
水稻穗分化期外源植物生长调节剂处理对颖花分化与退化的影响
2019-09-10 07:22文廷刚王伟中杨文飞顾大路钱新民施洪泉孙爱侠吴传万
江苏农业学报 2019年3期
文廷刚 王伟中 杨文飞 顾大路 钱新民 施洪泉 孙爱侠 吴传万







摘要:以南粳9108为材料,在水稻生长的第一苞分化期(I期)、枝梗及颖花分化期(Ⅱ期)、颖花分化一雌雄蕊形成期(Ⅲ期)、雌雄蕊形成一减数分裂期(Ⅳ期)喷施乙烯利( Eth,lOxl0mol/L)和多效唑(PP333,300 mg/L),以清水为对照(CK),研究不同时期外源调节剂处理对稻穗一次枝梗颖花和二次枝梗颖花的分化、退化以及穗部性状和产量构成的影响。结果表明,乙烯利在幼穗分化各时期处理均显著增加了水稻总颖花分化数和退化数,但每穗颖花数均表现为降低,尤以Ⅳ期每穗颖花数降低最多,其中主要降低了二次枝梗颖花数。多效唑在幼穗分化前期(I和Ⅱ期)处理总颖花分化数、总颖花退化数和每穗颖花数均表现为增加;而在后期(Ⅲ和Ⅳ期)处理则相反。相关性分析结果显示,水稻结实率和实粒数与二次枝梗颖花数关系更紧密,而总颖花数和千粒质量与一次枝梗颖花数关系更紧密。方差分析结果显示,不同时期处理和调节剂处理以及不同时期处理与调节剂处理的交互作用对水稻穗部性状的影响均达到显著水平。因此,乙烯利处理通过减少一次枝梗颖花数和二次枝梗颖花数,从而降低了每穗颖花数和实粒数,空秕粒数增加,导致了结实率、千粒质量和产量的下降。多效唑前期(I和Ⅱ期)处理通过提升一次枝梗颖花数和二次枝梗颖花数,促进了每穗颖花数的增加,从而提高了实粒数、结实率和千粒质量,最终提高了产量;但在后期(Ⅲ和Ⅳ期)处理则相反。
关键词:水稻;植物生长调节剂;穗分化期;颖花分化与退化;结实率
中图分类号:S511
文献标识码:A
文章编号: 1000-4440( 2019) 03-0514-09
每穗粒数是水稻产量的主要构成因素之一,其形成与水稻颖花分化、退化、发育和结实等生理过程密切相关[1]。水稻穗分化一般持续30 d左右,可分为苞分化期、枝梗分化期、颖花分化期、花粉母细胞形成期、减数分裂期和花粉充实完成期等5个时期,每个时期均有各自的特点[2]。其中,枝梗和颖花分化期是决定每穗粒数起始数的关键时期,减数分裂期是决定颖花退化数的关键时期,在这2个时期共同作用下决定了最后的穗粒数,而此时期也是穗分化对外界环境最敏感的时期。前人研究结果表明,温度、氮肥、CO2浓度以及调节剂等因素在水稻幼穗分化期均会显著影响水稻颖花的分化和退化,进而影响穗粒数的形成和产量的提升[3-6]。
植物激素與水稻穗部性状发育密切相关。研究结果显示,植物激素对作物幼穗分化及籽粒发育、灌浆等生理过程起到重要的调控作用[7-8]。黄静静等[9]研究发现,在水稻穗分化期喷施6-BA能显著提高每穗枝梗和颖花数,促进水稻穗长发育。顾大路等[1O]研究发现乙烯利可以延迟水稻的开花,且不影响水稻结实率。宫彦龙等研究指出,幼穗分化期喷施油菜素内酯可增加二次枝梗结实率,提高穗粒数[11];而在水稻幼穗分化的苞分化期、枝梗分化期、颖花分化期、花粉母细胞形成及减数分裂期和花粉充实完成期喷施细胞分裂素后,水稻穗长、二次枝梗结实率和穗结实率均显著高于对照[12],乙烯利是一种植物生长调节剂,也是乙烯释放剂[13],乙烯被认为是一种重要的气体植物激素,广泛存在于植物的各个组织中,并参与调节植物生长、发育以及响应外界环境等一系列生物学过程,如种子萌发、细胞的分裂与伸长、幼苗及成株形态建成、叶片衰老、性别分化、果实成熟及抵抗生物和非生物胁迫等[14]。多效唑是一种高效低毒的植物生长延缓剂,具有调节植物生长、抑制茎秆伸长、促进分蘖、提高产量和增强抗逆性等多种作用[15]。研究结果显示,多效唑在水稻幼穗分化期间使用可影响幼穗分化、延缓抽穗,常在杂交稻制种中调控水稻花期[16-17]。虽然前人对水稻颖花的分化和退化的研究报道较多,但是均主要集中在水稻品种[18]、肥料[1]、高温[3]等因素对颖花的影响,而研究乙烯利和多效唑在不同时期施用对水稻颖花分化与退化调控的报道较少。本研究以南粳9108为试验材料,在幼穗分化不同时期喷施乙烯利和多效唑处理,研究颖花性状对乙烯利和多效唑的调控响应,以期为调控水稻穗部发育以及穗粒数的形成,指导水稻生产提供依据。
1 材料与方法
1.1 材料
供试水稻品种为南粳9108。2种植物生长调节剂分别为15%的多效唑(PP333)可湿性粉剂和40%乙烯利(Eth)水剂。
1.2 试验设计
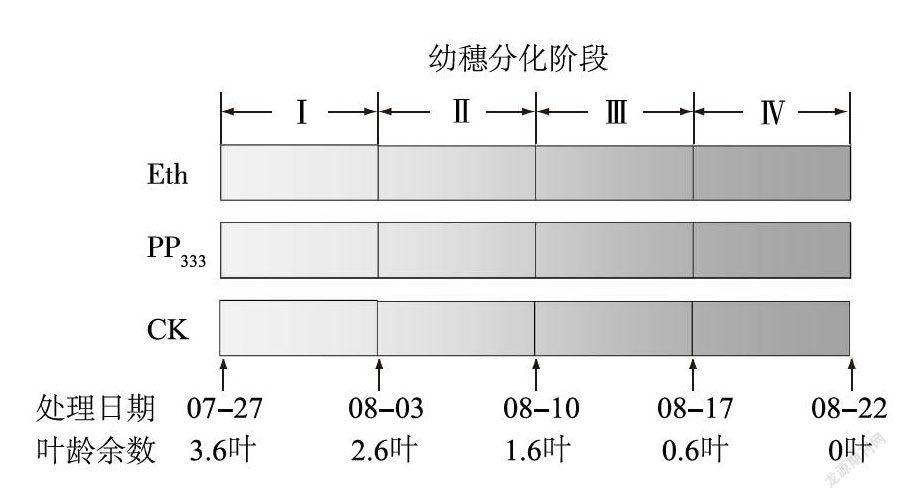
试验于2017年在江苏徐淮地区淮阴农业科学研究所现代农业高新科技园区内进行。南粳9108于5月2日播种,大田秧盘育秧,6月12日人工移栽,株距15 cm,行距20 cm,小区面积为20m-。试验采用随机区组设计,设4个处理时期,3个调节剂处理,3次重复。处理时期参考水稻叶龄模式[2]将水稻幼穗分化期细分为:第一苞分化期(I)、枝梗一颖花分化期(Ⅱ)、颖花分化一雌雄蕊形成期(Ⅲ)及雌雄蕊形成一减数分裂期(Ⅳ),具体处理日期见图1。外源调节剂处理分别为:乙烯利(0.01 mol/L)、多效唑(300 mg/L),以喷清水为对照( CK)。调节剂使用喷雾筒喷施,用药量为50ml/m2,喷雾时以植株上部叶片湿润又不滴水为标准。秧苗移栽前,大田基施尿素150 kg/hm2,过磷酸钙360 kg/hm2,氯化钾110 kg/hm2,移栽后追施尿素109 kg/hm2作为分蘖肥,其余管理措施同大田常规栽培管理。
1.3 测定项目与方法
1.3.1 颖花分化与退化调查水稻稻穗全抽出时,每处理取长势一致的水稻10穴,选取其中穗子大小基本相同的主茎穗15个,考察每穗一次枝梗上的颖花数和退化数、二次枝梗上的颖花数和退化数,并计算每穗颖花数、每穗颖花退化数及一次枝梗、二次枝梗上的颖花分化数。计算公式如下:
颖花数=一次枝梗上的颖花数+二次枝梗上的颖花数
颖花退化数=一次枝梗上的颖花退化数+二次枝梗上的颖花退化数
一次枝梗颖花分化数=一次枝梗上的颖花数+一次枝梗上的颖花退化数
二次枝梗颖花分化数=二次枝梗上的颖花数+二次枝梗上的颖花退化数
1.3.2 穗部性状水稻成熟后,每处理选取长势一致的水稻10穴,取其中15个穗子大小基本相同的主茎穗,考察水稻每穗一次枝梗数、二次枝梗数、穗粒数、饱粒数、秕粒数、结实率和千粒质量。
1.3.3 数据处理利用Excel 2016和SPSS 18.0软件对数据进行统计分析,用最小显著差数法( LSD)进行差异显著性检验,并用Sigmaplot 14.0软件作图。
2 结果与分析
2.1 不同时期调节剂处理水稻穗部主要性状间的差异
如表l所示,不同处理组合间在水稻穗部主要性状,包括总颖花分化数、每穗颖花数、总颖花退化数、一二次枝梗分化数以及实粒数、结实率等上均存在极显著影响。不同时期主效应除对一次枝梗颖花分化数比例和二次枝梗颖花分化数比例无显著影响外,在其余穗部性状上均存在极显著影响;而调理剂处理主效应对所有主要穗部性状均有极显著影响,且高于不同时期主效应。二者交互作用除在二次枝梗颖花分化数比例上无显著影响外,对其余穗部性状也存在极显著的影响。
2.2 不同时期调节剂处理对水稻每穗颖花数的影响
由图2可见,乙烯利和多效唑处理的每穗颖花数随着处理时间的推移总体呈下降趋势。与对照相比,乙烯利在穗分化I、Ⅱ、Ⅲ和Ⅳ期处理均显著降低了每穗颖花数,分别降低了2.77%、4.00%、10.24%和11.47%。多效唑处理在I和Ⅱ期处理均能显著增加每穗颖花数,尤以Ⅱ期增幅最大;而Ⅳ期处理则显著降低了每穗颖花数。可见,在幼穗分化期的不同时期使用乙烯利对每穗颖花的形成均有抑制作用,特别是在颖花分化一雌雄蕊形成期(Ⅲ期)至雌雄蕊形成一减数分裂期(Ⅳ期)抑制作用最显著。与乙烯利不同,多效唑在枝梗及颖花分化期(Ⅱ期)使用能显著增加每穗颖花数,而在雌雄蕊形成一减数分裂期(Ⅳ期)使用则出现显著抑制作用。
2.3 不同时期调节剂处理对水稻每穗颖花分布的影响
对不同时期调节剂处理后的每穗颖花进行观察(表2)可知,两调节剂处理的一次枝梗颖花数和二次枝梗颖花数的变化趋势与每穗颖花数基本一致。与对照相比,在穗分化各时期乙烯利处理均显著降低了一次枝梗颖花数和二次枝梗颖花数,其中以Ⅳ期处理降低最多。进一步分析一次枝梗颖花数和二次枝梗颖花数所占比例后发现,I和Ⅱ期乙烯利处理的一次枝梗颖花数比例较对照显著降低,在Ⅲ和Ⅳ期处理则较对照显著增加;二次枝梗颖花数的比例则刚好相反。由此可见,乙烯利在幼穗分化的I和Ⅱ期处理,一次枝梗颖花数和二次枝梗颖花数均表现为降低,但主要以一次枝梗颖花数降低为主,二次枝梗颖花数降低较少,从而增加了二次枝梗颖花数在总颖花数中的比例,并成为总颖花数的主要组成部分。在幼穗分化的I和Ⅱ期多效唑处理较对照增加了一次枝梗颖花数,Ⅳ期处理则降低;二次枝梗颖花数在I、Ⅱ期多效唑处理较对照有增加,在Ⅳ期处理则出现降低。多效唑在各时期处理的一次枝梗颖花比例和二次枝梗颖花比例和对照无显著差异。
2.4 不同时期调节剂处理对水稻颖花分化的影响
如表3所示,在水稻幼穗分化的I、Ⅱ、Ⅲ和Ⅳ期乙烯利处理均显著增加了总颖花分化数,分别较对照增加了9.30%、7.88%、6.25%和5.39%;其中一次枝梗颖花分化数分别增加了8. 45%、7.48%、7.06%和6. 37%,二次枝梗颖花分化数分别增加9.70%、8.02%、5.69%和4.73%。穗分化各时期乙烯利处理的一次枝梗颖花分化数比例和二次枝梗颖花分化数比例与对照相比无显著差异。在幼穗分化的I、Ⅱ期多效唑处理显著增加了总颖花分化数,分别较对照增加了5.44%和2.85%,其中一次枝梗颖花分化数和二次枝梗颖花分化数均表现出增加;在Ⅲ和Ⅳ期处理则显著降低了总颖花分化数,其中一次枝梗颖花分化数在Ⅲ期处理为增加,Ⅳ期为降低,而二次枝梗颖花分化数Ⅲ和Ⅳ期处理均降低。一次枝梗颖花分化数比例在幼穗分化各时期均显著增加,而二次枝梗颖花分化数比例则表现相反,均为降低。这说明多效唑在幼穗分化的I、Ⅱ期处理,总颖花分化数表现的增幅主要是由一次枝梗颖花分化数增加引起,二次枝梗颖花分化数增幅较小。Ⅲ期多效唑处理,二次枝梗颖花分化数大量减少,一次枝梗颖花分化数少量增加;Ⅳ期多效唑处理则表现为一次枝梗颖花分化数和二次枝梗颖花分化数均下降,其中二次枝梗颖花分化数降幅较大。
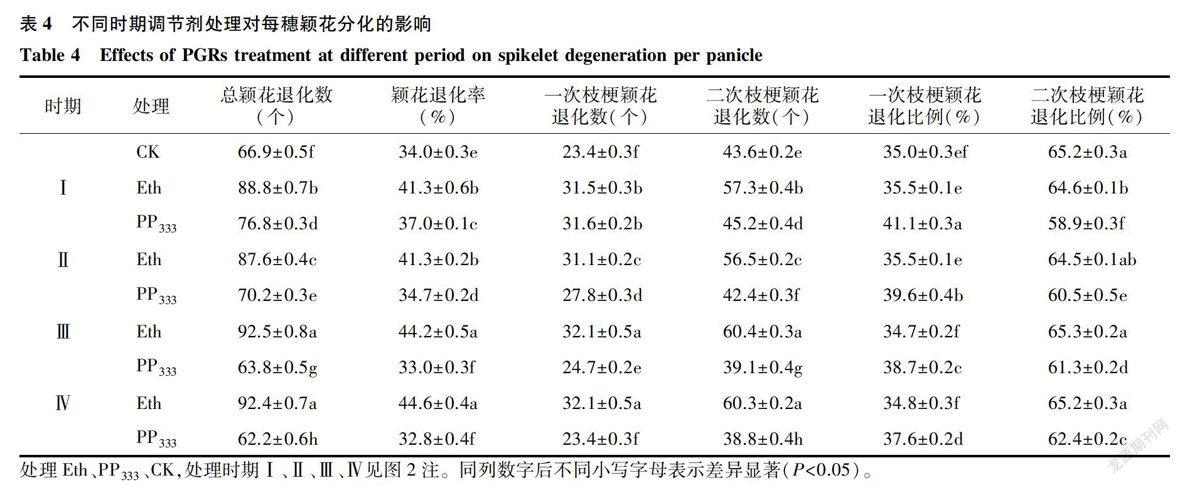
2.5 不同时期调节剂处理对水稻颖花退化的影响
如表4所示,乙烯利处理的总颖花退化数和退化率整体呈增加趋势,而多效唑处理则表现为I期和Ⅱ期增加、Ⅲ和Ⅳ期降低。與对照相比,乙烯利在幼穗各时期处理均增加了总颖花退化数、退化率以及一次枝梗颖花退化数和二次枝梗颖花退化数,其中以Ⅲ期处理增幅最大,分别达38.27%、30.11%、37.18%和38.53%。各时期乙烯利处理的一次枝梗颖花退化数比例和二次枝梗颖花退化数比例与对照均无显著差异。多效唑在I、Ⅱ期处理总颖花退化数和退化率均较对照增加,Ⅲ和Ⅳ期处理则下降;而一次枝梗颖花退化数在I、Ⅱ、Ⅲ期均较对照上升,二次枝梗颖花退化数除I期增加外,其余时期则下降。一次枝梗颖花退化数比例在各时期均高于对照,而二次枝梗颖花退化数比例则低于对照。表明,多效唑在I期和Ⅱ期处理时,一次枝梗颖花退化数的增幅大于二次枝梗颖花退化数的增幅,因此一次枝梗颖花退化数及其比例表现为增加,二次枝梗颖花退化数及其比例为下降;而在Ⅲ期和Ⅳ期多效唑处理,一次枝梗颖花退化数仍表现为增加,但二次枝梗颖花退化数则急剧减少,因而其总颖花退化数表现为降低。
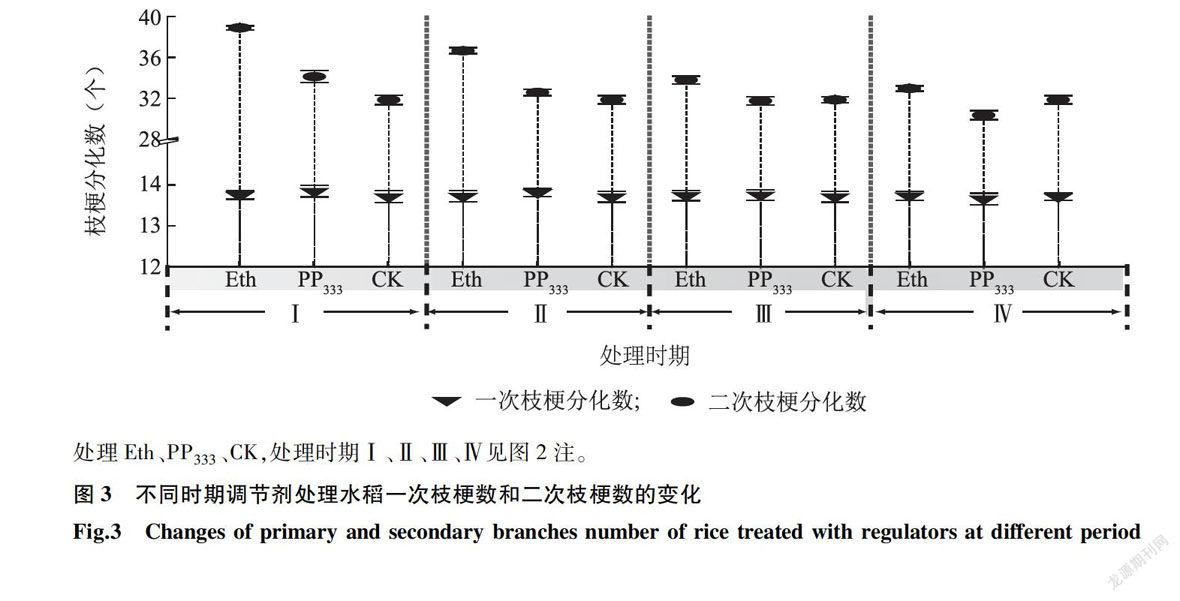
2.6 不同时期调节剂处理对水稻枝梗分化的影响
由图3可知,不同时期乙烯利和多效唑处理对水稻一次枝梗和二次枝梗分化的影响各异。在I、Ⅱ、Ⅲ、Ⅳ期乙烯利处理均增加了一次枝梗分化数,但与对照相比未达显著水平,二次枝梗分化数较对照增加显著。在I、Ⅱ和Ⅲ期多效唑处理均增加了一次枝梗分化数和二次枝梗分化数,Ⅳ期处理一次枝梗分化数和二次枝梗分化数则下降。
2.7 不同时期调节剂处理对水稻穗部性状的影响
如表5所示,不同时期乙烯利和多效唑处理的实粒数、结实率和千粒质量整体呈现下降趋势。乙烯利在幼穗分化各时期处理,实粒数、结实率和千粒质量均显著低于对照,其中以Ⅳ期处理降幅最大,达16.86%、7.17%和7.66%;而秕粒数的变化则完全相反,以Ⅳ期处理增幅最大,达82.93%。多效唑在I和Ⅱ期处理时,实粒数和结实率均显著高于对照;而在Ⅲ和Ⅳ期处理则刚好相反。I和Ⅱ期多效唑处理的秕粒数显著降低,达70.73%和45.12%,Ⅲ和Ⅳ期处理秕粒数则增加。
2.8 水稻一次枝梗颖花数、二次枝梗颖花数与总颖花数和实粒数的相关性
由图4可见,水稻一次枝梗颖花数和二次枝梗颖花数与总颖花数均呈正相关,相关系数分别为0.984和0.952,可见总颖花数与一次枝梗颖花数的相关性更紧密。水稻一枝梗颖花数和二次枝梗颖花数与实粒数也呈正相关,相关系数分别为0.968和0.995,由此可知实粒数与二次枝梗颖花数的相关性更紧密。
2.9 水稻穗部各性状间的相关性分析
由表6可知,结实率与一次枝梗颖花数、二次枝梗颖花数、每穗颖花数、一次枝梗分化数以及千粒质量呈极显著正相关;而与二次枝梗颖花退化数和总颖花退化数呈显著负相关。千粒质量与一次枝梗颖花数、二次枝梗颖花数、每穗颖花数、一次枝梗分化数呈极显著正相关;而与二次枝梗颖花退化数和总颖花退化数呈显著负相关。这说明一次枝梗颖花数、二次枝梗颖花数、每穗颖花数和一次枝梗分化数越多,水稻结实率越高、千粒质量越重;而二次颖花退化数或总颖花退化数越多,结实率就越低,千粒质量也越小。同时,一次枝梗分化数与一次枝梗颖花数、一次枝梗颖花分化数呈显著正相关,而与二次枝梗颖花数和二次枝梗颖花分化数呈负相关。说明一次枝梗分化数越大,一次枝梗颖花数和一次枝梗颖花分化数就越多,但二次枝梗颖花数和二次枝梗颖花分化数就越少。二次枝梗分化数与二次枝梗颖花数、一次枝梗颖花退化数、二次枝梗颖花退化数、总颖花退化数、二次枝梗颖花分化数和总颖花分化数均呈显著正相关,而与一次枝梗颖花数和一次枝梗颖花分化数呈负相关,因而当二次枝梗分化数增加时,二次枝梗颖花数、一次枝梗颖花退化数、二次枝梗颖花退化数、总颖花退化数、二次枝梗颖花分化数和总颖花分化数均会增加,而一次枝梗颖花数和一次枝梗颖花分化数则会降低。
3 讨论
水稻幼穗分化过程中,颖花的分化与退化是穗粒数的基础。研究结果显示,幼穗分化的枝梗及颖花分化期是决定每穗起始粒数的关键,也是决定每穗最高颖花数的时期,而减数分裂期是决定颖花退化数的关键时期,也是决定每穗总粒数的重要时期[19]。由此可见,颖花起始关键期即为本试验的Ⅱ期(枝梗一颖花分化期),颖花退化关键期为本试验的Ⅳ期(雌雄蕊形成一减数分裂期)。本试验中,乙烯利在幼穗分化期的不同时期使用均降低了每穗颖花的形成,但降低幅度不同,其中以颖花分化期至减数分裂期(Ⅲ期和Ⅳ期)对颖花形成的抑制作用最显著,可见乙烯利在颖花起始关键期使用对颖花数的减少作用小于乙烯利在颖花退化关键期使用。这说明颖花退化关键期较颖花起始关键期对乙烯利处理更敏感,且颖花退化关键期每穗颖花数的显著降低很有可能是颖花退化数增加引起的,多效唑处理则不同,I期和Ⅱ期多效唑处理均显著增加了每穗颖花数,尤以Ⅱ期最高;而其在Ⅳ期则显著降低了每穗颖花数。这表明颖花起始关键期和颖花退化关键期对多效唑处理均有显著响应,但效果不同。
赵长华等[20]研究指出,每穗颖花数的多少是由颖花分化数和颖花退化数共同决定的,而每穗颖花分化数又由每穗的一次枝梗和二次枝梗颖花分化数决定,颖花退化数则由一次、二次枝梗颖花退化数决定。本试验中,乙烯利不管是在前期(I和II期)处理还是后期(Ⅲ和Ⅳ期)处理,其总颖花分化数均显著高于对照,但由于其总颖花退化数的幅度也显著高于对照,因而每穗颖花数出现下降。这说明乙烯利处理可以促进颖花分化,但也会加速颖花退化,其在整个幼穗分化期处理效果均一致。具体来说,在幼穗分化前期(I和II期)处理,每穗颖花数的降低主要是一次枝梗颖花数减少引起的,而后期(Ⅲ和Ⅳ期)处理每穗颖花数降低主要以二次枝梗颖花数减少为主。多效唑在前期(I和II期)处理总颖花分化数和总颖花退化数均表现为增加,但分化数的增幅大于退化数,故每穗颖花数表现为增加;而在后期(Ⅲ和Ⅳ期)处理总颖花分化数和总颖花退化数均降低,故每穗颖花数表现为降低。这说明多效唑在幼穗分化不同时期处理效果不同,其既能促进颖花分化与退化也能减少颖花分化与退化。总的说来,乙烯利和多效唑在幼穗分化不同时期处理均可以通过调控一次和二次枝梗颖花的分化和退化情况来达到控制每穗颖花数的作用。
姚有礼等[18]研究结果表明,一次枝梗颖花分化数和二次枝梗颖花分化数分别主要取决于一次枝梗分化数、二次枝梗分化数。不同品种每穗二次枝梗分化数、每个一次枝梗上的二次枝梗分化数均主要决定于每穗一次枝梗分化数,故认为在品种间一次枝梗分化数是决定每穗颖花分化数的首要因子。本试验中,结实率与一次枝梗分化数显著相关,而与二次枝梗分化数呈负相关,说明一次枝梗分化数越多,结实率越高;但二次枝梗分化数越多,结实率会降低。這可能与二次枝梗退化数有关,尽管二次枝梗分化数会显著大于一次枝梗分化数,但其退化数也高,导致现存二次枝梗数少,从而造成结实率低。这点可从二次枝梗分化数与总颖花退化数呈显著正相关上看出,而一次枝梗分化数与总颖花退化数呈负相关。同时,一次枝梗分化数还与每穗颖花数、总颖花分化数和二次枝梗分化数呈正相关。这说明一次枝梗分化数的增加,每穗颖花数、总颖花分化数和二次枝梗分化数均会增加,这与姚有礼的研究结果一致。郑福余[21]研究指出,水稻二次枝梗颖花数一般占总颖花数的60%以上,但其成粒率和千粒质量却明显低于一次枝梗颖花,所以提高二次枝梗颖花的成粒率和千粒质量可显著增产。本研究中,结实率与二次枝梗颖花数的相关性较一次枝梗颖花数更紧密,表明二次枝梗颖花数越多,结实率就越高;但千粒质量与二次枝梗颖花数的相关性较一次枝梗颖花数低,说明二次枝梗颖花数对千粒质量的重要程度低于一次枝梗颖花数。这与郑福余的研究结果基本一致。本试验中,乙烯利处理显著减少了实粒数、增加了秕粒数,导致结实率和千粒质量显著降低;多效唑在前期(I和Ⅱ期)处理实粒数、结实率和千粒质量均有所增加,秕粒数下降;但在后期(Ⅲ和Ⅳ期)处理,产量性状则完全相反。由此可知,幼穗分化不同时期不同外源植物生长调节剂处理对水稻一次、二次枝梗颖花分化与退化、实粒数形成以及结实率和千粒质量等产量性状的影响不一致,乙烯利处理减少了每穗颖花数和实粒数,增加空秕粒数,从而降低了结实率和千粒质量,导致产量降低;多效唑处理对现存颖花数、实粒数、结实率和千粒质量的影响随着处理时期的不同而不同。
参考文献:
[l] 王惠芝.氮素穗肥调控水稻颖花数形成的生理机制研究[D].南京:南京农业大学,2007.
[2] 凌启鸿,蔡建中,苏祖芳.叶龄余数在稻穗分化进程鉴定中的应用价值[J].中国农业科学,1980,13(4):1-11.
[3]王亚梁,张玉屏,曾研华,等.水稻穗分化期高温对颖花分化及退化的影响[J].中国农业气象,2015,36(6):724-731.
[4] 曾研華,张玉屏,向镜,等,籼型常规早稻穗分化期低温对颖花形成和籽粒充实的影响[J].应用生态学报,2015,26(7):2007-2014.
[5]李刚华,王惠芝,王绍华,等,穗肥对水稻穗分化期碳氮代谢及颖花数的影响[J].南京农业大学学报,2010,33(1):1-5.
[6]杨洪建,王余龙,黄建晔,等.开放式空气C02浓度增高对水稻颖花分化和退化的影响[J].应用生态学报,2002,13(10):1215-1218.
[7]阿加拉铁,薛大伟,李仕贵,等.植物激素与水稻产量的关系[J].中国稻米,2006,13(5):1-3.
[8]董庆坤,刘慧丽.植物激素调控籽粒大小的研究进展[J].华南师范大学学报(自然科学版),2015,47(6):72-78.
[9]黄静静,王绍华,李刚华,等.6-苄基腺嘌呤对水稻颖花分化影响机制的研究[J].南京农业大学学报,2009,32(3):8-13.
[10]顾大路,王伟中,王红军,等,乙烯利在水稻中后期应用的效果研究[J].江苏农业科学,2003,31(5):26-28,
[11]宫彦龙,徐海,夏原野,等.幼穗分化期喷施表油菜素内酯( epi-BR)对水稻穗部性状的影响[J].作物杂志,2016,32(2): 133-138.
[12]宫彦龙,雷月,夏原野,等.幼穗分化期喷施细胞分裂素(CTK)对水稻穗部性状及株型性状的影响[J].作物杂志, 2017,33(5):112-118.
[13]闫志强,徐海,宫彦龙,等.籼粳稻的花时对乙烯利调控的响应及其敏感性差异[J].沈阳农业大学学报,2015,46(6): 641-647.
[14]李宗霆,周燮.植物激素及其免疫检测技术[M].南京:江苏科学技术出版社,1993:101-102.
[15]闫凯莉,韩云,谭廷钢,等.15%多效唑可湿性粉剂对水稻生长、产量及品质的影响[J].热带农业科学,2016,36(2):73-76.
[16]孙克新.多效唑调节杂交稻制种花期的效应及其解除[J].杂交水稻,2001,16(1):17-18.
[17]赫明涛,王军,孙扣忠,等.水稻制种花期相遇的调节技术[J].江苏农业科学,1999,27(4):11-12.没有卷号。
[18]姚友礼,王余龙,蔡建中,等,水稻大穗形成机理的研究——穗肥施用时期对每穗颖花分化、退化和现存数的影响[J].江苏农学院学报,1997,18(4):29-35.
[19]姜辉,姜树坤,陈丽丽,等.水稻顶端颖花退化的形态与发育学研究[J].黑龙江农业科学,2016,39(1):7-11.
[20]赵长华,丁艳峰.水稻穗粒数形成的生理生化研究进展[J].耕作与栽培,2001,21(1):5-8.
[21]郑福余.水稻二次枝梗颖花的灌浆程度对产量的影响[J].黑龙江农业科学,1997,20(4):16-18.
猜你喜欢
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农产品市场周刊(2017年35期)2017-10-25
湖北农业科学(2016年23期)2017-03-17
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
天津农业科学(2016年4期)2016-04-20
安徽农学通报(2015年10期)2015-06-15
湖北农业科学(2014年9期)2014-08-08
