
银耳芽孢和菌丝的转录组分析
2019-10-15 08:36朱涵予
食品与机械 2019年9期
陈 政 朱涵予 -
(衡阳师范学院生命科学与环境学院,湖南 衡阳 421000)
银耳是一种可食用的胶质菌,具有极高的食用、药用及经济价值,历来受到中国及东南亚国家人们的喜爱[1]。银耳中富含多糖、蛋白质、维生素、微量元素、黄酮类和多酚类营养物质[2-3]。佀国涵等[4]分别提取银耳芽孢、菌丝体和子实体中的凝集素,发现3种材料中凝集素的凝集活性和稳定性均不相同。Li等[5]提取了银耳子实体中主要的酚酸类化合物如4-对羟基苯甲酸、龙胆酸、4-香豆酸并测定其抗氧化活性。刘洋等[6]建立了银耳芽孢总蛋白的提取方法。还有大量研究[7-9]通过改变银耳菌丝体和芽孢的发酵条件以提高银耳多糖产量。银耳芽孢具有易培养、生长速度快的优点,可运用于食品领域,已开发了许多产品如银耳芽孢发酵液、芽孢多糖粉[10]。而在营养成分方面的研究主要集中在银耳菌丝体和子实体上,对芽孢研究鲜有报道[9],且仅针对银耳多糖,此领域空白较多。
RNA-Seq可准确研究所有mRNA的丰度信息,从RNA水平分析细胞代谢和基因表达与一些生命现象的关系,通过对转录组差异的研究,得到大量的关于功能基因、代谢途径的相关信息[11-12]。通过研究银耳芽孢和菌丝整体基因的表达变化,可进一步了解银耳中营养物质的代谢途径,挖掘功能基因。RNA-Seq在双孢蘑菇[13]、牛樟芝[14]、冬虫夏草[15]、灵芝[16]、茯苓[17]、灰树花[18]等食药用真菌中均有应用。
试验选用银耳双核菌丝M1332及其2个单核亲本芽孢Y13、Y32为材料,运用RNA-Seq进行比较分析,全面探讨银耳芽孢到菌丝生长过程中的基因表达量的变化,分析此过程涉及的代谢途径及相关功能基因,以期为银耳的代谢调控、功能基因的克隆等提供依据。
1 材料与方法
1.1 材料与仪器
1.1.1 材料与试剂
银耳双核菌丝M1332及其2个单核亲本芽孢Y13、Y32:华中农业大学食品科学技术学院食品微生物实验室保藏;
马铃薯葡萄糖琼脂(PDA)培养基:青岛高科园海博生物技术有限公司;
三氯甲烷、乙醇、异丙醇:分析纯,国药集团化学试剂有限公司;
焦炭酸二乙酯(DEPC):分析纯,上海西格玛奥德里奇贸易有限公司;
RNaisoTMplus:大连宝生物工程有限公司;
TransScript®first-strand cDNA Synthesis Supermix反转录试剂盒、TransStart®Green qPCR SuperMix试剂盒:北京全式金生物技术有限公司。
1.1.2 仪器与设备
高速离心机:5418R型,德国Eppendorf AG公司;
Nanodrop分光光度计:NanoDrop 3300型,美国Thermo Scientific公司;
荧光定量PCR仪:ABI Prism7900型,美国应用生物系统公司;
电泳仪:DYCP-31DN型,北京市六一仪器厂。
1.2 方法
1.2.1 总RNA提取 PDA培养基上培养菌丝20 d,芽孢培养7 d后收集菌体提取RNA。采用RNaisoTMplus试剂盒提取总RNA,详细操作参照产品说明书。通过琼脂糖凝胶电泳、测定A260/280和A260/230以检测RNA质量。检测合格的总RNA委托深圳华大基因科技有限公司进行 Illumina HiseqTM4000测序。
1.2.2 转录组文库的数据组装 测序所得raw reads数据,过滤去除低质量、包含接头污染及未知碱基含量>5% 的reads,得到clean reads。利用Trinity(version 2.0.6)软件对clean reads进行denovo组装[19]。通过序列之间的overlap信息组装得到contigs,根据序列的Paired-end信息将contigs组装得到转录本序列。最后用Tgicl(version 2.0.6)进行聚类去冗余得到Unigene。
使用Blastx对Unigene进行NR、NT、Swiss-Prot、COG、KEGG注释,使用InterProScan5(version 5.11)进行InterPro注释,使用Blast2GO(version 2.5.0)以及NR注释结果进行GO(Gene Ontology)注释。根据功能注释结果,按Blastdb优先级确定Unigene的CDs序列。以上数据库未能得到注释的Unigene则使用ESTScan(version 3.0.2)进行预测确定其序列及方向。
1.2.3 差异表达基因的KEGG富集分析 每个样品采用FPKM(reads per kb per million reads)方法计算基因的表达量。并用倍数变化法筛选两样本之间的差异表达基因,筛选标准为:错误发现率(FDR)≤0.01且倍数差异≥2。
将3个样本间获得的差异表达基因根据KEGG注释的结果以及官方分类,将差异表达基因进行生物通路分类,用R软件中的phypen函数进行富集分析,筛选出显著性富集的通路Pathway(定义q值<0.05为在差异表达基因中显著富集的通路)。
1.2.4 实时荧光定量(qRT-PCR)检测分析 不同样品的总RNA反转录后采用TransStart®Green qPCR SuperMix试剂盒进行qRT-PCR,具体试验步骤参照产品说明书。反应条件为94 ℃预变性30 s,再进行30个循环,包括94 ℃ 5 s,60 ℃ 15 s,72 ℃ 10 s,于ABI Prism7900荧光定量PCR仪上进行。以银耳β-tubulin作为内参,绘制溶解曲线,最终数据以2-ΔΔCt进行分析。设计的基因及其引物序列分别为β-tubulin(F:5′-GATGACCATTTCTTGCTTC-3′,R:5′-GTTCTGACATTTGCTACCG-3′);CL131.contig1(F:5′-CCGCCTTGGTCTTCCTCATT-3′,R:5′-TAGTTGCTCCCGCCCTTGTA-3′);CL2725.contig1(F:5′-GGCAGCCGACAAGCGACATA-3′,R:5′-ACCACCGCAGGCGA-CGAAAT-3′);unigene3454(F:5′-CTCTCGTCTACAAGGGCTCC-3′,R:5′-GAGGAAGAAAGGGCGA-ACTG-3′),委托武汉天一辉远公司合成引物。
2 结果与分析
2.1 银耳转录组数据的产出及组装
建库测序单个样品测序深度>4.24 Gb,获得13.16 Gb碱基序列信息,各样品的测量结果准确度Q30(序列的测序错误率低于0.1%的比例)>93.04%,GC含量57.95%~58.10%,clean reads占raw reads的比例>86.27%,过滤后reads>28.25 Mb(表1),说明测序质量较好,数据可用于进一步组装分析。
利用Trinity(version 2.0.6)软件对过滤后的所有clean reads序列进行denovo组装,获得的转录本通过Tgicl进一步去冗余筛选,得到的Unigene共计16 190条,总的长度27 939 037 bp,平均长度1 725 bp,GC含量57.98%,N50(指从组装最长的Unigene依次向下求长度的总加和,当累加长度达到组装长度的1/2时,对应的Unigene长度为N50长度)为2 541 bp。对组装的Unigene长度进行统计,大于3 000 bp的2 558条,小于3 000 bp 的片段分布均匀,数目分布上与N50结果较为一致,说明denovo组装结果较好。
2.2 Unigene的注释
组装完毕后,对Unigene进行NR、NT、GO、COG、KEGG、Swissprot和Interpro数据库注释,结果显示,12 458条在NR数据库里匹配到同源基因,占所有Unigene的76.95%;其他数据库依次为32.95%,59.09%,61.71%,44.49%,53.93%,24.22%。以NR数据库为例进行分析,结果显示,有12 458条Unigene在NR数据库中可找到相似序列。其中,55.75%的Unigene比对到金黄银耳(T.mesenterica),13.35%比对到新型隐球酵母(Cryptococcusneoformans)。根据注释结果共检测出12 366个CDs,未注释上的Unigene使用ESTScan预测后获得975个CDs。同时还检测出1 105个SSR位点,以及预测出553个编码转录因子的Unigene。
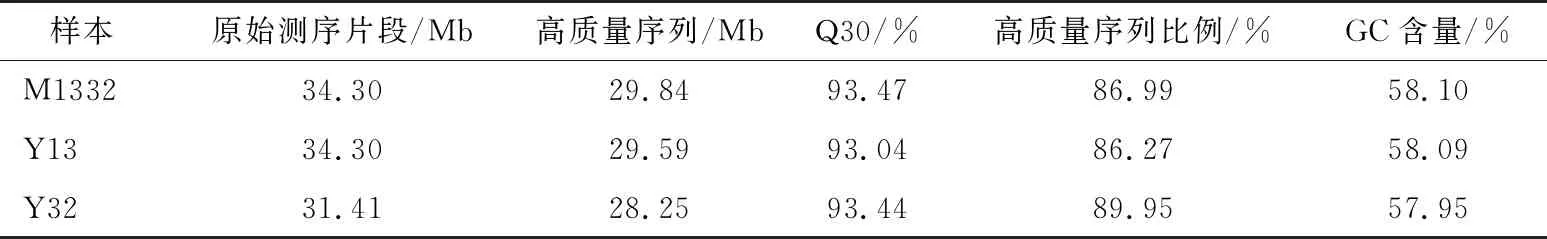
表1 各样本测序产出数据†
† Q30:序列的测序错误率<0.1%的比例。
2.3 COG注释
将所有Unigene与COG数据库比对,结果显示,共有7 203条Unigene在COG数据库中找到同源信息,共被分成25大类(图1),其中常规功能预测最多,共有2 339条,占所有分类的32.47%。其次有1 588条预测到碳水化合物的运输和代谢,为银耳中碳水化合物如银耳多糖的研究提供基础。核酸结构预测得到的最少,仅有2条,其他类别的基因表达丰度各不相同。
2.4 GO分析
结合GO数据库对所有Unigene进行功能分类,结果显示,共有3 921条Unigene被注释到了GO分类上(图2)。其中样本基因数量在1 000条以上的生物过程分类中主要聚集于代谢过程(1 870个)、细胞过程(2 032个)和单一有机体的过程(1 454个);在细胞组分主要聚集于细胞(1 706个)、细胞成分(1 696个)、细胞器(1 177个)和生物膜(1 184个);在分子功能分类中主要聚集于蛋白结合(1 227个)和催化活性(1 614个)。

A. 翻译,核糖体结构和生物合成 B. 转录 C. 信号转导机制 D. 次级代谢产物的合成、转运和代谢 E. RNA加工与修饰 F. 复制、重组和修复 G. 翻译后修饰、蛋白质转换、伴侣 H. 核苷酸转运与代谢 I. 核结构 J. 脂质转运与代谢 K. 细胞内的转运、分泌和囊泡运输 L. 无机离子转运与代谢 M. 一般功能预测 N. 未知功能 O. 胞外结构 P. 能量产生与转化 Q. 防御机制 R. 细胞骨架 S. 辅酶转运与代谢 T. 染色质结构与动力学 U. 细胞壁/膜/膜的生成 V. 细胞运动 W. 细胞周期调控、细胞分裂、染色体分隔 Y. 碳水化合物转运和代谢 Z. 氨基酸转运与代谢
图1 银耳Unigene的COG分类
Figure 1 COG functional classifications of all Unigene
2.5 差异表达基因KEGG富集分析
银耳菌丝M1332与芽孢Y13、Y32相比,上调的基因分别为2 296,3 533个,下调的基因分别为3 207,2 754个,芽孢Y13、Y32相比,上调的基因为3 803个,下调的基因为1 773个(图3)。
将鉴定出来的差异表达基因,进行KEGG富集分析。结果显示,7个通路富集水平显著,包括细胞内噬、代谢途径、减数分裂、氨基糖和核酸糖的代谢、酪氨酸代谢、精氨酸和脯氨酸的代谢、丙氨酸和天冬氨酸以及谷氨酸的代谢(表2)。

1. 生物阶黏附 2. 生物调节 3. 细胞组分的组织与生物合成 4. 细胞过程 5. 发育过程 6. 定位建立 7. 生长 8. 定位 9. 运动 10. 代谢过程 11. 多细胞有机体过程 12. 多细胞生物过程 13. 生物过程正向调控 14. 生物过程负向调控 15. 生物过程调控 16. 生殖 17. 生殖过程 18. 应激反应 19. 信号 20. 单机体过程 21. 细胞 22. 细胞组分 23. 胞外区 24. 胞外区组分 25. 大分子复合物 26. 膜 27. 膜组分 28. 膜封闭内腔 29. 拟核 30. 细胞器 31. 细胞器组分 32. 病毒 33. 病毒组分 34. 抗氧化活性 35. 连接 36. 催化活性 37. 电子载体活性 38. 酶调节活性 39. 鸟苷酸交换因子活性 40. 分子传感器活性 41. 核酸结合转录因子活性 42. 蛋白结合转录因子活性 43. 蛋白标签 44. 受体活性 45. 结构分子活性 46. 转运活性
图2 银耳Unigene的GO分类
Figure 2 Gene ontology classifications of all Unigene

FDR:错误发现率 FC:差异倍数。
通过对菌丝M1332、芽孢Y13、Y32的KEGG富集分析,差异表达基因主要富集在氨基酸代谢通路上,说明在两亲本芽孢配对形成菌丝的过程中有许多氨基酸代谢相关基因协同调控参与。而芽孢Y13、Y32的差异基因KEGG富集分析的结果显示,此过程中两芽孢的基因表达情况既有协同又有分工。
菌丝与芽孢相比,差异基因均有富集在氨基糖和核酸糖代谢途径上,说明在芽孢形成菌丝的过程中两芽孢均有基因参与此代谢途径。氨基糖作为微生物的次级代谢产物,是许多生物大分子如抗生素、壳聚糖、糖蛋白、脂多糖和黏多糖等的组成成分。在氨基转移酶的作用下,L-谷氨酸或谷氨酰胺上的氨基取代了磷酸糖或核酸糖上的羟基而形成氨基糖。被氨基取代后,糖的理化性质和生物功能会发生显著变化[20]。核酸糖,因其能形成高键能的糖苷键(ΔG°>-29.288 kJ/mol)而广泛参与多糖、糖蛋白、蛋白聚糖、脂多糖等的合成[21]。氨基糖和核酸糖代谢途径的相关转录组信息为今后银耳中生物大分子的代谢研究提供参考。
此外,大部分差异基因富集在氨基酸代谢通路中。谷氨酰胺酶能将谷氨酰胺脱氨后形成谷氨酸,经过一系列还原反应生成脯氨酸和天冬氨酸。谷氨酰胺来源的谷氨酸还可以通过供给氨基的方式参与其他氨基酸的合成,如丙氨酸[22]。研究[23-24]显示,癌细胞中谷氨酰胺酶的含量比正常细胞高,小鼠模型中谷氨酰胺酶基因的缺失推迟了肝癌细胞的肿瘤形成。氨基酸合成相关基因还参与其他生物学过程如KEGG富集得到的代谢通路减数分裂中。灰盖鬼伞的脯氨酸富集蛋白基因参与减数分裂过程,该基因的突变体具有担孢子产量减少,孢子存活率低且无法进行完整的减数分裂过程的现象[13]。
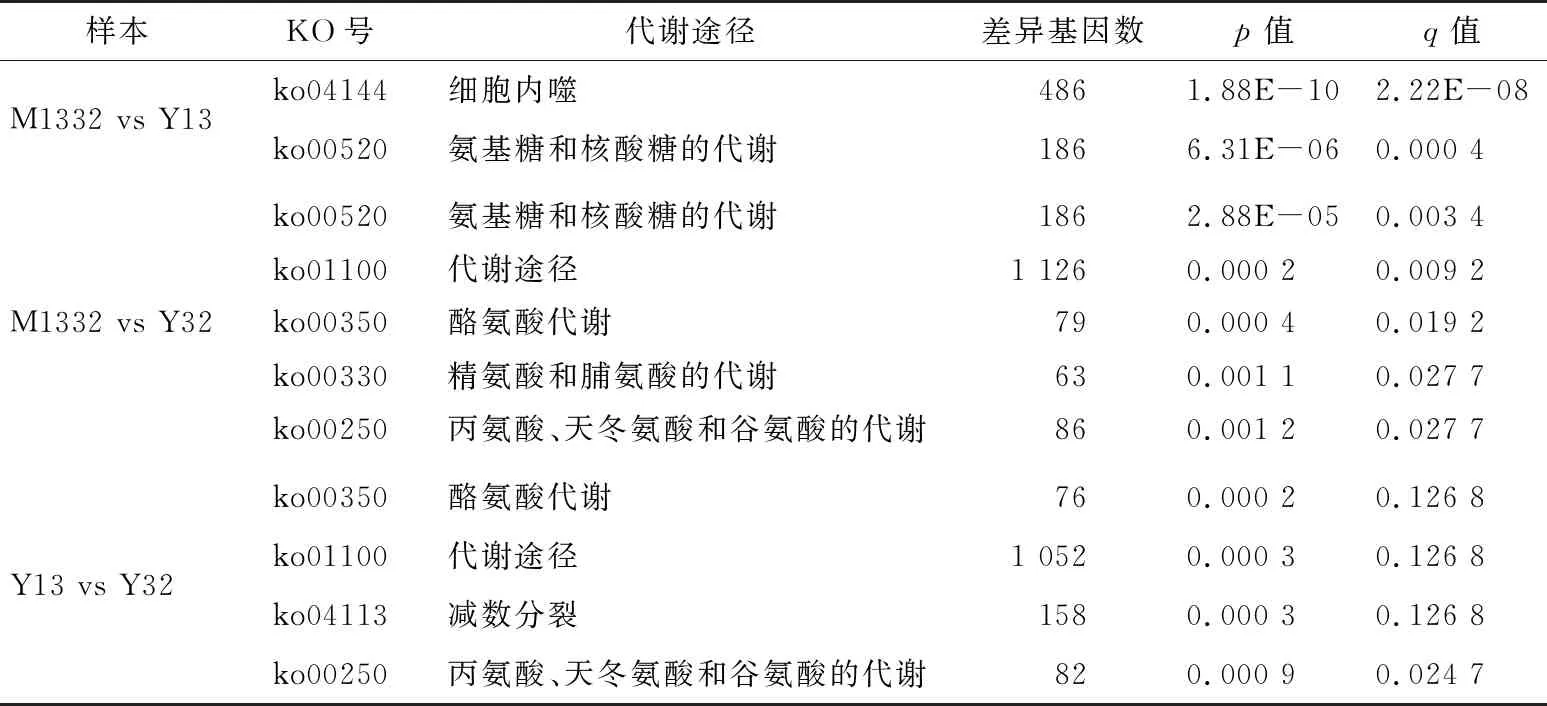
表2 KEGG通路富集分析†
†q值越小表示该通路富集水平越显著,p值越小表示在该通路可信度越高。
2.6 qRT-PCR表达分析
随机挑选3个KEGG富集分析结果中的差异表达基因,设计定量引物,验证相关基因在转录水平的表达。定量结果表明,qRT-PCR验证的3个基因在3个材料中的表达量改变倍数与转录组测序的结果一致(图4),测序结果较为准备可信,可供后续研究参考。

图4 差异基因的定量表达分析
3 结论
试验对银耳菌丝和芽孢进行转录组测序,了解了芽孢配对形成菌丝这一过程中全部基因的表达情况,获得了大量相关的转录本信息。转录组数据的深入挖掘有利于全面了解银耳菌丝和芽孢中基因信息和表达差异情况,今后应开展银耳在菌丝和芽孢两个时期的多糖和氨基酸等的合成和代谢途径相关差异表达基因的克隆和基因功能方面的研究。
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
湖南饲料(2021年4期)2021-10-13
浙江农业学报(2021年5期)2021-05-31
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
海峡姐妹(2018年8期)2018-09-08
娃娃乐园·综合智能(2018年5期)2018-04-12
新农业(2017年15期)2017-09-11
家庭医药·快乐养生(2017年6期)2017-06-16
