
外源赤霉素对GAR 小麦光合特性影响的初探
2019-08-17 06:48张宁宁严加坤
山西农业科学 2019年8期
张宁宁,鱼 慧,严加坤
(1.榆林学院生命科学学院,陕西榆林719000;2.榆林市南郊农场,陕西榆林719000)
赤霉素作为一类对植物生长发育具有一定调节作用的激素,其生理功能可以概括为:促进生长、打破休眠、影响性别分化、促进开花、改善品质和增加抗性等[1-3]。目前赤霉素在生理作用方面研究比较透彻的是促进生长方面。
当前,赤霉素在调节植物抗性方面的研究定论不一。在毛白杨中,利用外源赤霉素处理显著提高了其叶片净光合速率、气孔导度、叶片蒸腾及树高等生理指标,表明在水分正常条件下,外源赤霉素对于提高植物生长具有一定的促进作用[4]。相似的研究结论在玉米中也有报道,如研究发现,GA3显著提高了干旱条件下玉米的抗性[5]。然而,在大豆研究中,盐胁迫下,外源赤霉素显著增加了内源活性赤霉素(GA1和GA4)及茉莉酸甲酯的含量,但其降低了内源脱落酸及水杨酸的含量,而后2 种物质是植物应答非生物胁迫的重要调控激素[6-8]。虽然外源赤霉素缓解了盐胁迫带来的生物量降低,但其缓解盐胁迫中是否具有正效应还有待验证。韩德复[9]研究发现,在不同程度盐碱化草地上生长的羊草叶片赤霉素含量显著低于其在无盐碱化草地羊草叶片含量,而中度盐碱化草地羊草叶片脱落酸含量显著高于无盐碱化草地含量。说明羊草在长期适应盐碱化的过程中形成了内源激素调控机制。同时也说明降低内源赤霉素含量有利于羊草适应非生物胁迫。
赤霉素敏感型(GAR)矮秆基因如Rht8 和Rht13在旱地小麦育种中已经得到广泛的应用,研究发现,赤霉素敏感型矮秆基因Rht13 在水分正常的条件下使小麦具有比rht 品系更长的旗叶;还发现Rht13 和Rht8 品系具有比rht 品系更高的光合速率。而GASPERINI 等[10]研究发现,Rht8 基因并没有降低内源活性赤霉素的含量。但是,赤霉素对Rht13及Rht8 的光合速率影响机制以及赤霉素对小麦抗旱性的影响尚不清楚。
本研究通过盆栽称重控水试验研究了2 种水分条件下矮秆基因Rht13、Rht8 及其对照rht 基因对小麦抗旱性的影响,旨在揭示外源赤霉素与净光合速率之间的关系及其机制。
1 材料和方法
1.1 材料
以Rht8 基因供体晋麦47 与Rht13 基因供体Magnif M1为亲本进行杂交得到F1;F1自交产生F2,经过分子标记鉴定共得到F2群体296 个;之后进行连续自交,并且以生长周期、株高等性状为标准进行筛选,得到F6纯合度较高的重组自交系株系;选取矮秆基因材料Rht13 株系1 个、Rht8 株系1 个及不含矮秆基因rht 株系1 个进行后续试验。
1.2 方法
试验在人工气候室内进行。采取完全随机设计,试验用盆为高30 cm、直径15 cm 的乳白色PVC管制盆。每盆装有杨凌本地塿土5 kg、石子0.5 kg。培养条件为光照12 h,光照强度400 μmol/(m2·s),白天温度25 ℃,夜间温度18 ℃,湿度维持在50%~60%。
消毒的种子经过4 ℃春化24 h,之后在20 ℃催芽48 h 至种子露白。选择大小均一的种子,每盆点种6 粒。在三叶期开始定苗3 株,六叶期开始为期28 d 的春化处理,温度5 ℃,湿度70%±5%,光照时间12 h、光照强度400 μmol/(m2·s)。在春化结束之前,田间持水量维持在75%±5%,春化结束后开始进行干旱胁迫,胁迫组田间持水量维持在35%±5%。外源赤霉素(100 μmol/L GA3)按照GASPERINI的方法喷施[11]。共4 个处理,即灌水处理(WW)、灌水+赤霉素处理(WW+GA)、干旱处理(DR)及干旱+赤霉素处理(DR+GA)。
1.3 指标测定及方法
在外源赤霉素处理20,35 d 分别使用Li-6400光合仪测定完全展开的新叶净光合速率。同时使用SPAD-502 叶绿素仪读取叶片相对叶绿素含量。叶片大小使用直尺测量并记录。之后,取样测定干物质量。
1.4 数据统计分析
试验结果采用算数平均数±标准方差表示。利用SPSS 19.0 数据分析软件进行统计分析,采用LSD 进行显著性分析。作图及相关关系拟合采用Sigmaplot 12.5 软件进行。
2 结果与分析
2.1 外源赤霉素对GAR 小麦生物量相关指标的影响
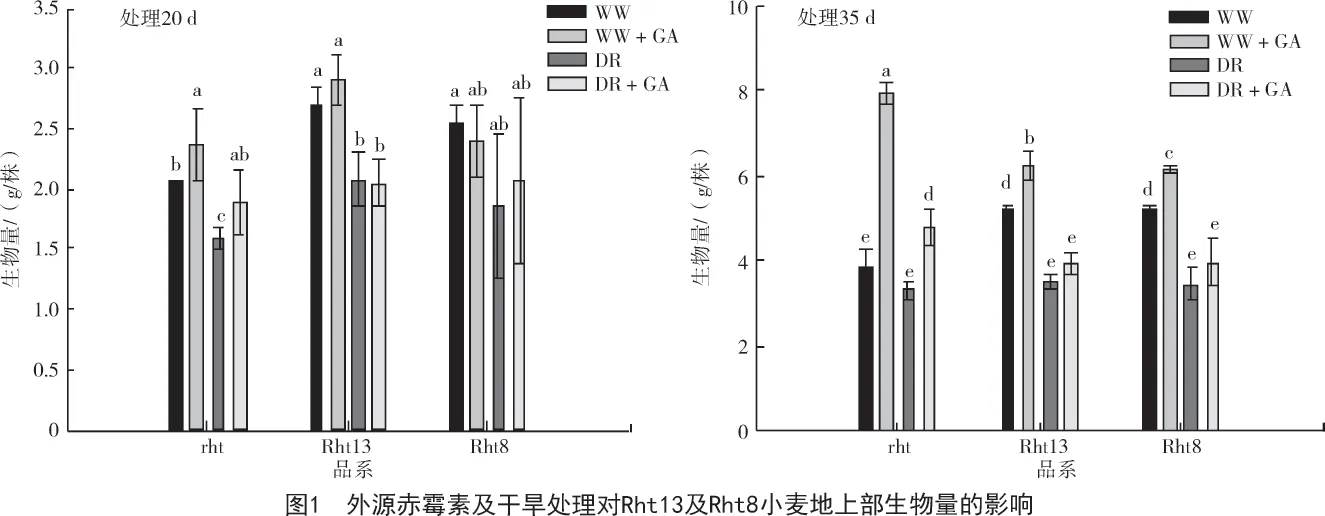
赤霉素作为农业上广泛应用的提高作物叶片伸长的调节物质,其对作物生物量的改良方面已经得到证实[12]。由图1 可知,喷施赤霉素35 d 后,2 种水分条件下,所有在测品系地上部生物量均显著提高(P<0.05);但是处理20 d,赤霉素对生物量的增加效果并不显著。rht 品系在外源赤霉素的作用下,地上部生物量的增加幅度比Rht13 和Rht8 小麦大的多,然而在没有外源赤霉素的条件下,赤霉素敏感型矮秆基因小麦的地上部生物量对照组显著高于rht 品系。这与前人发现的Rht8 矮秆基因在小麦生长早期对小麦没有影响的结果存在差异[13],可能是由于该研究结论是在统计了胚芽鞘长度、叶伸长速率及株高后得出的,并不能全面代表苗期小麦的生理指数。另外,试验设计及实施条件的不同对矮秆小麦的研究结果也有一定的影响[11]。
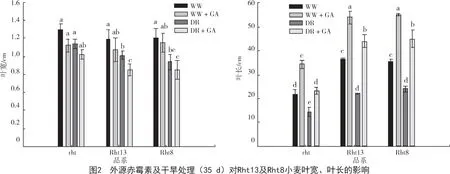
从图2 可以看出,赤霉素处理组和非赤霉素处理组小麦在叶宽方面并不具有显著性差异,但有降低的趋势。并且在干旱条件下,非赤霉素处理组赤霉素敏感矮秆基因品系的叶宽相比对照品系rht 显著降低,灌水组虽然叶宽没有显著降低但呈现出降低的趋势。赤霉素处理显著增加了Rht13、Rht8 及rht 品系的小麦叶长(P<0.05),尤其是赤霉素敏感的Rht13 和Rht8 品系;而在非赤霉素处理组,赤霉素敏感的Rht13 和Rht8 品系的叶片长度显著高于rht 品系(P<0.05),结果与大田试验得到的结果及前人研究结果[14]一致。
2.2 GAR 小麦对外源赤霉素的光合响应
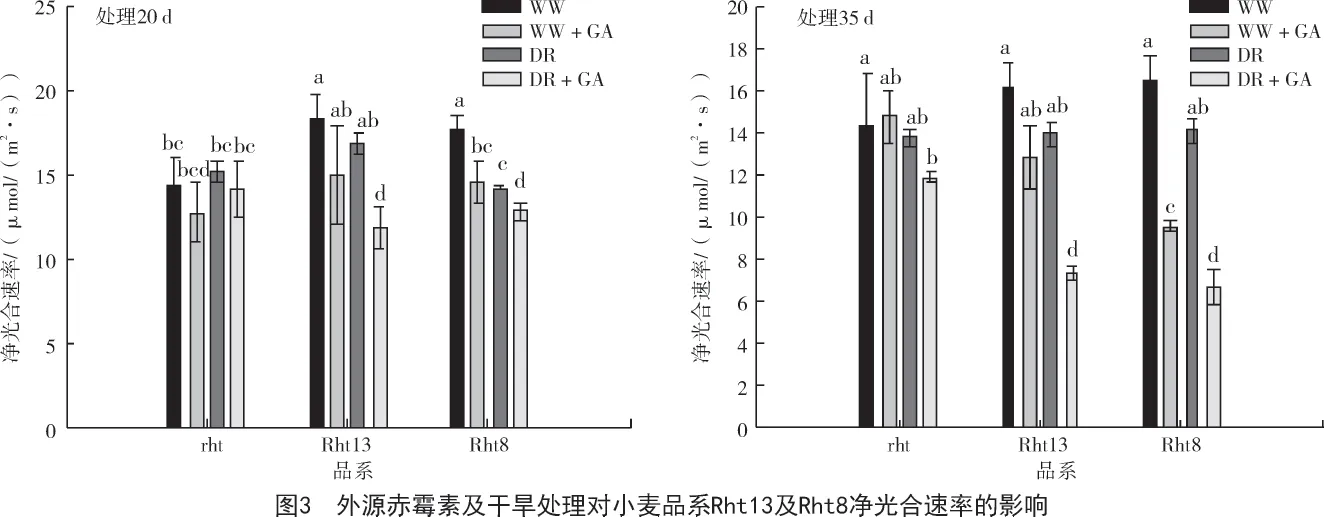
进一步分析处理后净光合速率的变化将有助于清晰地认识赤霉素施加后Rht13 和Rht8 品系相比rht 品系降低了地上部生物量的原因。从图3 可以看出,在赤霉素处理20 d,灌水条件下,仅有Rht8品系光合出现显著性降低(P<0.05),而Rht13 和rht 品系开始呈现出降低的趋势(P>0.05);干旱条件下Rht13 和Rht8 均出现净光合速率显著降低(P<0.05)。在处理后第35 天,灌水处理中,外源赤霉素显著降低了Rht13 和Rht8 的净光合速率,而rht 品系净光合速率并没有比非赤霉素处理组降低。在干旱组,赤霉素处理显著降低了Rht13 和Rht8 品系的净光合速率(P<0.05),而rht 品系的净光合速率相比Rht13 和Rht8 要高。
2.3 叶绿素变化及其与其他生长指标的相关关系
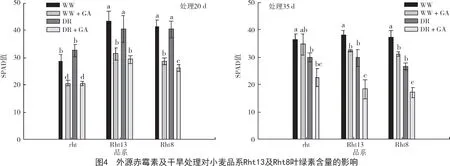
分析叶绿素含量结果发现,干旱及赤霉素处理20 d 后,在相同处理条件下rht 品系叶片叶绿素含量显著低于Rht13 和Rht8 品系(P<0.05)。而赤霉素处理35 d 后,相同水分条件下,rht 品系叶绿素含量略高于Rht13 和Rht8 品系;非赤霉素处理组,其相同水分条件下叶绿素含量几乎一致(图4)。
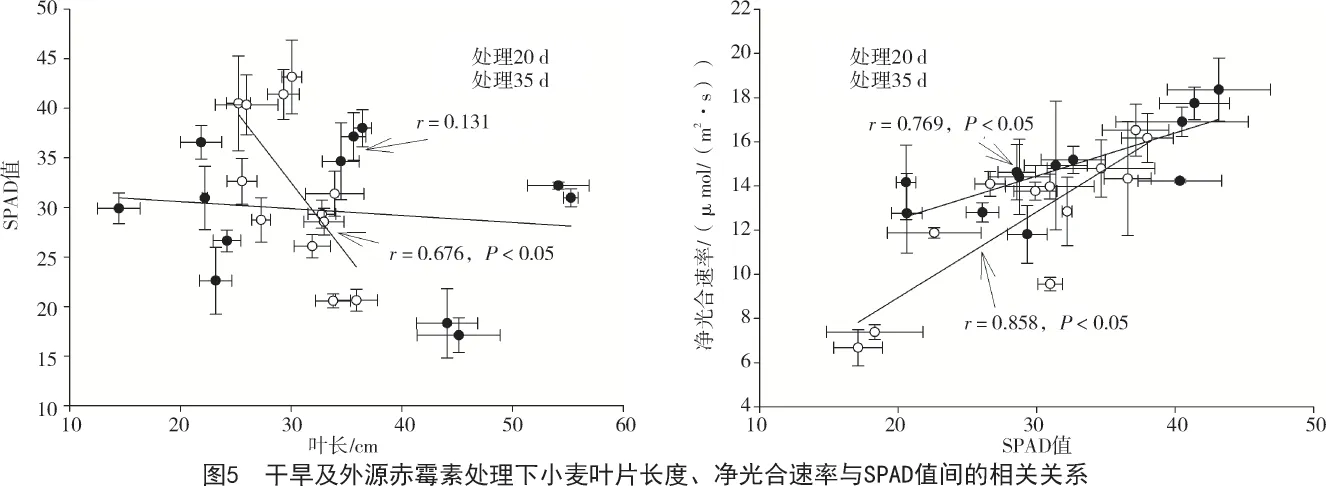
相关性分析显示,小麦叶片净光合速率与小麦叶片叶绿素含量间呈现出显著的正相关关系(图5),处理20 d 时相关系数为0.769(P<0.05);第35天时,r=0.858(P<0.05)。而小麦叶片叶绿素含量也与小麦的叶片长度在处理35 d 时呈现出显著的负相关关系(r=0.676,P<0.05)。叶绿素含量与净光合速率间呈现正相关关系早已得到证明[15-17]。说明赤霉素处理下Rht13 和Rht8 品系过分生长的叶片降低了叶片叶绿素含量,而低的叶绿素含量直接导致了净光合速率的降低。另一方面,净光合速率的降低继而导致生物量的降低。并且这种过分生长的叶片(Rht13 和Rht8 叶长分别为54.125,55.250 cm)导致小麦有向杂草化发育的倾向,最终不能正常拔节。而之前的研究发现,在逆境胁迫下,植物降低体内赤霉素含量、降低生长速率从而获得更强的抵抗胁迫的能力[14]。而对水稻赤霉素合成突变体研究发现,赤霉素合成缺陷提高了水稻对干旱的抗性[18]。在拟南芥中过表达GA20ox 带来的赤霉素含量积累同样降低了其对干旱的耐受性,而突变掉ga3ox 中的2 个基因和ga20ox 中的2 个基因则提高了其对干旱的抗性[19-20]。这与本研究的结果相似,在干旱的条件下,外源施加赤霉素将不利于小麦对干旱抗性的提高。
3 结论
在外源赤霉素处理下,叶片过分生长导致了Rht13 和Rht8 品系小麦叶片叶绿素含量的剧烈下降,进而导致了其净光合速率的降低。本研究发现,过高的赤霉素含量对Rht13 及Rht8 品素小麦的不利程度远大于其对rht 品系的负面效应。因此推测,可能Rht13 和Rht8 基因使小麦在某一种或几种活性赤霉素合成途径上存在缺陷,从而导致其对赤霉素处于长期的“饥饿”状态,从而使其对外源赤霉素更加敏感、反应更为剧烈;同时由于合成的缺陷使得赤霉素含量降低,从而使其具有更高的抗旱性。
猜你喜欢
农业工程技术(2021年12期)2021-12-23
江西农业学报(2021年2期)2021-12-08
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
农业与技术(2020年12期)2020-07-04
云南农业科技(2019年2期)2019-04-04
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
浙江农业科学(2015年2期)2015-11-24
