S4),10.0≤D<20.0cm;Ⅴ级,成年树(S5),20.0≤D<30.0cm;Ⅵ级,大树(S6),D≥30.0cm。高度结构以2m为级差,也分为6个高度级。
2.2.2 结构特征分析 分别统计各径级和高度级南方红豆杉的株数,以径级和高度级为横坐标,以株数为纵坐标,绘制南方红豆杉种群径级和高度级结构图。采用陈晓德[15]的量化方法对种群动态进行定量描述,计算公式如下:
式中,Vn表示种群从n到n+1级的株数变化动态,Vpi表示整个种群结构的数量变化动态指数,P为种群受外界干扰的风险概率,Sn、Sn+1分别表示第n级与第n+1级的种群个体数。Vpi与Vn为正、负、0时,分别反映相邻2级种群个体数量的增长、衰退、稳定的动态关系。只有当P值最大时,才对种群动态Vpi构成最大影响。
2.2.3 静态生命表和存活曲线 生命表是判断植物种群发展变化趋势的重要指标之一,静态生命表多用于长寿命木本植物种群统计研究。通过将各龄级存活个体数进行标准化,计算各龄级发展到下一龄级时的死亡数、死亡率、致死力和期望寿命,编制植物种群静态生命表,绘制存活曲线,以分析出种群现实状况和植物种群与环境竞争关系[1,13,16]。具体计算公式:标准化存活数Ix=aX/a0×1000,标准化死亡数dx=lx-lx+1,死亡率qx=dx/lx,存活数Lx=(lx+lx+1)/2,寿命期望值ex=Tx/lx,致死力kx=[n]x-[n]x+1。其中a0为各龄级中最大存活个体数,ax为x龄级存活个体数,Tx为x龄级及以上龄级个体总数和;[n]x为标准化存活数的自然对数值。本文在南方红豆杉径级划分标准的基础上,并假设不同年龄南方红豆杉每年的胸径生长量相同,综合样地数据对胸径大于5.0cm的个体,按10cm的级差编制静态生命表,绘制存活曲线。
2.2.4 时间序列预测模型 时间序列分析是一种综合研究时间数列预测和回归预测的方法,该方法侧重于根据时间序列以往的变动规律推测今后的发展趋势[17],可用于预测多种复杂条件下种群变化趋势。本文利用一次移动平均法,分别对未来2个龄级和4个龄级后的南方红豆杉种群数量进行时间序列预测[18]。计算公式如下:
式中,n表示需要预测的未来龄级数量,Mt[1]表示未来n个龄级后t龄级的种群大小,Xk为当前k龄级的种群大小。
3 结果与分析
3.1 种群分布现状和结构特征 野外调查发现,在整个东牙溪流域,南方红豆杉分布在海拔350~1300m的丘陵低山地区,主要是海拔700~1100m范围内。大树多分布在村头水尾等村庄周围,呈斑块状分布,坑源村牛架山、埔头城庄山头、居阳村同心墓等地,均有大树片林集中分布。在远离村庄居住区的林分内,较少有南方红豆杉大树分布。本次调查发现,胸径大于30cm的红豆杉大树有300多株;最大的1株位于中村乡大培坑村,胸径196cm;高度0.5m以下的小规格幼苗较多,但高度1m以上胸径5cm以下的幼树较少。
东牙溪流域南方红豆杉种群结构详见图1。从图1可以看出:组成南方红豆杉种群的个体数,首先随径级和高度级的增加而减少,径级和高度级在第Ⅳ级时,个体数都达到最低,而后个体数又逐渐增加。同时,南方红豆杉种群中幼苗幼树较多,中老年个体较少,其中幼苗(S1)和幼树(S2)个体数各占整个种群的52.83%和19.41%,两者合计达72.24%,中年树(S4)仅占4.42%,整个种群数量随高度级和径级的变化趋势基本一致,均呈反“J”型,表明南方红豆杉种群结构虽然有波动,种群数量也不多,但目前还属稳定型种群。根据量化分析方法进行种群动态分析,南方红豆杉种群相邻各级个体数量变化动态指数V分别为63.26%、60.13%、42.86%、5.26%和-57.30%,整个种群径级结构动态指数Vpi为47.03%,大于0,受随机干扰时种群年龄结构动态指数Vpi′为0.26%,也大于0,随机干扰风险极大值即种群结构对随机干扰的敏感性指数P为0.00556,表明种群结构表现相对稳定,对外界干扰比较敏感。
3.2 静态生命表和存活曲线 静态生命表(表1)和存活曲线(图2)表明:南方红豆杉种群的幼年个体数量明显高于后面几个阶段,种群从幼苗到幼树、幼树到小树生长阶段,数量急剧下降,分别有36.70%、48.77%能顺利成活生长,只有17.90%的幼苗能顺利长成小树。本次调查发现,在村头水尾的南方红豆杉片林调查样地的主林层下有较多红豆杉幼苗幼树,而在毛竹林调查样地内基本没有幼树小树。南方红豆杉成年植株有的不结实,会结实的一般结实量都比较大,且种子有后熟现象。红豆杉果实味甘甜,色红艳,鸟类松鼠喜食。鸟类松鼠食用果实后,排泄出不消化的种子散布在母树周围,遇到合适的生境就会萌发生长,因此幼苗数量丰富。本次调查发现,毛竹林调查样地内红豆杉幼苗、幼树数量稀少,应该与村民经营管理毛竹林劈灌除草时将南方红豆杉幼树一并劈除有关。本次调查也发现有1个样地的地上残留有很多红豆杉种子,但种壳已发黑,咬开发现种仁已干枯,失去活性,无法正常发芽生长,这可能与该样地所处的小环境有关。
南方红豆杉种群死亡率、致死力和寿命期望曲线(图3)显示,南方红豆杉种群死亡率qx和致死力Kx曲线变化趋势一致。致死力Kx在第Ⅰ级、第Ⅴ級和第Ⅸ级出现峰值。第Ⅰ级的峰值显示了幼苗长成幼树的高死亡率;第Ⅴ级的峰值显示了红豆杉长成大树(胸径25cm)后数量大幅减少,可能与20世纪90年代兴起的毛竹丰产培育热,毛竹林内混生乔木被采伐有关;第Ⅸ级的峰值显示植株个体接近生理衰老,形成死亡高峰。
寿命期望ex表明第x龄级个体的生存能力。南方红豆杉种群平均寿命期望从第Ⅰ级到第Ⅳ级,一直都是逐步增大,在第Ⅳ、Ⅵ、Ⅶ级达到最大且基本保持稳定,显示当地环境十分适宜红豆杉生长,同时也表明南方红豆杉从第Ⅳ~Ⅶ级是生长旺盛期,生存质量最高。在第Ⅴ级形成1个低谷的原因,应该与造成该级高致死力的原因相同。从第Ⅷ后,种群平均寿命期望值逐步下降,表明南方红豆杉种群的生理活性逐步衰退。
3.3 时间序列预测 南方红豆杉种群结构时间序列分析详见表2。由表2可知:经过2个龄级的发展,南方红豆杉种群一直到第Ⅵ级,各龄级数量均呈增长趋势;第Ⅶ、Ⅷ、Ⅸ级均保持相对稳定的数量。经过4个龄级的发展,南方红豆杉种群数量一直到第Ⅷ级,各龄级均呈增长趋势,直到第Ⅸ级才略有下降。目前南方红豆杉苗期更新旺盛,幼苗幼树数量较多,整个种群维持在比较充足的后备资源,再经过2个、4个龄级的发展,整个种群仍处于不断壮大。因此,从种群动态预测结果来看,只要不人为破坏,维持好现有种群,东牙溪流域的南方红豆杉种群能逐渐壮大。
4 结论与讨论
目前,东牙溪流域南方红豆杉主要分布在人类活动频繁的村头水尾、毛竹林内。受人类生产经营活动的高强度人为干扰,过去在东牙溪流域广泛存在的南方红豆杉,分布日益破碎,形成相互隔离的小斑块。
本文在对东牙溪流域南方红豆杉种群内各斑块植株数量进行调查统计的基础上,编制静态生命表,分析其存活曲线和死亡率、致死力和寿命期望曲线,反映了该珍稀濒危树种种群的基本属性。结果表明:南方红豆杉种群数量虽然不大,但幼年个体数量比较丰富,中年个体数量较少,成年个体数量相对比较稳定。其原因如下:(1)南方红豆杉成年植株结实量大,果实经鸟类松鼠食用后,促进了种子后熟,有利于种子发芽,长成大量众幼苗幼树;(2)东牙溪流域的生态环境十分适宜南方红豆杉生长,历史上该流域曾经有大量南方红豆杉分布也映证了这点;(3)近20多年来,当地森林资源保护力度较大,特别是近年东牙溪流域被划为饮用水源保护区,保护力度持续增强,滥砍乱伐林木现象大幅减少,因此幼苗数量丰富。此外,当地林农收入主要依靠毛竹林经营,毛竹经营活动强度较高,每年都要通过刀劈、割灌机割除甚至施用除草剂等各种措施去除杂灌,不可避免清除了大量红豆杉幼苗幼树,影响了种群数量,这在一定程度上迟滞了南方红豆杉种群的繁衍更新。
种群动态预测结果也显示,减少人为破坏,维持保护好现有种群,东牙溪流域南方红豆杉种群依然具备恢复发展和壮大的潜能。特别是划为饮用水源保护区后,森林资源保护力度持续增强,保证了南方红豆杉成年植株的正常生长发育和开花结实,保证了种群的更新和发展。如果能进一步推广毛竹林生态经营技术,尽可能保留毛竹林内的幼苗、幼树,甚至在毛竹林下人工套种红豆杉幼苗、幼树,进一步扩大种群数量,对促进东牙溪流域南方红豆杉种群的恢复十分有利。
参考文献
[1]李先琨,苏宗明,向悟生,等.濒危植物元宝山冷杉种群结构与分布格局[J].生态学报,2002,22(12):2245-2253.
[2]洪伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[3]谢宗强,陈伟烈,路鹏,等.濒危植物银杉的种群统计与年龄结构.生态学报,1999,19(4):523-528.
[4]马姜明,刘世荣,史作民,等.川西亚高山暗针叶林恢复过程中岷江冷杉天然更新状况及其影响因子[J].植物生态学报,2009,33(4):646-657.
[5]吴明开,沈志君,刘海,等.梵净山自然保护区珙桐天然种群生命表与生存分析[J].生态学杂志,2012,31(6):1419-1424.
[6]费永俊,雷择湘,余昌均,等.中国红豆杉属植物的濒危原因及可持续利用对策[J].自然资源,1997(5):59-63.
[7]岳红娟,仝川,朱锦懋,等.濒危植物南方红豆杉种子雨和土壤种子库特征[J].生态学报,2010,30(16):4389-4400.
[8]胡忠俊,赵娟.江西三清山南方红豆杉的种群结构与分布分析[J].安徽农学通报,2018,24(21):120-122,143.
[9]李先琨,向悟生,苏宗明.南方红豆杉无性系种群结构和动态研究[J].应用生态学报,2004,15(2):177-180.
[10]Shi G L,Bai B,Lu C H.Seed rain and seed bank of Chinese yew (Taxus chinensis var. mairei) population in Tianmu Mountain[J]. Acta Ecol Sin,2010,30(5):276–279.
猜你喜欢
运动(2017年1期)2017-09-03
绿色科技(2016年23期)2017-03-15
广西农学报(2015年4期)2015-09-11
