
喀斯特高原峡谷优势种叶片功能性状分析
2019-09-10 07:22程雯喻阳华熊康宁张俞许敏谭代军
广西植物 2019年8期
关键词:土壤养分
程雯 喻阳华 熊康宁 张俞 许敏 谭代军
摘要: 該研究对喀斯特石漠化高原峡谷地区优势种的叶片功能性状进行了分析,调查了17种植物的叶片厚度、叶面积、叶片鲜重、叶片干重、叶干物质含量、比叶面积、叶组织密度等能反映植物生存策略且易于测量的叶片功能性状,并采用逐步回归的方法探究了叶片功能性状与土壤养分之间的关系。结果表明:(1)不同优势种叶片功能性状差异明显,叶片的厚度为0.18~0.78 mm、鲜重为0.07~6.51 g、干重为0.04~3.19 g、叶面积为3.07~325.64 cm2、叶干物质含量为318.61~573.22 mg·g-1、比叶面积为60.98~236.90 cm2·g-1、叶组织密度为0.022 1~ 0.036 g·cm-3。(2)植物通过较小的比叶面积与较大的叶干物质含量来减少水分散失、增加养分储存,以适应高温、缺水少土的环境。(3)叶片功能性状之间存在广泛的相关关系,且均达到极显著水平。叶片厚度对鲜重、干重均表现为促进效应,比叶面积与叶干物质含量及叶组织密度之间均存在极显著的抑制效应。(4)叶片功能性状随土壤养分的变化产生规律性变化。土壤全氮、全钾和有机碳对叶片功能性状产生显著影响,尤以土壤有机碳的影响更为显著。进一步分析发现,喀斯特高原峡谷地区植被恢复及重建的优势种或建群种为翅荚香槐、清香木、枇杷等;施用有机肥能够提高花椒、金银花等经济林树种的适应能力。
关键词: 优势种, 叶片功能性状, 土壤养分, 喀斯特高原峡谷
中图分类号: Q948, S727/728文献标识码: A文章编号: 1000-3142(2019)08-1039-11
Abstract: Illuminating the leaf functional traits of different dominant species and discussing the correlation between leaf functional traits and rhizospheric soil nutrients in the karst plateau-canyon areas, that contribute to reveal the dominant plants’ survival strategies in specific region. In the study reported here analyzed the leaf functional traits of dominant species in the karst plateau-canyon, the following leaf functional traits were measured at each sample, leaf thickness, leaf area, leaf fresh weight, leaf dry weight, leaf dry matter content, specific leaf area, leaf tissue density. These indices were chosen because they were adequately reflected plant survival strategies and easily measurable, then explored the relationship between leaf functional traits and 4 soil nutrients by stepwise regression. The results were as follows: (1) In this area, the leaf functional traits of different dominant species were obviously different. Leaf thickness was 0.18-0.78 mm, leaf fresh weight was 0.07-6.51 g, leaf dry weight was 0.04-3.19 g, leaf area was 3.07-325.64 cm2, leaf dry matter content was 318.61-573.22 mg·g-1, specific leaf area was 60.98-236.90 cm2·g-1, leaf tissue density was 0.022 1-0.036 g·cm-3. (2) Compared to other areas, the dominant species in this area had smaller specific leaf area and bigger leaf dry matter content, which can reduce water loss and increase nutrient storage, that to adapt to high temperature and lacking of water and soil. (3) Almost all correlations significantly differed among the leaf functional traits. Leaf thickness had a promotional effect on fresh and dry weight, and specific leaf area had a significant inhibiting effect on leaf dry matter content and leaf tissue density. (4) The leaf functional traits changed regularly with the change of soil nutrients. Soil total nitrogen, total kalium and organic carbon had significant effects on leaf functional traits, especially there was very significant in organic carbon. Further analyses indicated that the most appropriate dominant species or constructive species for vegetation restoration and reconstruction in karst plateau-canyon was selected, they were Cladrastis platycarpa, Pistacia weinmannifolia, Eriobotrya japonica. In addition, applying organic fertilizer to Zanthoxylum bungeanum and Lonicera japonica, etc. could improve their adaptability. The research results above have deepened the understanding of vegetation in karst plateau-canyon areas, which provided theoretical and practical bases for vegetation restoration and reconstruction.
Key words: dominant species,leaf functional traits,soil nutrient,karst plateau-canyon area
植物功能性状(functional traits)一直是植物学与生态学研究的热点。在与环境的相互作用中,植物会通过形成多种内在生理和外在形态方面的对策以适应环境对其产生的不利影响,同时改变其对生态系统功能的贡献(孟婷婷等,2007)。植物这种能够响应生存环境的变化并对生态系统功能有一定影响的植物性状统称为植物功能性状(Diaz et al.,2001),其中与叶片相关的性状称为叶片功能性状。叶片作为与环境接触面积最大、对环境刺激最敏感的器官之一,其功能性状与植物对资源的利用能力及效率紧密相关(Sack et al.,2013)。对植物叶片功能性状的研究能够深入认识植物对其储存资源的分配方式,揭示不同植物对生境的适应策略(孙梅等,2017),从而服务于生态恢复工作中关于物种筛选(杨锐等,2015)、群落配置(曹科等,2013)等相关问题。植物叶片功能性状与海拔、光照、温度、水分等环境因素紧密相关。在大尺度范围内,植物叶片大小与年均气温及年降水量之间存在显著正相关关系,与海拔之间存在显著负相关关系(Peppe et al.,2011);比叶质量(leaf mass per area,LMA)随年降水量的增加而减小,随海拔的升高而增大(Scheepens et al.,2010)。在较小尺度范围内,土壤对于植物叶片性状的影响虽然缓慢,但土壤中的水分、氮、磷、钾、有机碳等均参与植物的养分循环与生命活动,因而对其叶片功能性状会有所影响。李丹等(2016)的研究表明土壤养分对植物功能性状有显著影响,植物通过功能性状的协调或组合以适应特定的土壤环境;康勇等(2017)的研究证实了无论是个体、种内、种间还是群落尺度,叶片功能性状变化与土壤内有机碳、氮、磷含量均密切相关。目前,对植物叶片功能性状与土壤养分之间的响应关系研究较为欠缺,且尚未形成统一的观点。
中国南方喀斯特属世界三大喀斯特集中连片区域之一,且分布面积最大、发育类型最全,贵州省位于中国南方喀斯特的中心地带,发育有多种喀斯特地貌。因此,贵州省喀斯特地区的研究结果对于相似地区具有较高的借鉴和参考价值。喀斯特地区人地矛盾突出,土地呈退化趋势,表现为石漠化亟需进行预防和治理,实现生态与经济的协调发展(熊康宁等,2011),退化植物群落的恢复与优化配置是喀斯特地区石漠化综合治理的重点内容之一(熊康宁等,2016)。优势种是对特定生境适应能力较强的树种,对喀斯特高原峡谷区优势种功能性状的分析,能够揭示该区植物对高温、干旱、土壤资源匮乏的生境所采取的较优适应策略,为植被恢复的物种筛选、群落配置及森林可持续经营奠定科学基础。因此,本文选取中国南方喀斯特关岭-贞丰花江中度-强度石漠化治理示范区的17个优势种为研究对象,分析植物叶片功能性状之间及其与根区土壤养分的响应,期望探讨并回答以下科学问题:喀斯特高原峡谷区优势种的叶片功能性状的特征如何;叶片功能性状与其根区土壤之间存在怎样的响应关系。
1材料与方法
1.1 研究地区概况
本研究区(105°36′30″—105°46′30″ E,25°39′13″—25°41′00″ N)位于贵州省西南部关岭县以南、贞丰县以北的北盘江流域花江河段,海拔为370~1 473 m,属亚热带湿润季风气候,生境要素垂直分异明显,为典型干热河谷气候。冬春温暖干旱,夏秋湿热,光热资源丰富,年均温为18.4 ℃,年均极端最高温为32.4 ℃,年均极端最低温为6.6 ℃,年总积温达6 542.9 ℃。降水充沛,年均降水量达1 100 mm,但时空分布不均,加上地区本身存在“双层结构”地质特殊性,部分地区季节性旱情严重。该区出露土层主要为中、上三叠统地层,岩石厚度大,耕作层浅薄,大多为10~20 cm,碳酸盐岩在95%以上,土壤以石灰土为主,质地黏重,缺乏團粒结构。
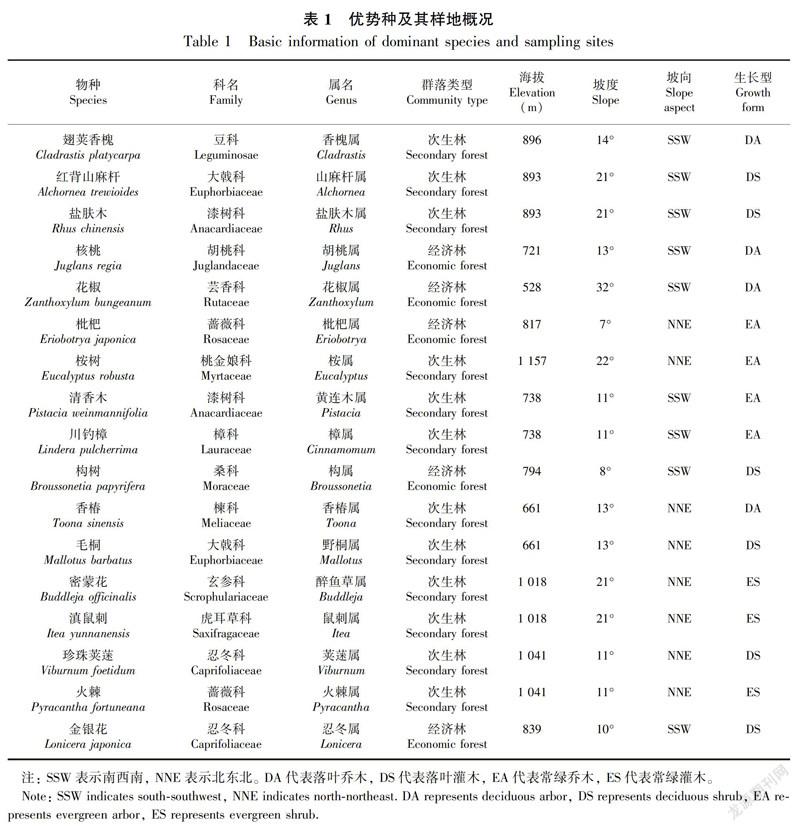
研究区虽属中-强度石漠化等级,但水热资源丰富,土壤养分含量较高(盛茂银等,2015),加之“九五”至“十三五”期间以植被恢复、水土保持等为核心的石漠化综合治理工程的实施,物种多样性显著增加。在海拔500~800 m地区已经形成了较大规模的花椒(Zanthoxylum bungeanum)、核桃(Juglans regia)特色经济林,封山育林地区已发育出以川钓樟(Lindera pulcherrima)、清香木(Pistacia weinmannifolia)为优势种的常绿林,以香椿(Toona sinensis)、毛桐(Mallotus barbatus)为优势种的落叶林。海拔800~1 000 m除较大规模构树(Broussonetia papyrifera)、枇杷(Eriobotrya japonica)、金银花(Lonicera japonica)的种植,在封山育林区也发育出了以翅荚香槐(Cladrastis platycarpa)为优势种的落叶阔叶林。在海拔1 000 m以上的封山育林地区主要有三类次生林,以密蒙花(Buddleja officinalis)、滇鼠刺(Itea yunnanensis)为优势种的常绿阔叶林;以珍珠荚蒾(Viburnum foetidum)、火棘(Pyracantha fortuneana)为优势种的常绿灌木林;以柏木(Cupressus funebris)、桉树(Eucalyptus robusta)为优势种的针阔混交林。
1.2 植被调查与样品采集
2017年7—8月,以样线法对研究区进行多次踏查,在各海拔范围(<700、700~800、800~900、>900 m,其中<700 m与>900 m的区域内群落较为单一)分别选择3~5个生长稳定且地带性较强的植物群落作为研究对象。设置20 m × 20 m的典型样地,每个林分设置3个平行,以样方法开展植物群落学调查。测定群落内乔木、灌木的名称、株数、株高、冠幅、胸径/基径,计算每个物种的重要值并进行排序,次生林优势树种筛选依据是各群落重要值排名第一或前二(第一、二相差较小)的树种,共计17种(表1)。经济林优势种为当地种植超过30 a且已形成稳定群落的树种,被认为是石漠化生态恢复的主要树种。每一样地内选取3株相互干扰较小的优势树,同步采集叶片和根区土壤。叶片的采集方法:采集各个方向外冠层、完全展开、无病虫害且未遮阴的叶片,共10片。根区土壤的采集方法:在已选择的优势种树下采集土样,采样点距离该优势树根部不超过1.0 m,且距其他树种根部不低于1.5 m。去除腐殖质层后,采用环刀法采集物理土样,因研究区土层浅薄,10 cm以下土壤砾石含量较高,因此仅取去除腐殖质层后的0~10 cm土壤作为物理土样,现场称鲜重后装入自封袋带回实验室。其次在采集物理土样处采集去除腐殖质层后0~20 cm的化学土样,均匀混合后取1.5 kg带回实验室。
1.3 指标测定与计算
1.3.1 叶片功能性状指标的选择与计算采下叶片后擦拭叶片表面灰尘,立即使用数显游标卡尺(精度0.02 mm)在叶片前、中、末端分别测量叶片厚度,测量时避开叶脉,取平均值并记录为叶片厚度(leaf thickness, LT);用电子天平(精度0.01 g)称叶片鲜重(fresh weight, FW)并记录;测定叶片鲜重时,每一叶片重复测量5次,选择较为接近的3个数值,且这3个数值的变异系数控制在5%之内,取其中位数作为测定值。如此测定10个平行,计算其平均值和标准差。此外,考虑到小叶片可能存在测量误差较大的问题。因此,本研究中涉及到叶片的干重、鲜重的测定均由同一人完成,以减少操作误差。最后采用图纸法测定叶面积:裁剪10 cm × 10 cm的滤纸,称其重量,记为M;在滤纸上画出叶片轮廓,沿轮廓剪裁并称重m,m的测定遵循上述叶片鲜重的测量标准,最终得出叶片面积(s),计算公式为s (cm2)=100 × m/M。
叶片带回实验室后,在烘箱中120 ℃杀青30 min,75 ℃烘48 h至恒重,称量并记录干重(dry weight,DW)。计算比叶面积(specific leaf area,SLA)、叶组织密度(leaf tissue density,LTD)、叶干物质含量(leaf dry matter content,LDMC)等代表性的植物叶片功能性状。相关性状计算公式:比叶面积(SLA, cm2·g-1)=叶面积/叶干重;叶组织密度(LTD,g·cm-3)=叶干重/(叶面积·叶厚度);叶干物质含量(LDMC,mg·g-1)=叶干重/叶鲜重。
1.3.2 土壤指标的测定将采集的化学土壤样品带回实验室后剔除可见的动植物残体及砾石等杂物,自然风干后研磨,分别过2 mm、0.15 mm筛备用。测定土壤容重(bulk density,BD)、全氮(total nitrogen,TN)、全磷(total phosphorus,TP)、全钾(total kalium,TK)、有机碳(organic carbon,OC)。土壤容重采用环刀法测定,土壤全氮采用半微量开氏法测定,全磷的测定采用NaOH熔融-钼锑抗比色法,全钾采用NaOH熔融-火焰光度法测定,有机碳采用重铬酸钾容量法-外加热法测定(鲍士旦,2008)。
1.4 数据处理和分析
用Microsoft Excel 2010对数据进行初步计算整理后,用OriginPro 2017作图,用SPSS22.0完成统计分析。不同优势种叶片功能性状差异用单因素方差分析(one-way ANOVA),用Spearman相关分析检验各功能性状的相关性,用逐步回归分析确定叶片功能性状与土壤养分之间的关系。
2结果与分析
2.1 不同优势种叶片功能性状变异特征
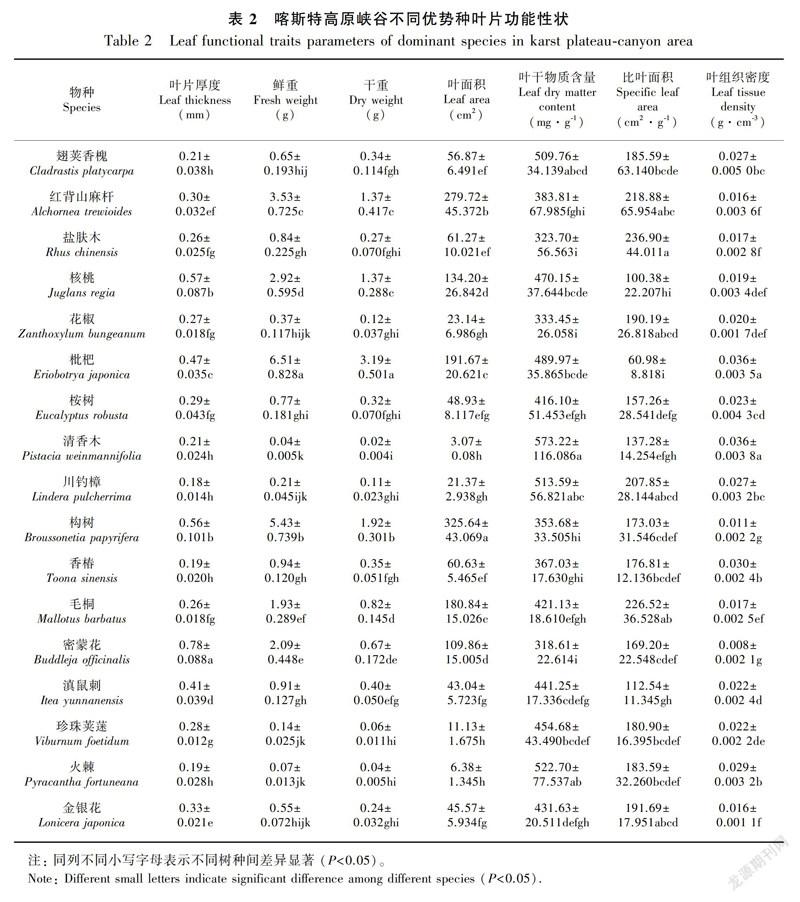
17种优势种植物的叶片性状如表2所示,不同物种各个性状之间存在一定差异。17种植物的叶片厚度变化范围为0.18~0.78 mm,总体变异系数为50.02%;叶片鲜重的变化范围为0.07~6.51 g,总体变异系数为116.07%;叶片干重变幅为0.04~3.19 g,总体变异系数为124.03%;叶面积介于3.07~325.64 cm2间,总体变异系数为101.48%;叶干物质含量最大的是清香木(573.22 mg·g-1),最小的是密蒙花(318.61 mg·g-1),总体变异系数为20.51%;比叶面积最大的是盐肤木(236.90 cm2·g-1),最小的是枇杷(60.98 cm2·g-1),总体变异系数为31.96%;叶组织密度的最大值为枇杷(0.036 g·cm-3),最小值为密蒙花(0.008 g·cm-3),总体变异系数为37.55%。由表1、表2可知,灌木叶片比叶面积略高于乔木,常绿树种叶片比叶面积略高于落叶树种,经济林树种比叶面积略低于次生林树种。此外,不同海拔植物叶片功能性状差异显著,但尚未发现其明显的变化规律。
2.2 不同地区优势种的叶片功能性状特点
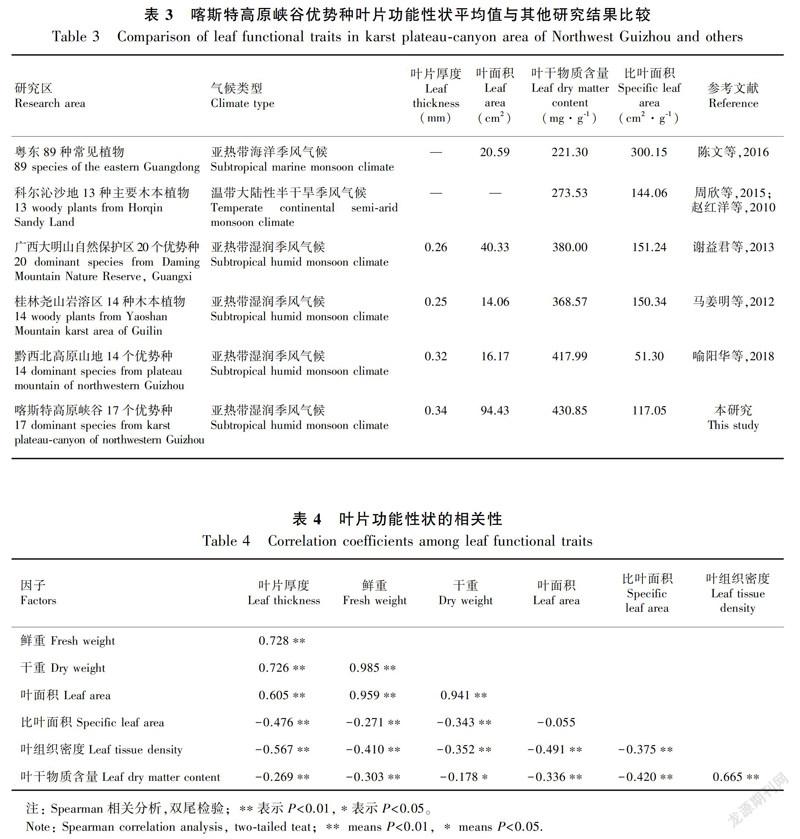
表3显示,贵州地区树种叶片厚度略大于广西地区,叶面积的排序为本研究>广西大明山>粤东>黔西北>桂林尧山,叶干物质含量的排序为本研究>黔西北>广西大明山>桂林尧山>科尔沁沙地>粤东,比叶面积的排序为粤东>广西大明山>桂林尧山>科尔沁沙地>本研究>黔西北。
2.3 叶片功能性状之间的关系
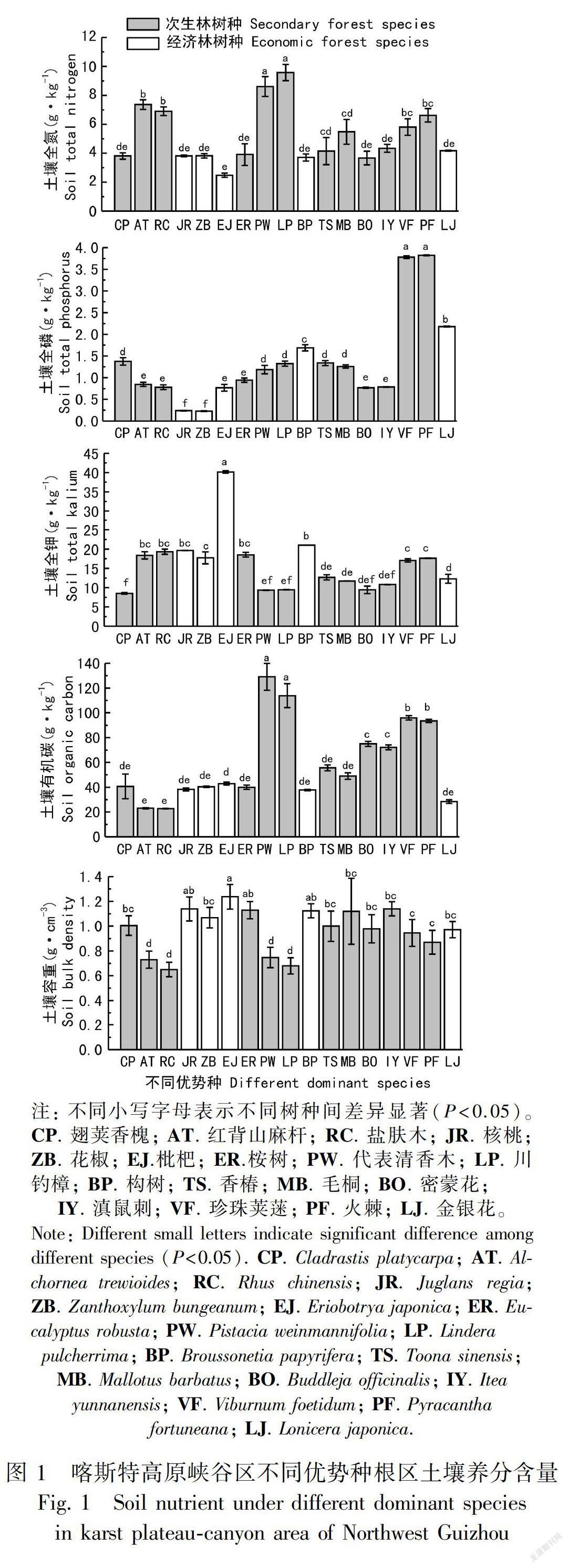
17个优势种叶片功能性状的相关性分析显示,叶片厚度与叶片鲜重、干重、叶面积呈极显著正相关(P<0.01,下同);叶片鲜重与干重、叶面积呈极显著正相关;叶片干重与叶面积呈极显著正相关;叶片厚度、鲜重、干重、叶面积均与比叶面积、叶干物质含量、叶组织密度呈极显著负相关;比叶面积与叶组织密度、叶干物质含量呈极显著负相关;叶组织密度與叶干物质含量呈极显著正相关(表4)。
2.4 叶片功能性状与土壤的关联性
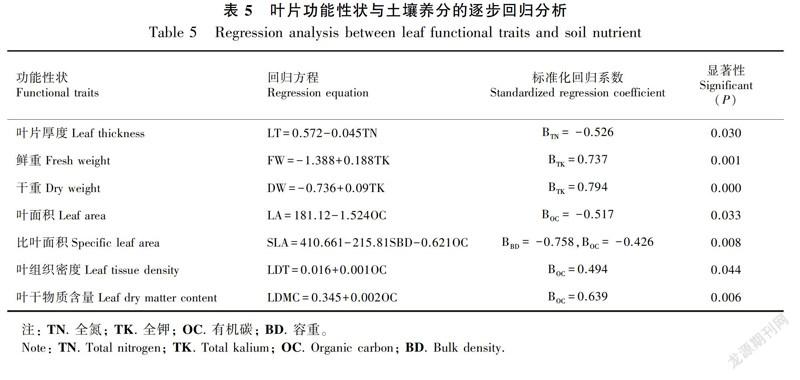
不同优势种根区土壤理化性质存在较大差异(图1)。次生林的土壤容重(红背山麻杆0.73 g·cm-3、清香木0.75 g·cm-3、珍珠荚蒾0.95 g·cm-3、火棘0.87 g·cm-3等)总体低于经济林(核桃1.14 g·cm-3、花椒1.07 g·cm-3、枇杷1.24 g·cm-3、构树1.12 g·cm-3等);次生林的比叶面积(红背山麻杆218.88 cm2·g-1、盐肤木236.90 cm2·g-1、川钓樟207.85 cm2·g-1、毛桐226.52 cm2·g-1等)总体高于经济林(核桃100.38 cm2·g-1、花椒190.19 cm2·g-1、枇杷60.89 cm2·g-1、构树173.81 cm2·g-1等); 与次生林树种根区土壤相比, 经济林根区土壤全氮、全磷、有机质含量总体更低,全钾含量更高。逐步回归分析结果显示,在喀斯特高原峡谷地区,土壤养分中尤以全氮、全钾、有机碳的含量对叶片功能性状的影响较为显著(表5)。其中,叶片厚度与土壤全氮含量显著负相关,叶片鲜重、干重与土壤全钾含量呈显著正相关,叶面积、比叶面积、叶组织密度、叶干物质含量均与土壤有机碳含量具有显著的相关性。比叶面积的回归方程中土壤容重的标准化回归系数(-0.758)绝对值大于土壤有机碳 (-0.426), 表明影响比叶面积的主要因子是土壤容重,其次是土壤有机碳含量。
3讨论与结论
3.1 不同地区优势种叶片功能性状差异
从全球尺度来看,不同气候类型植物的叶片功能性状会表现出很大的差异(Wright et al.,2004),在同一气候带中,植物叶片性状也会随其生境的具体地形、海拔等环境因子产生差异(冯秋红等,2008)。喀斯特高原峡谷地区,高温、季节性干旱及缺少土壤是当地树种需要适应的主要逆境因子,因而植物形成了相应的有别于其他地区的叶片功能性状组合, 这些性状差异能够反映植物本身固有的遗传特性以及其为适应其生境而采取的注: 不同小写字母表示不同树种间差异显著(P<0.05)。CP. 翅荚香槐; AT. 红背山麻杆; RC. 盐肤木; JR. 核桃; ZB. 花椒; EJ.枇杷; ER.桉樹; PW. 代表清香木; LP. 川钓樟; BP. 构树; TS. 香椿; MB. 毛桐; BO. 密蒙花; IY. 滇鼠刺; VF. 珍珠荚蒾; PF. 火棘; LJ. 金银花。
方案策略(刘晓娟和马克平,2015)。本研究中各个地区植物叶面积之间存在显著差异但没有明显规律,这可能因为植物遗传特性对于叶面积的影响远大于环境因素。
比叶面积是衡量植物对生境资源利用效率的重要指标(Vendramini et al.,2002)。粤东地区植物具有最大的比叶面积,本研究的比叶面积处于较低水平,仅高于黔西北高原山地。粤东地区在气候条件、土壤资源等方面均为植物提供了丰富的可利用资源,因此其比叶面积愈大,表明植物叶片捕获光能量的能力愈强,光合效率愈高,则植物具有更快的资源流动速度与回归效率(Ninemets et al.,2007),即获得更快的相对生长速率以维持其植物体争夺资源的优势,则植物对生境资源的利用效率较高。而本研究区与黔西北高原山地区石漠化发育,水土资源匮乏是植物生长的主要制约因素,更小的比叶面积表明单位干重所占面积更小,即叶片干物质的密度越大,植物光合效率越低,养分循环与回归速率越慢,加之该地区土壤养分总量较少,植物将更多的养分投入到基础生理活动,导致该区植物大多生长为“小老头树”,即说明该区植物对于生境资源的利用效率相对较低。
叶干物质含量的大小反映了植物积累养分元素的能力(祁建等,2008)。贵州两个研究区叶干物质含量更高,表明该地区植物投入叶片建成的比例更高,其耐受性越强,植物对养分的积累能力越强;粤东地区较低的叶干物质含量则表明植物用于叶片建成投入的比例较小,叶片组织密度较低,叶片内活动更强,植物吸收更多养分被用于加速植物生长,由此产生的对资源越来越强的竞争导致植物对养分的消耗变高,因此植物对养分的积累能力较低。对比结果中科尔沁沙地植物的叶干物质含量仅高于粤东,而低于黔桂地区,可能因为科尔沁沙地虽存在季节性缺水,但其土层厚且松散,对于木本植物根系深入土层吸水有利,吸收的营养物质较大程度地投入到其根系发展(王凯等,2014),因此用于叶片建成部分较少;而黔桂地区土层较薄,尤其是黔西北和本研究所选的喀斯特高原峡谷地区,植物根系的生长受到阻碍, 因此大部分植物将吸收的养分更多地用于植物叶片的构建,增大地表生物量,加快生长速率,从而储存更多的营养物质以适应贫瘠的土壤环境。
3.2 喀斯特高原峡谷优势种叶片功能性状之间的协调关系
叶片功能性状之间存在多种关联性,一系列叶片功能性状组合同时发生作用,以实现植物对环境的适应(施宇等,2011),这反映了植物对环境适应策略的趋同性。喀斯特高原峡谷土壤及水分资源匮乏,植物为适应该环境采取了较为明显的功能平衡策略。其中,比叶面积与叶干物质含量、叶组织密度均呈极显著负相关关系,这与杨士梭等(2014)的研究结论一致。较大的叶组织密度和叶干物质含量反映了植物合成的干物质更多投入到叶片的构建,叶肉密度增大、叶片含水量减少,即增加了叶片内部水分向叶片表面扩散的距离或阻力(王瑞丽等,2016),相应的比叶面积则处于较小的水平,从而降低了水分散失,获得更强的对于干旱环境的抗逆性。此外,比叶面积是叶面积与叶干重的比值,但除比叶面积与叶面积关系不密切,其余指标之间均存在显著或极显著的相关性,其原因可能是研究区太阳辐射强度大、大气温度高、叶片温度变化明显,而不同树种由于层片、捕光性能、方位和叶夹角等不一样,所以叶片温度的变化趋势不一致,因此叶片含水量波动较大。
3.3 喀斯特高原峡谷优势种叶片功能性状对土壤因子的响应
植物叶片功能性状与其生境土壤的养分资源存在一定的响应关系,土壤的肥力情况决定了植物对资源的利用策略(刘旻霞和马建祖,2012),且土壤中氮、磷、有机碳含量的变化对植物功能性状有显著的影响(白雪等,2014)。在喀斯特高原峡谷地区,土壤容重及土壤养分中的全氮、全钾、有机碳的含量均对植物叶片功能性状具有重要影响,对植物适应环境的策略具有强烈的调控作用。
喀斯特高原峡谷地区,高温干旱是其主要的环境特点之一,植物的光合能力会因由叶片防御系统控制的气孔关闭而明显受阻,而钾可以通过促进叶绿素的合成和类囊体膜的稳定(Terry et al.,1983),增强气孔的调节能力,从而促进植物的光合作用,积累养分,获得较高的生物量,因而在一定范围内,土壤全钾含量越高,植物获得更大的生物量,则叶片鲜重与干重越大。
在喀斯特高原峡谷地区,植物叶面积与比叶面积随土壤有机碳含量的增加而减小,而叶干物质含量与叶组织密度的变化规律则相反。通常,喀斯特地区次生林的水土流失导致土层浅薄;同时,裸露岩石土壤养分聚集效应使其养分含量较高,此外人为干扰较少,土壤动物及微生物活动较为剧烈,枯落物分解彻底,土壤有机碳含量相对较高。本研究所选的次生林优势种如清香木、滇鼠刺、火棘、珍珠荚蒾等,为适应缺水、少土的条件,减少叶片水分蒸发,形成了较小的叶面积和比叶面积;但土壤养分条件较好,植物有较多的可利用养分用于合成叶片干物质,从而较大程度保存养分以适应恶劣的环境条件,因此叶组织密度与叶干物质含量处于较高水平。
3.4 基于叶片功能性状的恢复树种选择与经营
较大的叶干物质含量与较小的比叶面积是喀斯特高原峡谷地区植物适应环境的主要策略。据此筛选出对干旱、高温环境适应性较强的树种包括翅荚香槐、清香木、火棘、核桃、枇杷等,可用作该地区植被恢复和优化配置的优势种或建群种。
叶片功能性状与土壤养分的逐步分析结果显示,研究区土壤有机碳显著影响叶片比叶面积及叶干物质含量,随土壤有机碳含量的增加,比叶面积减小、叶干物质含量增高。由此可推测经济林中花椒、构树、金银花等树种的叶干物质含量较低、比叶面积较大的原因与有机碳含量较低有关,次生林树种适应性较强与其有机碳含量丰富和周转快速相关。因而,适量施用有机肥能够提高经济树种对高温、缺水少土环境的适应能力,以达到更高的生态经济效益。对天然次生林,培育针阔混交林,增加凋落物数量,加速有机质的分解和循环,是提高森林生态系统适应性的重要途径。
参考文献:
BAI X, CHENG JH, ZHENG SX, et al., 2014. Ecophysiological responses of Leymus chinensis to nitrogen and phosphorus additions in a typical steppe [J]. Chin J Plant Ecol, 38(2): 103-115. [白雪, 程军回, 郑淑霞, 等, 2014. 典型草原建群种羊草对氮磷添加的生理生态响应 [J]. 植物生态学报, 38(2): 103-115.]
BAO SD, 2008. Soil agro-chemical analysis. [M]. 3rd ed. Beijing: China Agriculture Press: 25-144. [鲍士旦, 2008. 土壤农化分析 [M]. 3版. 北京: 中国农业出版社: 25-114.]
CAO K, RAO MD, YU JZ, et al., 2013. The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest [J]. Biodivers Sci, 21(5): 564-571. [曹科, 饶米德, 余建中, 等, 2013. 古田山木本植物功能性状的系统发育信号及其对群落结构的影响 [J]. 生物多样性, 21(5): 564-571.]
CHEN W, WANG JH, MA RJ, et al., 2016. Variance in leaf functional traits of 89 species from the eastern Guangdong of China [J]. Chin J Ecol, 35(8): 2101-2109. [陳文, 王桔红, 马瑞君, 等, 2016. 粤东89种常见植物叶功能性状变异特征 [J]. 生态学杂志, 35(8): 2101-2109.]
DIAZ S, CABIDO M, 2001. Vive la difference: Plant functional diversity matters to ecosystem processes [J]. Trend Ecol Evol, 16(11): 646-655.
FENG QH, SHI ZM, DONG LL, 2008. Response of plant functional traits to environment and its application [J]. Sci Silv Sin, 44(4): 125-131. [冯秋红, 史作民, 董莉莉, 2008. 植物功能性状对环境的响应及其应用 [J]. 林业科学, 44(4): 125-131.]
KANG Y, XIONG MX, HUANG J, et al., 2017. Variation in woody plant functional traits of the tropical cloud forests in Bawangling, Hainan Island [J]. Acta Ecol Sin, 37(5): 1572-1582. [康勇, 熊梦辉, 黄瑾, 等, 2017. 海南岛霸王岭热带云雾林木本植物功能性状的分异规律 [J]. 生态学报, 37(5):1572-1582.]
LI D, KANG SRL, ZHAO MY, et al., 2016. Relationships between soil nutrients and plant functional traits in different degradation stages of Leymus chinensis steppe in Nei Mongol, China [J]. Chin J Plant Ecol, 40(10): 991-1002. [李丹, 康萨如拉, 赵梦颖, 等, 2016. 内蒙古羊草草原不同退化阶段土壤养分与植物功能性状的关系 [J]. 植物生态学报, 40(10): 991-1002.]
LIU MX, MA JZ, 2012. Responses of plant functional traits and soil factors to slope aspect in alpine meadow of South Gansu, Northwest China [J]. Chin J Appl Ecol, 23(12): 3295-3300. [刘旻霞, 马建祖, 2012. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应 [J]. 应用生态学报, 23(12):3295-3300.]
LIU XJ, MA KP, 2015. Plant functional traits-concepts, applications and future directions [J]. Sci Sin (Vitae), 45(4): 325-339. [刘晓娟, 马克平, 2015. 植物功能性状研究进展 [J]. 中国科学: 生命科学, 45(4): 325-339.]
MA JM, ZHANG XZ, LIANG SC, et al., 2012. Leaf traits of common plants in Yaoshan Mountain of Guilin, China [J]. J Guangxi Norm Univ (Nat Sci Ed), 30(1): 77-82. [马姜明, 张秀珍, 梁士楚, 等, 2012. 桂林尧山常见植物叶片性状研究 [J]. 广西师范大学学报(自然科学版), 30(1): 77-82.]
MENG TT, NI J, WANG GH, 2007. Plant functional traits, environments and ecosystem functioning [J]. Chin J Plant Ecol, 31(1): 150-165. [孟婷婷, 倪健, 王国宏, 2007. 植物功能性状与环境和生态系统功能 [J]. 植物生态学报, 31(1): 150-165.]
NIINEMETS , 2007. Photosynthesis and resource distribution through plant canopies [J]. Plant Cell & Environ, 30(9): 1052.
PEPPE DJ, ROYER DL, CARIGLI NO, et al., 2011. Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications [J]. New Phytol, 190(3): 724-739.
QI J, MA KM, ZHANG YX, 2008. Comparisons on leaf traits of Quercus liaotungensis on different slope positions in Dongling Mountain of Beijing [J]. Acta Ecol Sin, 28(1): 122-128. [祁建, 馬克明, 张育新, 2008. 北京东灵山不同坡位辽东栎(Quercus liaotungensis)叶属性的比较 [J]. 生态学报, 28(1): 122-128.]
SACK L, SCOFFONI C, JOHN G P, et al., 2013. How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis [J]. J Exp Bot, 64(13): 4053-4080.
SCHEEPENS JF, FREI ES, STOCKLIN J, 2010. Genotypic and environmental variation in specific leaf area in a widespread alpine plant after transplantation to different altitudes [J]. Oecologia, 164(1): 141-150.
SHENG MY, XIONG KN, CUI GY, et al., 2015. Plant diversity and soil physical-chemical properties in karst rocky desertification ecosystem of Guizhou, China [J]. Acta Ecol Sin, 35(2): 434-448. [盛茂银, 熊康宁, 崔高仰, 等, 2015. 贵州喀斯特石漠化地区植物多样性与土壤理化性质 [J]. 生态学报, 35(2): 434-448.]
SHI Y, WEN ZM, GONG SH, 2011. Comparisons of relationships between leaf and fine root traits in hilly area of the Loess Plateau, Yanhe River basin, Shaanxi Province, China [J]. Acta Ecol Sin , 31(22): 6805-6814. [施宇, 温仲明, 龚时慧, 2011. 黄土丘陵区植物叶片与细根功能性状关系及其变化 [J]. 生态学报, 31(22): 6805-6814.]
SUN M, TIAN K, ZHANG Y, et al., 2017. Research on leaf functional traits and their environmental adaptation [J]. Plant Sci J, 35(6): 940-949. [孙梅, 田昆, 张贇, 等, 2017. 植物叶片功能性状及其环境适应研究 [J]. 植物科学学报, 35(6): 940-949.]
TERRY N, 1983. Limiting factors in photosynthesis: IV. Iron stress-mediated changes in light-harvesting and electron transport capacity and its effects on photosynthesis in vivo [J]. Plant Physiol, 71(4): 855-860.
VENDRAMINI F, DIAZ S, GURVICH DE, et al., 2002. Leaf traits as indicators of resource-use strategy in floras with succulent species [J]. New Phytol, 154(1): 147-157.
WANG K, SONG LN, L LY, et al., 2014. Fine root adaptive strategy of main afforestation tree species in Horqin sandy land [J]. J Arid Land Resour Environ, 28(12): 128-131. [王凯, 宋立宁, 吕林有, 等, 2014. 科尔沁沙地主要造林树种细根适应策略 [J]. 干旱区资源与环境, 28(12): 128-131.]
WANG RL, YU GR, HE NP, et al., 2016. Altitudinal variation in the covariation of stomatal traits with leaf functional traits in Changbai Mountain [J]. Acta Ecol Sin , 36(8): 2175-2184. [王瑞丽, 于贵瑞, 何念鹏, 等, 2016. 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律—以长白山为例 [J]. 生态学报, 36(8): 2175-2184.]
WRIGHT IJ, REICH PB, WESTOBY M, et al., 2004. The worldwide leaf economics spectrum [J]. Nature, 428(6985): 821.
XIE YJ, LU XM, LAN JC, et al., 2013. Functional traits of evergreen broad-leaved forest in Daming Mountain Nature Reserve, Guangxi [J] J S Agric, 44(9): 1504-1510. [谢益君, 陆晓明, 蓝嘉川, 等, 2013. 广西大明山常绿阔叶林功能群间的功能性状关系 [J]. 南方农业学报, 44(9): 1504-1510.]
XIONG KN, CHEN YB, CHEN H, et al., 2011. Technology and model of rocky desertification control in Guizhou [M]. Guiyang: Guizhou Science and Technology Publishing House: 12-22. [熊康宁, 陈永毕, 陈浒, 等, 2011. 点石成金: 贵州石漠化治理技术与模式 [M]. 贵阳: 贵州科技出版社: 12-22.]
XIONG KN, ZHU DY, PENG T, et al., 2016. Study on ecological industry technology and demonstration for karst rocky desertification control of the karst plateau-gorge [J]. Acta Ecol Sin, 36(22): 7109-7113. [熊康宁, 朱大运, 彭韬, 等, 2016. 喀斯特高原石漠化综合治理生态产业技术与示范研究 [J]. 生态学报, 36(22): 7109-7113.]
YANG R, ZHANG BR, WANG LL, et al., 2015. The response of plant functional traits group to gradients of altitude in dry-hot valley of Yuan-Mou [J]. Ecol Environ Sci, 24(1): 49-56. [杨锐, 张博睿, 王玲玲, 等, 2015. 元谋干热河谷植物功能性状组合的海拔梯度响应 [J]. 生态环境学报, 24(1): 49-56.]
YANG SS, WEN ZM, MIAO LP, et al., 2014. Responses of plant functional traits to micro-topographical changes in hilly and gully region of the Loess Plateau, China [J]. Chin J Appl Ecol, 25(12): 3413-3419. [楊士梭, 温仲明, 苗连朋, 等, 2014. 黄土丘陵区植物功能性状对微地形变化的响应 [J]. 应用生态学报, 25(12): 3413-3419.]
YU YH, ZHONG XP, CHENG W, 2018. Analysis of relationship among leaf functional traits and economics spectrum of dominant species in northwestern Guizhou Province [J]. J For Environ, 38(2): 196-201. [喻阳华, 钟欣平, 程雯, 2018. 黔西北地区优势树种叶片功能性状与经济谱分析 [J]. 森林与环境学报, 38(2): 196-201.]
ZHAO HY, LI YL, WANG XY, et al., 2010. Variations in leaf traits of 52 plants in Horqin sandy land [J]. J Desert Res, 30(6): 1292-1298. [赵红洋, 李玉霖, 王新源, 等, 2010. 科尔沁沙地52种植物叶片性状变异特征研究 [J]. 中国沙漠, 30(6): 1292-1298.
ZHOU X, ZUO XA, ZHAO XY, et al., 2015. Plant functional traits and interrelationships of 34 plant species in South Central Horqin sandy land, China [J]. J Desert Res, 35(6): 1489-1495. [周欣, 左小安, 赵学勇, 等, 2015. 科尔沁沙地中南部34种植物叶功能性状及其相互关系 [J]. 中国沙漠, 35(6): 1489-1495.]
猜你喜欢
农业与技术(2016年22期)2017-03-07
农业与技术(2016年21期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
农民致富之友(2016年22期)2017-02-07
河北渔业(2015年5期)2016-03-01
天津农业科学(2015年12期)2015-12-03
湖北农业科学(2015年16期)2015-10-28
安徽农学通报(2015年15期)2015-08-19
湖北农业科学(2014年11期)2014-09-10
