
流域水文变化对胡杨荒漠河岸林林窗及形成木特征的影响
2019-10-25 03:38阿依加马力克然木玉米提哈力克塔依尔江艾山买尔当克依木祖皮艳木买买提艾力亚尔艾尼瓦尔
生态学报 2019年17期
阿依加马力·克然木,玉米提·哈力克,*,塔依尔江·艾山,买尔当·克依木,祖皮艳木·买买提,艾力亚尔·艾尼瓦尔
1 新疆大学资源与环境科学学院,乌鲁木齐 830046 2 新疆维吾尔自治区绿洲生态教育部重点实验室,乌鲁木齐 830046 3 中国科学院生态环境研究中心,城市与区域生态国家重点实验室,北京 100085
林窗,也称为林隙或林冠空隙,是森林生态系统中普遍存在的小尺度干扰形式[1-2]。林窗研究作为森林循环理论的基础之一,具有重要的理论和实际意义[3-4]。Watt首次提出林窗一词至今[5],林窗特征及其动态研究已取得很大进展,并成为当前森林生态学研究的热点之一[6-7]。朱教君和Hu等学者基于自然干扰过程的研究成果,建立了林窗直径与林窗边缘木高度的关系[2,8]。虽然,国内外学者对热带[9-10]、亚热带[11]、北温带[12-13]到寒温带[14]等不同气候区主要森林类型林窗形成机制进行了研究,但对暖温带干旱区林窗方面的研究很罕见[1]。
胡杨(PopuluseuphraticaOliv.)是杨柳科(Salicaceae)杨属中最古老的原始树种之一,是荒漠河岸林重要的建群种,也是珍贵的抗逆种质资源[15]。它已成为抵御风沙、遏制沙化、维护区域生态平衡、保护生物多样性和保障绿洲农牧业生产的重要屏障,在维持干旱区生态系统结构和功能方面起到关键作用[16]。至今,诸多学者针对塔里木河流域天然胡杨林的发生发育规律及生理生态特性做了多方面的研究,旨在力求揭示胡杨个体、种群、群落及生态系统不同尺度上的适应策略与调控机制[17-20],使得干旱区内陆河流域生态水文过程研究得以重要进展,从而提高了我国在干旱区受损生态系统相关研究领域的国际知名度[21-22]。然而,针对荒漠河岸林演替与更新的主要驱动因素——林窗干扰及其对流域水文过程响应机理的研究尚为欠缺。本文选取塔里木河中游具有代表性的6个样点予以研究,通过地面监测获取林窗及其形成木特征参数,分析了不同水文条件下林窗数量及其形成木种群结构特征,探讨林窗及其形成木对流域水文变化的响应,揭示胡杨种群的死亡机理,以期为塔里木河流域天然胡杨林的恢复与生态保护提供理论依据和科学参考。
1 研究区概况
研究区位于塔里木河中游轮台县境内国家胡杨林自然保护区英巴扎断面(图1),地理坐标为41°10′25″—41°16′00″N和84°15′18″—84°23′00″E,属暖温带极端干旱大陆性荒漠气候,多年平均气温9.7 ℃,最高气温43.6 ℃,最低气温-30.9 ℃;年平均降水量30—34 mm,年平均蒸发量却高达2400—2900 mm[23]。年均日照时数为 2778 h,≥ 10 ℃ 年积温为4039 ℃,多风沙、浮沉天气,最大风速达40 m/s[24]。英巴扎断面处于塔里木河中游的起点,河道过水情况良好,洪水季节6—8月河岸林常受到洪水漫淹[25]。该区域是我国胡杨林面积最大、受人类破坏较少的原始森林分布区[1]。然而,水热值的极端现象限制了植物种类的多样性,使得植物群落与生态系统的结构十分简单[26]。研究区代表性的植物有胡杨(Populuseuphratica)、灰杨(Populuspruinosa)、多枝柽柳(Tamarixramosissima)、黑果枸杞(Lyciumruthenicum)、铃铛刺(Halimodendronhalodendron)、芦苇(Phragmitesaustralis)、罗布麻(Apocynumvenetum)、花花柴(Karelinacaspica)、疏叶骆驼刺(Alhagisparsifolia)、甘草(Glycyrrhizauralensis)等。

图1 研究区域示意图Fig.1 Study area
2 材料与方法
2.1 样地设置与地面监测
野外工作于2017年5月10日至6月10日进行,考虑到塔里木河丰水和枯水发生特点以及塔里木河流域管理局布设的地下水监测井分布情况,依据离河道距离、胡杨分布密度以及国内外林窗判定标准,在英巴扎断面选定50 m × 50 m栅格单元的6个地面监测样地。在每个样地内均监测了10个林窗,分别测定林窗特征参数(高度,面积,形状)以及形成木株数、树高(TH)、胸径(DBH)、冠幅(CD)等树形指标(林窗内没有形成木的记为“0”)。同时获取每个样地附近的地下水监测井地下水埋深数据。
2.2 水文数据收集
1955年建站以来至2016年的径流量、耗水量等数据来自于塔里木河上游水流量进入中游的控制点——英巴扎水文站。1990—2016年的地下水埋深数据取自塔里木河流域管理局在研究区按水分梯度布设的49眼地下水监测井。
2.3 数据分析
2.3.1林窗年龄的估算
林窗年龄(Gap age)系指林窗自形成以来至调查时的年数[27]。由于调查时条件的限制,以林窗内最先倒下的一株形成木(形成林窗的腐木)的腐烂程度结合保护区周围采伐树桩的腐朽状况并参考当地林业工作者的实践经验来估算林窗年龄[28]。由此,参照前人的研究方法[29],把腐木的腐朽级别划分为9个等级,见表1。
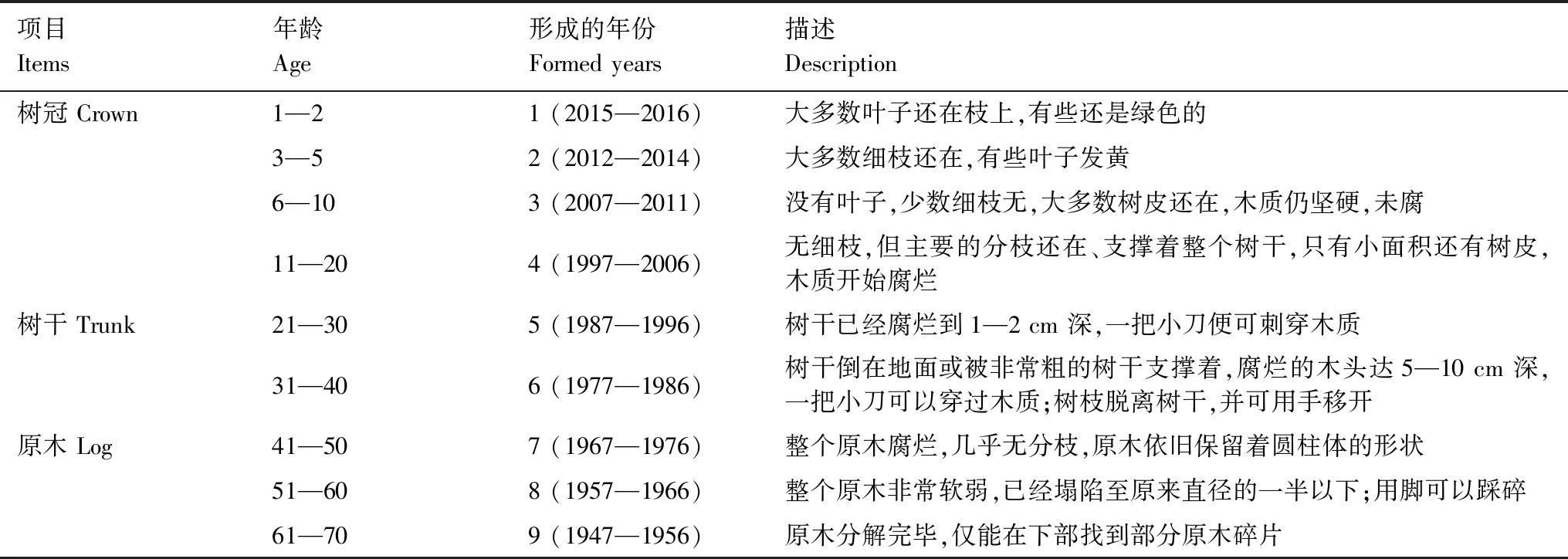
表1 腐木腐朽级别与林窗年龄的对应关系
2.3.2林窗形成木径级(年龄)划分与分布格局
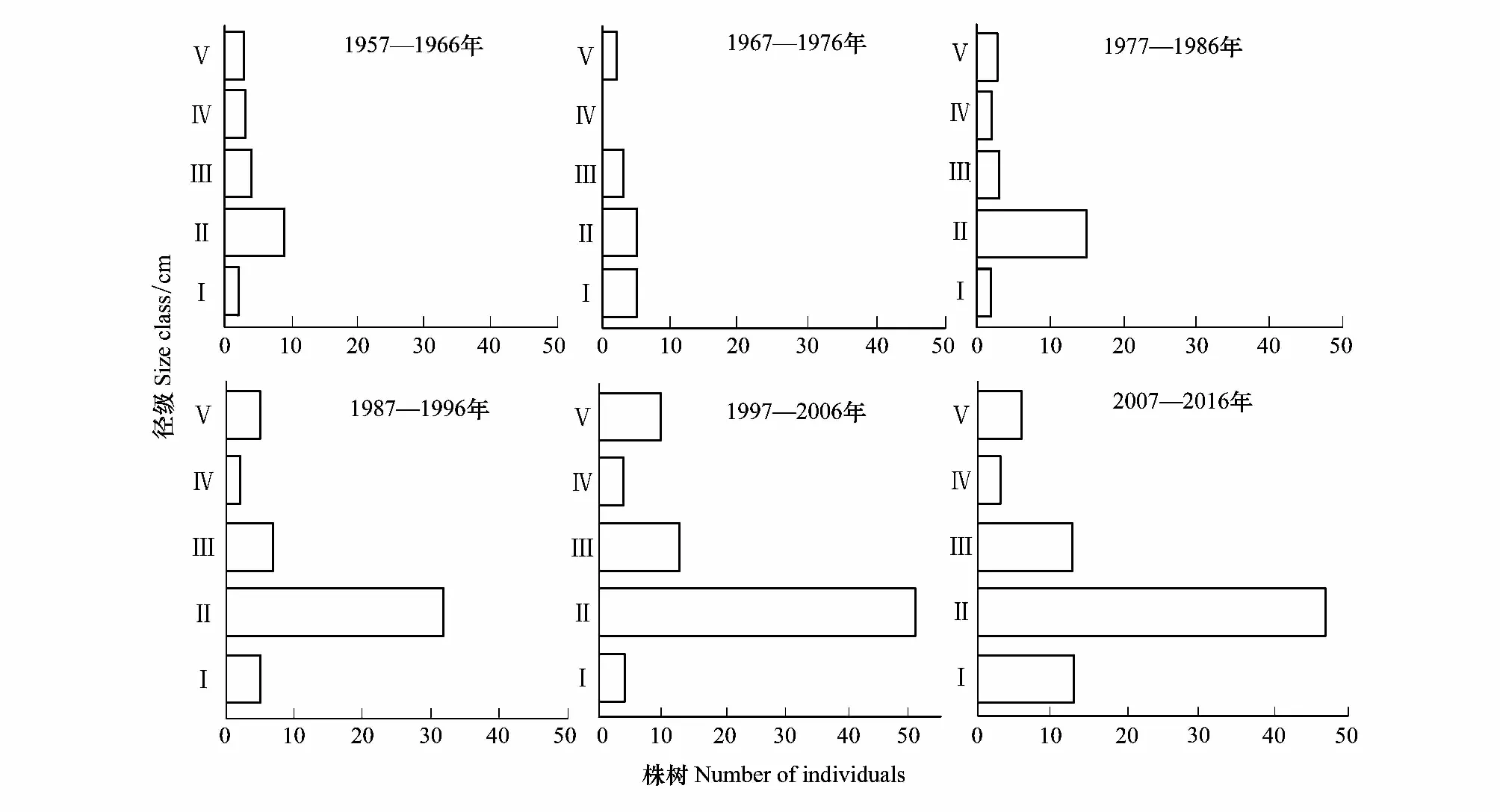
采用林窗乔木形成木径级结构代替其年龄结构的方法,将乔木形成木划分为5个径级:Ⅰ级幼龄树:胸径<5 cm,高度<4 m;Ⅱ级幼龄树:5 cm<胸径<10 cm,高度≥4 m;Ⅲ级中龄树:10 cm<胸径<15 cm;Ⅳ级中龄树:15 cm<胸径<20 cm;Ⅴ级大龄树(或老龄树):胸径>20 cm。按上述标准用拟合优度t检验法统计各年际不同径级林窗形成木胡杨株数,以径级为横坐标,林窗形成木数量出现频率为纵坐标,做出林窗形成木胡杨径级结构分布图,并判定塔里木河中游不同年代的林窗形成木种群总个体数与不同径级分布类型[30]。
2.3.3林窗形成木种群结构动态指数
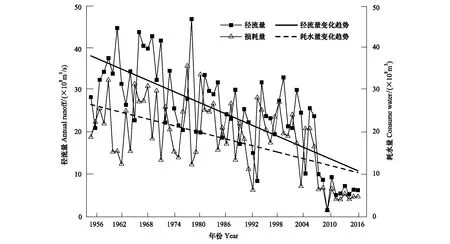
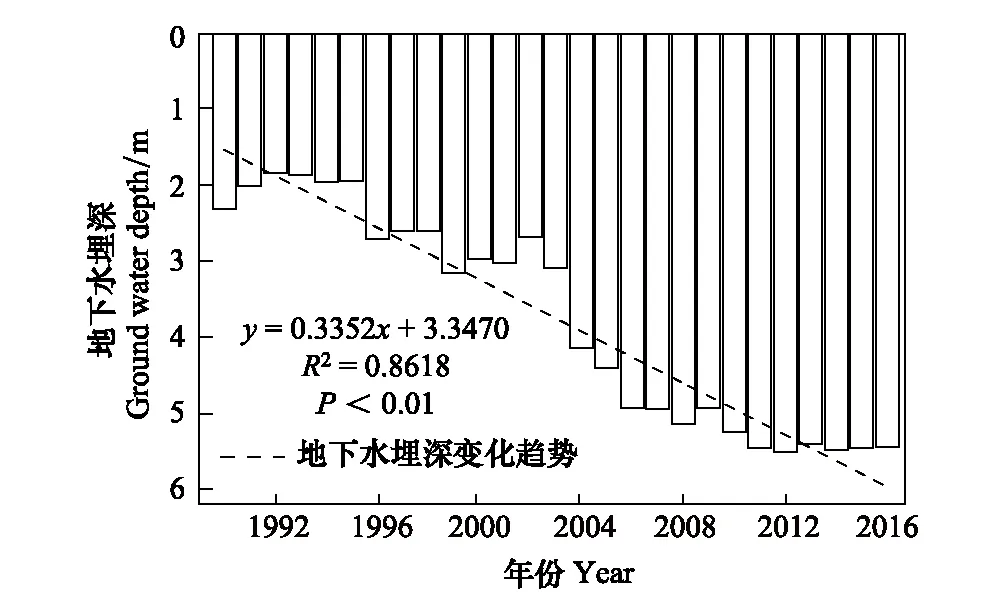
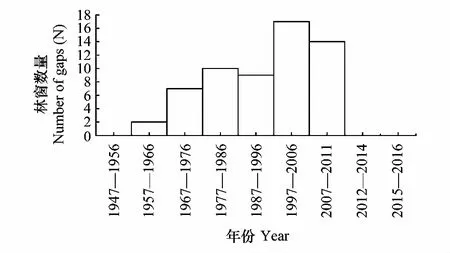
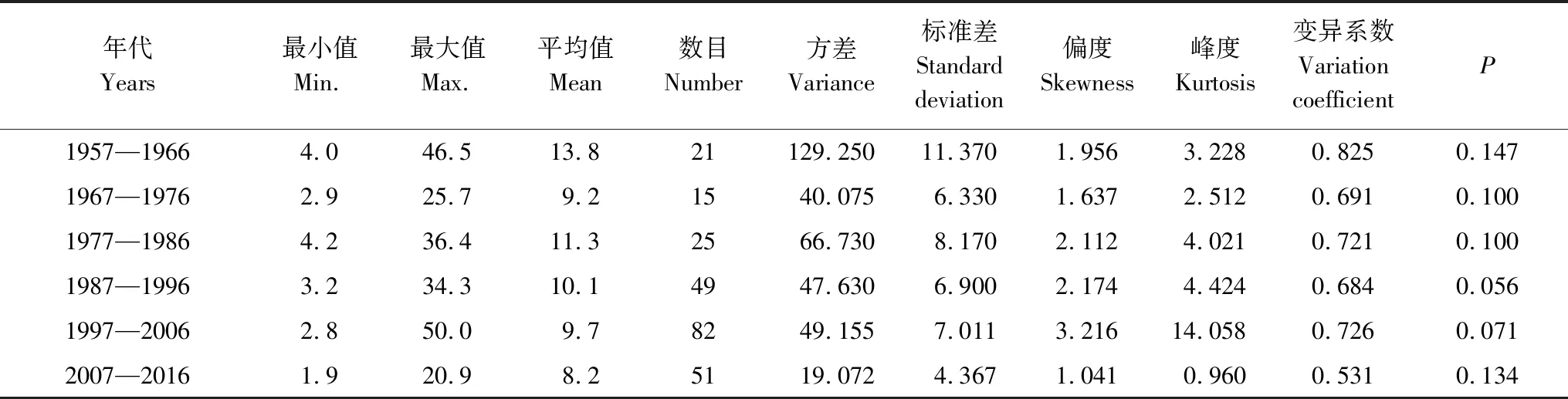
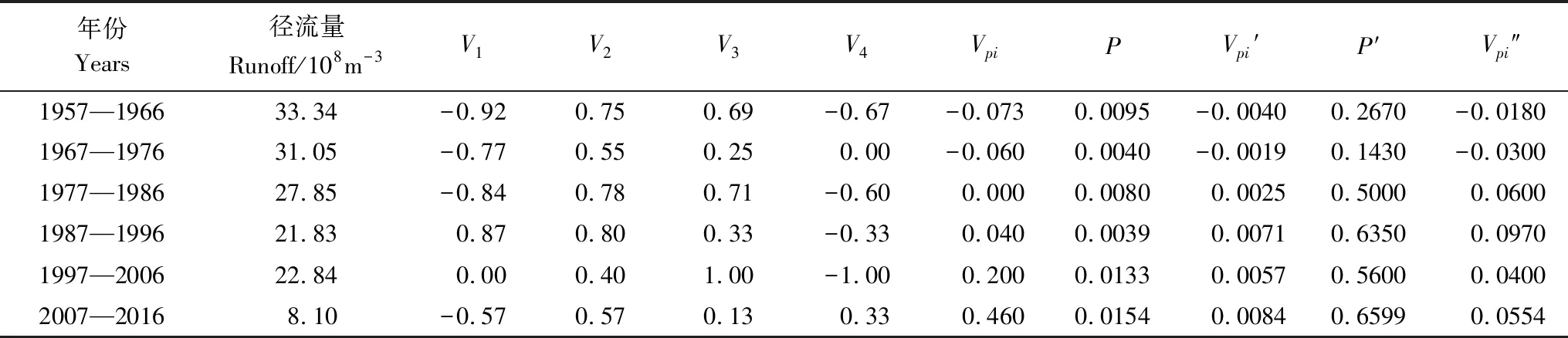
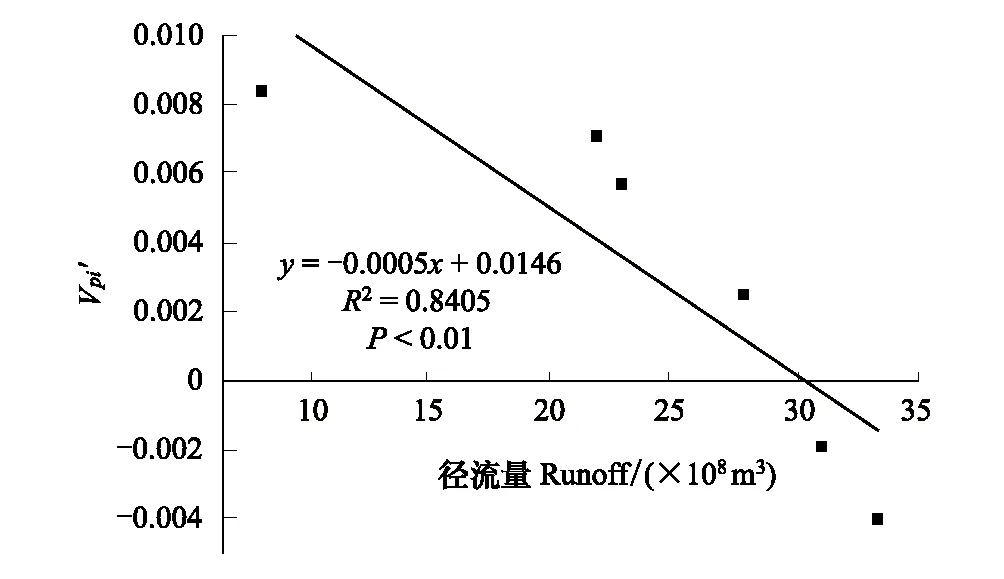
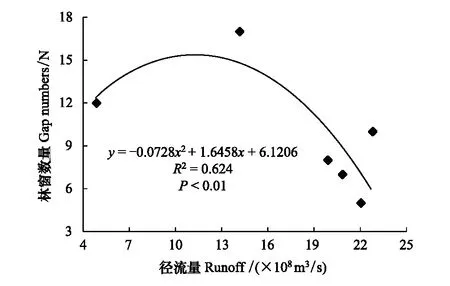
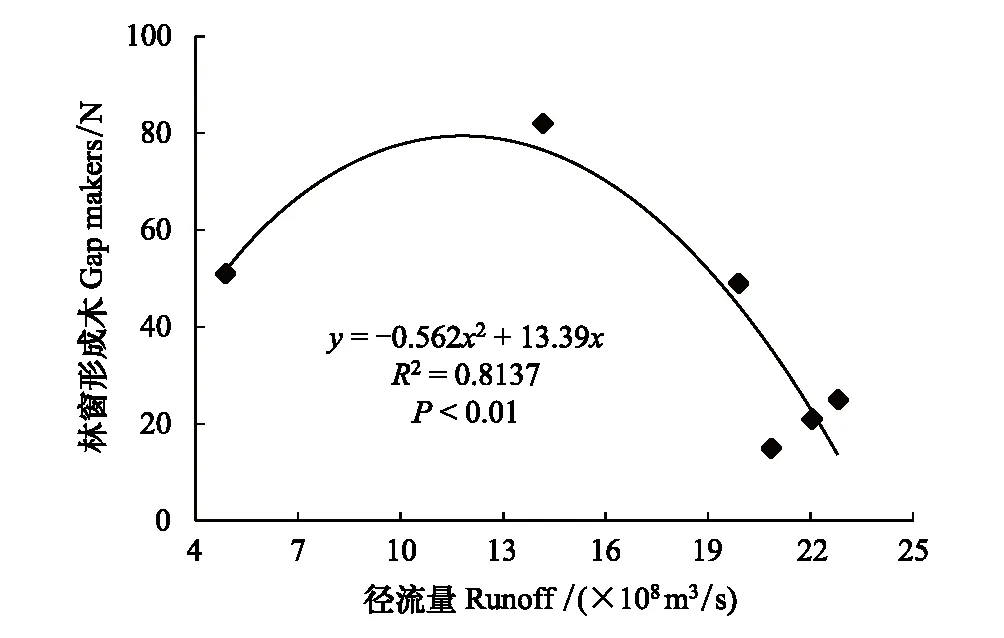
采用种群结构量化分析法,对研究区林窗形成木种群动态进行定量描述。种群结构动态指数(Vpi)可反映种群结构个体数量增长、稳定或衰退的变化趋势。若0 (1) 式中,Sn为第n径级形成木种群个体数,Sn+1为第n+1径级形成木种群个体数,(…)max为选取括号中数列最大值。种群年龄结构的变化动态指数(Vpi)计算公式为: (2) 式中,Sn与Vn式(1)中意义同上,K为林窗形成木种群年龄级数量,Vpi取负、零和正值与Vn一致。在考虑到外部干扰的情况下,经受随机干扰和非随机干扰的林窗形成木径级结构动态指数(Vpi′、Vpi″)的具体计算方法见文献[31]。 3.1.1年径流/耗水量变化 英巴扎水文站位于塔里木河上、中游的交界处,是上游水量下泄到中游的控制点站。其水文站1955至2016年径流/耗水量时序监测资料表明(图2),自20世纪80年代中期以来,塔里木河上游进入中游的年径流量与耗水量大幅度减少,河水无法以漫溢形式补给河道两岸的胡杨林。时间序列与年径流、耗水量的回归方程分别为y径流量=-1.892x+ 24.912(R2=0.785,P<0.01)和y耗水量=-0.165x+ 346.730(R2=0.524,P<0.01)。由回归曲线表明,中游年径流量与耗水量先呈直线下滑趋势,2010年后保持稳定状态,且耗水量趋势线先位于径流量线下方,然后逐渐合并。这说明向塔里木河下游人工调水后中游补给地下水的水量开始减少,从而引起塔里木河中游地区生态环境的恶化势头。 3.1.2地下水埋深变化 获取塔里木河流域管理局在研究区按水分梯度布设的49眼地下水监测井数据。由1990至2016年塔里木河中游地下水埋深变化情况可知,1990至2003年地下水埋深保持在3 m左右;2004年开始,地下水埋深均增深至 4 m以上,并随时间推移继续加深(图3)。由回归模型表明,塔里木河中游地下水埋深随着年度的变化先呈线性增加趋势,2010年后保持在 5 m左右(R2= 0.862,P<0.01),与年径流量与耗水量变化情况基本一致。这是由于塔里木盆地在40—50年前曾受到过大规模开垦的干扰,耕地面积及灌溉用水量骤然增加所引起的[32-33]。 图2 塔里木河中游年径流/耗水量变化Fig.2 Annual runoff/water consumption changes in the middle reaches of the Tarim River 3.2.1不同年度林窗干扰状况 本文采用腐烂等级代替林窗发生时间[28],将林窗发生时间划分为9个等级(表1)。由60个林窗的调查结果可知,塔里木河中游河岸林林窗形成木种群组成简单,仅有胡杨、灰杨两种乔木。图4得知,本次调查的林窗最大年龄约50—60 a,最小年龄约6—10 a,说明塔里木河流域荒漠河岸林的林窗是在近50 a内形成的。各样地内以1997—2006年形成的林窗数量最多(17个),其次是2007—2011年(14个);1957—1966年形成的林窗最少,仅有2个。在荒漠河岸林中,林窗形成木在60—70 a左右逐渐分解完毕,此年龄阶段的倒木残骸难以辨认。因此,森林中没有60 a以上的林窗存在。塔里木河流域经过50多年大规模的水土开发和人为活动的影响,水环境发生巨大变化,从而引起该地区地表水骤减、地下水资源日益匮乏[34],致使胡杨死亡而形成的林窗现象频繁出现。 图3 塔里木河中游地下水埋深变化Fig.3 Ground water depth changes in the middle reaches of the Tarim River 图4 塔里木河中游林窗数量年际变化Fig.4 Change of canopy gap numbers in the middle reaches of the Tarim River 3.2.2林窗形成木种群结构特征 塔里木河中游不同年度林窗胡杨形成木种群数量及其径级结构差异明显(表1),反映了不同水分条件对胡杨种群的影响。由图5可知,除了1967—1976年代以外,塔里木河中游各年林窗形成木胡杨Ⅱ级幼龄树大幅度增加,其中,1997—2006年最多。据统计,1967—1976年间林窗形成木Ⅳ级中龄树缺失(株数为0),其他各径级形成木也极度稀缺(总株数为15)。1987—2016年间各径级林窗形成木较为丰富,尤其是2007—2016年间极为明显(表1)。综上所述,20世纪50年代后塔里木河流域受大尺度干扰[35],再加上2000年塔里木河中游堤防修建后,地下水维系的胡杨种群与水资源失去联系,从而其遭遇陆续丧失或稀缺[15],表现为幼龄胡杨难以生存,种群更新能力明显降低的特征。 图5 胡杨林窗形成木种群径级结构Fig.5 DBH structure of Populus euphratica gap makers population 年代Years最小值Min.最大值Max.平均值Mean数目Number方差Variance标准差Standard deviation偏度Skewness峰度Kurtosis变异系数 Variation coefficient P1957—19664.046.513.821129.25011.3701.9563.2280.8250.1471967—19762.925.7 9.215 40.075 6.3301.6372.5120.6910.1001977—19864.236.411.325 66.730 8.1702.1124.0210.7210.1001987—19963.234.310.149 47.6306.9002.1744.4240.6840.0561997—20062.850.0 9.782 49.155 7.0113.21614.0580.7260.0712007—20161.920.9 8.251 19.072 4.3671.041 0.9600.5310.134 3.2.3林窗形成木种群结构动态 林窗形成木径级结构动态指数(Vpi)可反映形成木种群结构个体数量增长、稳定或衰退的变化趋势,而通过观察Vpi′、Vpi″的取值变化,可以分析林窗乔木形成木在经受随机与非随机干扰时的稳定程度。由表2可知,在无外部干扰下,塔里木河中游各年代(1957—1966、1967—1976、1977—1986、1987—1996、1997—2006和2007—2016)的Vpi值分别为:-0.073、-0.060、0.000、0.040、0.200和0.460。在考虑到外部干扰的情况下,经受随机干扰和非随机干扰的形成木径级结构动态指数(Vpi′、Vpi″)分别为:-0.0040、-0.0019、0.0025、0.0071、0.0057、0.0084和-0.0180、-0.0300、0.0600、0.0970、0.0400、0.0554。由此可见,1977—1986、1987—1996、1997—2006与2007—2016年间塔里木河中游胡杨林窗形成木种群结构呈增长趋势,1957—1966与1967—1976年间则为减少。实施生态输水的前期与初期(1997至2004),地下水埋深保持3 m左右,可满足各龄级胡杨生长发育的水分需求。随后,塔里木河中游径流量与耗水量持续减少,导致地下水埋深不断增加,河道两岸生态用水紧缺。林窗形成木结构动态指数增加趋势逐步提高,这是因为胡杨种群应对外界干扰的抵抗能力持续降低(表3)。 表3 林窗形成木种群径级结构动态分析 V1: Ⅰ—Ⅱ径级变化动态指数 Dynamic index value of Ⅰ—Ⅱclass sizes;V2: Ⅱ—Ⅲ径级变化动态指数 Dynamic index value of Ⅱ—Ⅲclass sizes;V3: Ⅲ—Ⅳ径级变化动态指数 Dynamic index value of Ⅲ—Ⅳclass sizes;V4: Ⅳ—Ⅴ径级变化动态指数 Dynamic index value of Ⅳ—Ⅴclass sizes;Vpi: 无外部干扰变化动态指数 Dynamic index value under no external disturbance;Vpi′: 完全随机干扰变化动态指数 Dynamic index value under completely random disturbance;Vpi″: 非随机干扰下变化动态指数 Dynamic index value of non-random disturbance;P: 完全随机干扰下的风险极大值Maximum risk value under completely random disturbance;P′: 非随机干扰下的风险极大值 Maximum risk value under non-random disturbance 图6 塔里木河中游林窗形成木种群结构动态与径流量的关系Fig.6 Relationship between structure dynamic of gap makers population and annual runoff in the middle reaches of the Tarim River 水是影响植物生存、生长发育诸多环境因子中最主要的限制因子之一[36]。地表径流是维系干旱区地下水依赖型胡杨种群生长发育的重要水资源,Vpi′ 表示林窗胡杨形成木种群在随机干扰下的径级结构动态。如图6所示,Vpi′与地表径流量具有显著性负相关,其回归方程为y= 0.0005x- 0.0146 (R2= 0.8405,P<0.01)。该模型显示,随径流量的增加,林窗形成木结构动态指数不断下降。当径流量降至25.86×108m3时,林窗形成木增长指数变为正值,林窗形成木种群出现增长型现象。这表明地表径流量的减少是导致胡杨种群死亡的主要原因。尤其是,地下水与其重要补给来源关系被切断,使地下水埋深逐渐加深[37],林分自然更新受制,严重阻碍了胡杨林的正常生长发育。 由图7可见,林窗数量与年径流量之间存在较好的拟合负相关,其拟合方程为y=-0.0728 x2+1.6458 x + 6.1206(R2=0.624,P<0.01)。拟合曲线表明,林窗数量随着年径流量的增加而不断下滑趋势,径流量越多,林窗数量就越少。径流量在6.53×108—17.81×108m3范围内林窗数量最多,说明在其范围的径流量很有可能对林窗的形成有着显著的影响。相对其而言,林窗形成木数量与流域的年径流量之间存在显著性负相关。如图8所示,林窗形成木数量随着年径流量的增加而大幅度减少趋势,其拟合方程为y=-0.562x2+13.39x(R2=0.8137,P< 0.01)。径流量越多林窗内死亡树木就越少,与林窗数量随着年径流量的变化趋势基本一致。这可能是由于地表水与地下水有直接的关系,间接影响到胡杨的生存生长,从而导致干旱区林窗现象的频繁出现。 图7 林窗数量与径流量关系Fig.7 Relationship between the number of forest gap and annual runoff 图8 林窗数量与径流量关系Fig.8 Relationship between the number of forest gap and annual runoff 从年际变化来看,塔里木河中游年径流量和耗水量均呈直线下滑趋势,同时地下水埋深不断加深。徐梦辰等学者借助稳定同位素技术分析水文补给关系,证明了塔里木河中游地下水主要依靠地表径流水来补给[15]。2000年的堤防修建和人工调水阻碍了洪水漫溢[38],地下水失去其补充水源,河道两岸不具备植物生存的环境条件,引起无法满足个体对水分因子的需求而种内竞争与排斥加剧。至于大龄树(或老龄树)阶段,个体对环境的要求高,尤其是对水资源的需求。然而,河道两岸地下水位不断降低,个体以积累大量资源为维持生存,对水分、光照、空间及营养的争夺达到顶峰,使个体面临死亡,从而导致林窗现象的频繁出现。 干旱区降水稀少,几乎没有有效降水供给胡杨生存[39-40]。洪水漫溢消失后,胡杨的生长发育主要依靠河道径流侧渗与生态输水补给。由林窗胡杨形成木种群结构动态指数与地表径流量的回归模型显示,当径流量降至25.86×108m3时,形成木种群转变为增长趋势。在塔里木河中游地表水逐年减少的影响下,河道两岸地下水位不断降低,胡杨生长所需的生态用水日益匮乏,直接制约了胡杨种群的生存。随地表径流量的降低,林窗形成木种群结构动态指数不断增加,除丰水期(1957—1966与1967—1986年代)呈衰退型以外,其余年段(1987—1996、1997—2006和2007—2016)均表现为增长型特征,这种动态指数反映了胡杨林林窗形成木种群对水文变化的响应机制。为应对干旱区环境恶化,荒漠河岸胡杨林林窗形成木在种群结构上做出了生态响应。地下水位的逐年降低使林窗形成木种群结构动态指数不断增加并转入增长趋势,形成木种群具有幼化且长期增长的特征。在不同水文影响下,胡杨林已出现了长期的林窗形成木种群增加现象。由拟合模型显示,林窗及其形成木随径流量的减少而大幅度增加趋势。径流量在6.53×108—17.81×108m3范围内林窗形成木数量将持续增长,将面临一次高峰死亡。由此可知,由于种群结构失调、胡杨林生存受限,若不采取适当的保育措施,胡杨种群将逐渐退化。 1)塔里木河中游地下水只能依靠地表径流的侧渗与生态输水补给,这不仅从影响范围还是补充水量上都无法取代来自洪水的自然漫溢效果,与此同时,受年径流量、耗水量逐年减少的影响,地下水补给更加受损。塔里木河中游水文条件的变化导致该地区水资源日益匮乏,河道两岸生态用水骤减,从而导致该区域干旱、盐碱胁迫加剧。 2)1970年前的外界干扰还没影响到水文过程,由于1970年后的人类干扰对环境变化具有迟滞性特征,1984年开始地表径流量大幅度减少,其滞后效应使地下水埋深2003年后持续增加,林窗胡杨形成木种群变为增长型,这验证了林窗形成木对水文变化的响应。 3)地下水作为维系胡杨林生存繁衍的基础,它主要依靠地表水的补给。从2000年起向塔里木河下游实施人工生态输水工程后,虽然下游“绿色走廊”得以一定程度的恢复,生态环境日渐好转,但对中游河段的水文过程带来了一定的压力乃至产生了负面影响。随着来自洪水漫溢的地表径流量日益匮乏,河道两岸生态用水逐渐紧缺,地下水位逐年下降,土壤表层干旱、盐碱胁迫加剧。因此,以中游河段牺牲生态环境来换取下游生态修复方法的利弊有待权衡。3 结果与分析
3.1 塔里木河中游水文特征
3.2 胡杨林林窗特征
3.3 林窗形成木结构动态与年径流量的关系
3.4 林窗及其形成木数量与径流量的关系
4 讨论
5 结论
猜你喜欢
广西林业科学(2022年4期)2022-11-07
公民与法治(2022年10期)2022-10-12
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
意林绘阅读(2019年2期)2019-03-05
下一代英才(2018年12期)2018-02-20
天文爱好者(2015年12期)2015-08-23
中南林业科技大学学报(2015年8期)2015-05-22
新西部(2014年10期)2015-01-05
吐鲁番(2014年2期)2014-02-28
