
基于iPBS标记的玫瑰香系葡萄遗传多样性分析与指纹图谱构建
2019-10-28 05:31张安世庞金城高登涛魏志峰
浙江农业学报 2019年10期
张安世,庞金城,刘 莹,高登涛,魏志峰,*
(1.焦作师范高等专科学校 理工学院,河南 焦作454000; 2.中国农业科学院 郑州果树研究所,河南 郑州 450009)
葡萄(VitisviniferaL.)是栽培最早、分布最广的果树之一,全球葡萄共有70多个种、约14 000个品种[1]。其中,玫瑰香系是葡萄品种的一个重要系列。玫瑰香系葡萄是由国内外育种工作者利用玫瑰香葡萄选育的系列品种,据统计,目前大约有76个品种。我国育种工作者用玫瑰香作亲本共育成了56个葡萄品种,已成为我国葡萄育种中的骨干亲本[2]。目前,全球的葡萄业产值已位居世界农产品第5位[3]。葡萄也是我国发展速度最快的果树种类之一,玫瑰香系葡萄更是以其特有的玫瑰香味且果粒大等特点深受广大消费者喜欢和育种工作者的偏爱。随着玫瑰香系葡萄新品种的不断选育,品种系谱也不断延伸,加上地区间的品种交流日益频繁,“同名异物”或“同物异名”的现象时有发生,因此,需要理想的鉴定技术快速、准确地区分表型极为相似的品种。
传统的形态学特征、同工酶谱等手段容易受环境条件和组织发育阶段的影响,在品种鉴定中有很大的局限性。由于DNA分子标记不受环境因子和时空条件影响,具有高效、快捷等优点,可直接反映品种间的遗传差异,已成为品种鉴定的有效方法之一。Inter-primer binding site (iPBS)是Kalendar等[4]提出的以LTR类反转录转座子保守位点设计引物进行扩增的一种分子标记技术,无需预先获知相关的LTR序列,比常规的分子标记技术更有优势[5]。目前,简单重复序列间扩增(ISSR)、扩增片段长度多态性(AFLP)、简单重复序列标记(SSR)等分子标记技术在葡萄中均有成功应用。研究表明,iPBS的多态性高于ISSR[6],比AFLP更易操作[7],比SSR成本更低,已逐渐应用于种质资源的遗传多样性分析和品种鉴定[8-9],在葡萄中也得到了初步应用[10-11]。为进一步了解玫瑰香系葡萄的遗传关系,本研究利用iPBS标记并结合已有系谱[2,12],对39个玫瑰香系葡萄品种进行多样性分析,建立DNA指纹图谱,为葡萄种质资源的鉴定、品种的保护利用等提供科学依据。
1 材料与方法
1.1 材料
供试的39个玫瑰香系葡萄种质材料由中国农业科学院郑州果树研究所葡萄种质资源圃提供(表1)。每个品种选取3株,每株采集2枚健康的幼叶,分别用封闭的塑料袋包装,保存于冰盒,及时送回实验室,液氮速冻,置于-80 ℃超低温冰箱中保存、备用。
1.2 葡萄基因组DNA的提取
每份材料取3枚不同植株的叶片等量混合,采用改良CTAB法[13]提取葡萄基因组DNA,用TE缓冲液将模板DNA浓度稀释至20 ng·μL-1,-20 ℃保存备用。
1.3 iPBS-PCR扩增
选用Kalendar等[4]发表的83个iPBS引物对供试材料进行扩增。反应体积10 μL,包含1.0 μL DNA、1.0 μL引物、5.0 μL 2×TaqMasterMix和3.0 μL RNase-Free water。iPBS-PCR扩增程序为:95 ℃ 5 min;95 ℃ 15 s,50 ℃ 1 min,68 ℃ 1 min,35个循环;72℃ 5 min,4 ℃保存。电泳采用1.5%琼脂糖凝胶分离扩增产物。
1.4 数据统计与分析
统计分析电泳图谱上清晰可辨的条带,在电泳迁移率的相同位置上,用“1”或“0”分别代表条带的“有”或“无”,构建1/0矩阵。利用POPGENE1.32软件计算相关遗传多样性参数[多态性百分率(PPL)、观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性(H)和Shannon’s多样性指数(I)]。各引物的多态性信息含量(PIC)参照黄秀等[14]的方法计算,DNA指纹图谱参考张林等[15]的方法绘制,用NTSYS-pc 2.0软件构建UPGMA聚类图。
2 结果与分析
2.1 iPBS引物筛选与多态性分析
利用83个iPBS引物对39个玫瑰香系葡萄品种进行了PCR扩增,筛选出了14条多态性高、重复性好、条带清晰的引物进行统计分析,结果显示(表2):14个引物共扩增出152个条带,其中132条带具有多态性,多态性比率为86.86%,平均每个引物扩增的位点总数为10.86个。引物2076的扩增图谱见图1。各引物的PIC的变化范围为0.796 5~0.914 9,平均值为0.863 8,说明所选引物在39个玫瑰香系葡萄品种间具有较高的多态性。观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)和Shannon’s信息指数(I)的平均值分别为1.868 4、1.409 0、0.251 8和0.390 7,也处于较高水平。利用SPSS 17.0软件对PIC、Ne、H和I进行了非参数Kruskal Wallis Test独立样本检验,结果表明:39个玫瑰香系葡萄种质间遗传多样性水平存在显著性差异(X2=49.945,P﹤0.00 1)。
表1 供试材料
Table1Grape materials used in study

序号No.品种名称Cultivarnames品种类型Cultivartype亲本来源Directparents1郑州早红Zhengzhouzaohong欧亚种V.vinifera玫瑰香×莎巴珍珠MuscatHamburgh×PearlofCsaba2蓓蕾Beilei欧美杂交种Euro-Americanhybrids不详Unknown3蓓蕾玫瑰Beileimeigui欧美杂交种Euro-Americanhybrids玫瑰香×蓓蕾MuscatHamburgh×Beilei4脆红Cuihong欧美杂交种Euro-Americanhybrids玫瑰香×白香蕉MuscatHamburgh×Baiixiangjiao5贵妃玫瑰Guifeimeigui欧美杂交种Euro-Americanhybrids红香蕉×葡萄园皇后Hongxiangjiao×Putaoyuanhuanghou6黑香蕉Heixiangjiao欧美杂交种Euro-Americanhybrids红香蕉×葡萄园皇后Hongxiangjiao×Putaoyuanhuanghou7红香蕉Hongxiangjiao欧美杂交种Euro-Americanhybrids玫瑰香×白香蕉MuscatHamburgh×Baiixiangjiao8红双味Hongshuangwei欧美杂交种Euro-Americanhybrids红香蕉×葡萄园皇后♀Hongxiangjiao×Putaoyuanhuanghou♀9红莲子Honglianzi欧亚种V.vinifera玫瑰香×葡萄园皇后MuscatHamburgh×Putaoyuanhuanghou10巨玫瑰Jumeigui欧美杂交种Euro-Americanhybrids沈阳玫瑰×巨丰Shenyangmeigui×Kyoho11康太Kangtai欧美杂交种Euro-Americanhybrids康拜尔早生Kangbaierzaosheng12康拜尔早生Kangbaierzaosheng欧美杂交种Euro-Americanhybrids早熟摩阿×(贝尔维台×玫瑰香)Zaoshumoe×(Beierweitai×MuscatHamburgh)13山东大紫Shandongdazi欧亚种V.vinifera山东早红Shandongzaohong14天使玫瑰香TTianshimeiguixiang欧美杂交种Euro-Americanhybrids不详Unknown15香悦Xiangyue欧美杂交种Euro-Americanhybrids沈阳玫瑰×8001Shenyangmeigui×800116紫珍香Zizhenxiang欧美杂交种Euro-Americanhybrids沈阳玫瑰×8001Shenyangmeigui×80011787-1欧亚种V.vinifera玫瑰香MuscatHamburgh18京秀Jingxiu欧亚种V.vinifera玫瑰香×红无籽露MuscatHamburgh×Hongwuzilu19卡拉斯玫瑰Kalasimeigui欧亚种V.vinifera玫瑰香×卡拉斯♀20玫瑰香MuscatHamburgh欧亚种V.vinifera黑汉×亚历山大BlauerTrollinger×MuscatofAlexandria21沈阳玫瑰Shenyangmeigui欧美杂交种Euro-Americanhybrids玫瑰香MuscatHamburgh22胜利Shengli欧亚种V.vinifera玫瑰香×巴尔坎斯基MuscatHamburgh×Baerkansiji23香妃Xiangfei欧亚种V.vinifera(玫瑰香×莎巴珍珠)×绯红(MuscatHamburgh×PearlofCsaba)×Feihong24意大利Yidali欧亚种V.vinifera玫瑰香×比坎♀MuscatHamburgh×Bikan25早玛瑙Zaomanao欧亚种V.vinifera玫瑰香×京早晶MuscatHamburgh×Jingzaojing26泽玉Zeyu欧亚种V.vinifera玫瑰香×龙眼MuscatHamburgh×Longyan27泽香Zexiang欧亚种V.vinifera玫瑰香×龙眼MuscatHamburgh×Longyan28早熟玫瑰香Zaoshumeiguixiang欧亚种V.vinifera玫瑰香×莎巴珍珠MuscatHamburgh×PearlofCsaba29早玫瑰Zaomeigui欧亚种V.vinifera玫瑰香×莎巴珍珠MuscatHamburgh×PearlofCsaba308612欧美杂交种Euro-Americanhybrids郑州早红×巨峰Zhengzhouzaohong×Kyoho31爱神玫瑰Aishenmeigui欧亚种V.vinifera玫瑰香×京早晶MuscatHamburgh×Jingzaojing32博多白Boduobai欧亚种V.vinifera意大利×罗扎基Yidali×Luozhaji33斯帕汉堡Sipahanbao欧美杂交种Euro-Americanhybrids玫瑰香×康拜尔早生MuscatHamburgh×Kangbaierzaosheng34大粒玫瑰香Dalimeiguixiang欧亚种V.vinifera玫瑰香Meiguixiang35红茧Hongjian欧亚种V.vinifera红意大利×龙尼坤♀Hongyidali×Longnikun36红高Honggao欧亚种V.vinifera意大利Yidali37红旗特早玫瑰Hongqitezaomeigui欧亚种V.vinifera玫瑰香Meiguixiang38红意大利Hongyidali欧亚种V.vinifera意大利Yidali39甲斐乙女Jiafeiyinv欧亚种V.vinifera蓓蕾玫瑰×甲斐路Beileimeigui×Jiafeilu
表2iPBS引物扩增结果与多态性信息
Table2Results and polymorphism information of iPBS primers
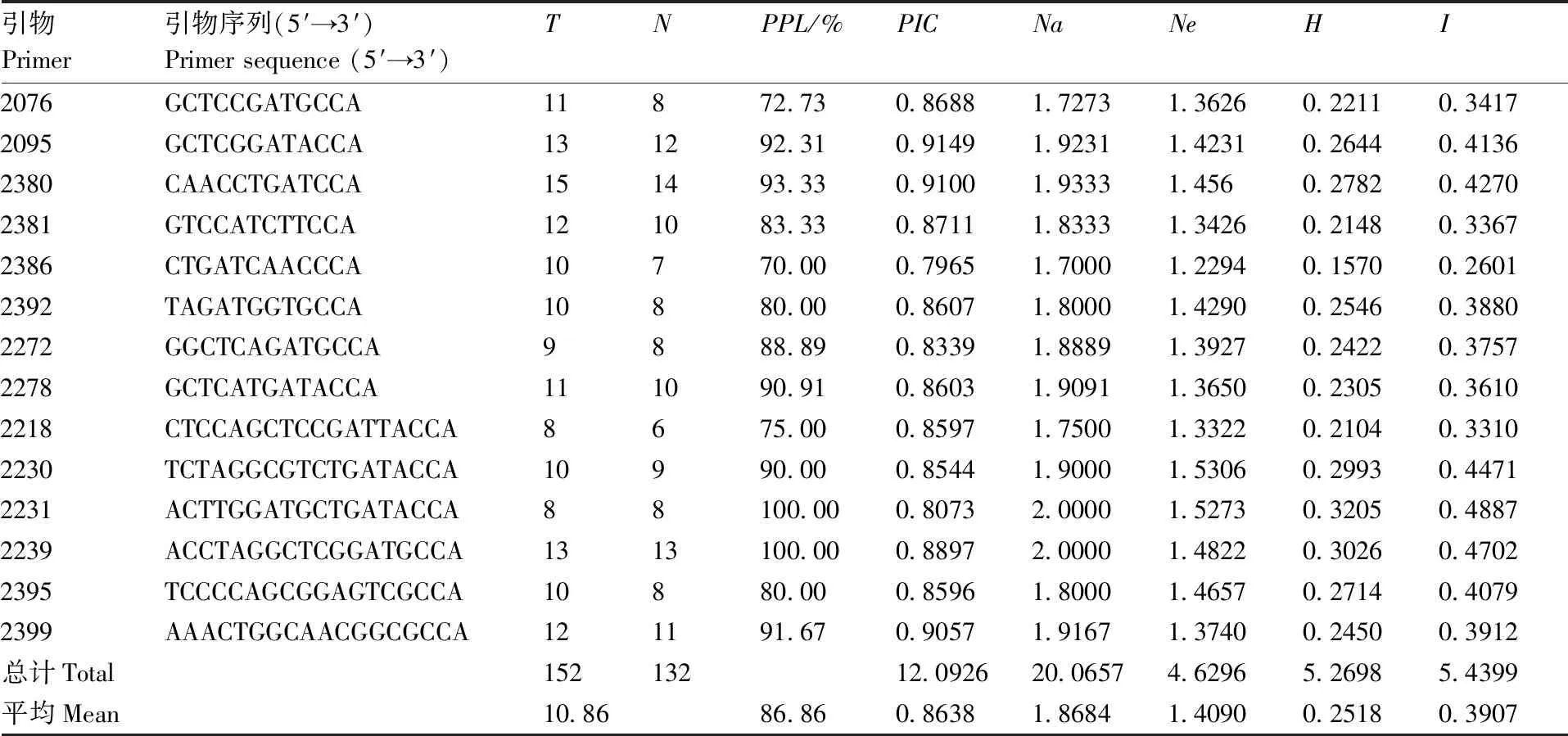
引物Primer引物序列(5′→3′)Primersequence(5′→3′)TNPPL/%PICNaNeHI2076GCTCCGATGCCA11872.730.86881.72731.36260.22110.34172095GCTCGGATACCA131292.310.91491.92311.42310.26440.41362380CAACCTGATCCA151493.330.91001.93331.4560.27820.42702381GTCCATCTTCCA121083.330.87111.83331.34260.21480.33672386CTGATCAACCCA10770.000.79651.70001.22940.15700.26012392TAGATGGTGCCA10880.000.86071.80001.42900.25460.38802272GGCTCAGATGCCA9888.890.83391.88891.39270.24220.37572278GCTCATGATACCA111090.910.86031.90911.36500.23050.36102218CTCCAGCTCCGATTACCA8675.000.85971.75001.33220.21040.33102230TCTAGGCGTCTGATACCA10990.000.85441.90001.53060.29930.44712231ACTTGGATGCTGATACCA88100.000.80732.00001.52730.32050.48872239ACCTAGGCTCGGATGCCA1313100.000.88972.00001.48220.30260.47022395TCCCCAGCGGAGTCGCCA10880.000.85961.80001.46570.27140.40792399AAACTGGCAACGGCGCCA121191.670.90571.91671.37400.24500.3912总计Total15213212.092620.06574.62965.26985.4399平均Mean10.8686.860.86381.86841.40900.25180.3907
T,位点总数;N,多态性位点数;PPL,多态性位点百分率;PIC,多态性信息含量;Na,观测等位基因数;Ne,有效等位基因数;H,Nei’s基因多样性;I,Shannon信息指数。
T, Total number of loci;N, Number of polymorphic loci;PPL, Percentage of polymorphic loci;PIC, Polymorphism information content;Na, Observed number of alleles;Ne, Effective number of alleles;H, Nei’s gene diversity;I, Shannon information index.
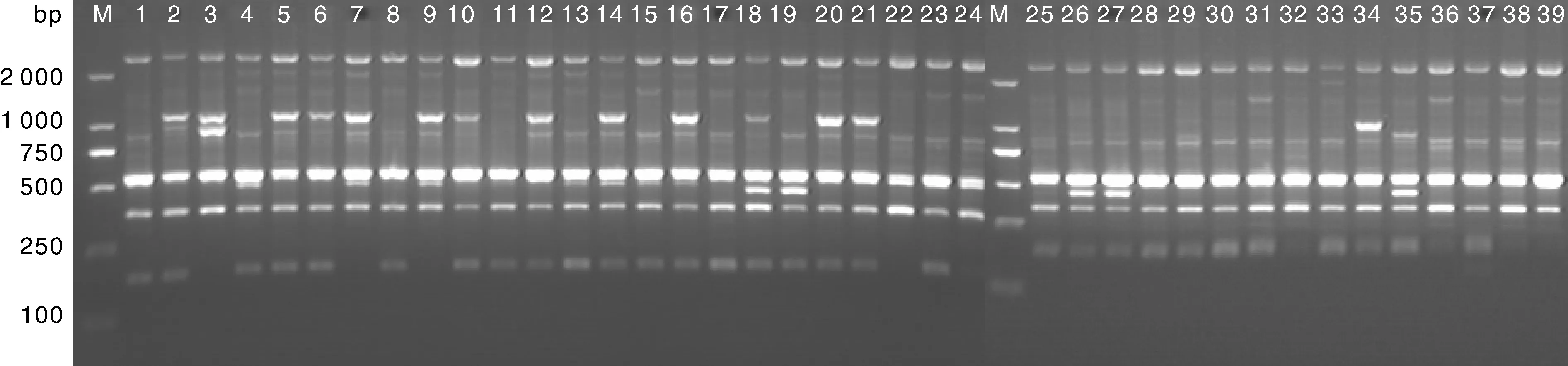
M,标准分子量;1~39,材料编号同表1。M, DL 2 000 standard marker; No.1-39 were the same as table 1.
2.2 玫瑰香系葡萄种质资源的聚类分析
利用NTSYS-pc 2.0软件构建了39个玫瑰香系葡萄品种的聚类图(图2),遗传相似性系数(GS)为0.526 3~0.940 8,变幅达0.413 5,说明39个玫瑰香系葡萄品种间存在较大的遗传差异。在GS为0.72处,可将39个玫瑰香系葡萄品种为4组。第Ⅰ组包括26个品种,分别为郑州早玉、早玫瑰、87-1、脆红、玫瑰香、沈阳玫瑰、大粒玫瑰香、胜利、天使玫瑰香、香悦、紫珍香、红茧、香妃、意大利、红高、红意大利、博多白、甲斐乙女、早熟玫瑰香、卡拉斯玫瑰、早玛瑙、泽玉、泽香、爱神玫瑰、红旗特早玫瑰和京秀;第Ⅱ组包括6个品种,分别为蓓蕾、贵妃玫瑰、黑香蕉、红双味、红香蕉和红莲子;第Ⅲ组包括6个品种,分别为巨玫瑰、康拜尔早生、康太、山东大紫、8612和斯帕汉堡;第Ⅳ组仅有蓓蕾玫瑰1个品种。其中,第Ⅰ组有5个欧美杂交种,21个欧亚种,第Ⅱ组有5个欧美杂交种和1个欧亚种,第Ⅲ组有5个欧美杂交种和1个欧亚种,第Ⅳ组蓓蕾玫瑰为欧美杂交种。从上述结果可知,欧亚种主要分布Ⅰ组,而欧美杂交种主要分布在第Ⅱ、Ⅲ和Ⅳ组。如果再将第Ⅰ组细分,可以分为5个小组。其中,第1组分布5个欧美杂交种(脆红、沈阳玫瑰、天使玫瑰香、香悦和紫珍香)和6个欧亚种(郑州早玉、早玫瑰、87-1、玫瑰香、大粒玫瑰香和胜利),其余4个小组全为欧亚种,共计15个品种。
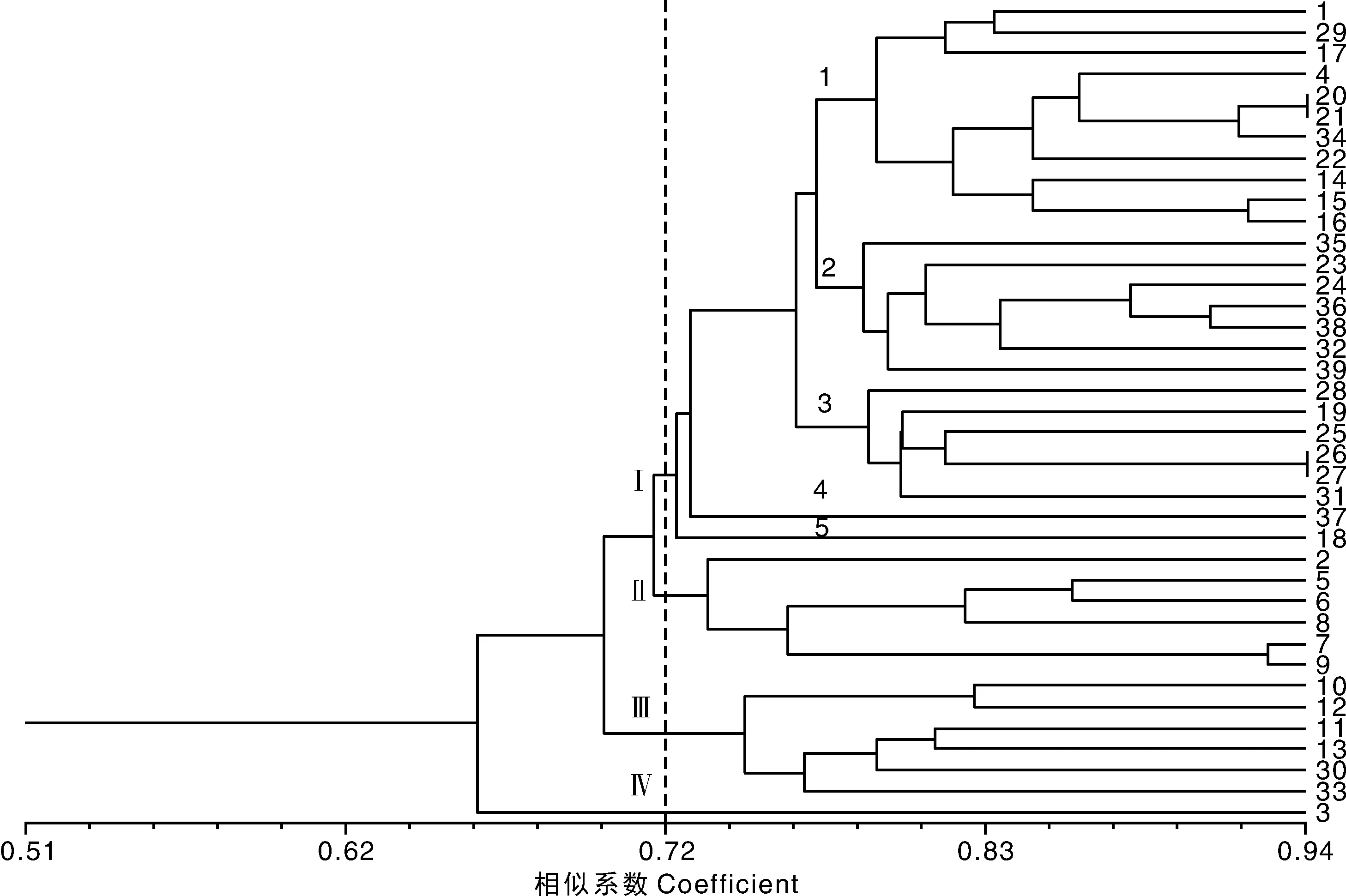
1~39材料编号同表1。No. 1-39 were the same as table 1.
2.3 DNA指纹图谱的构建
根据筛选的14个引物对玫瑰香系葡萄的扩增结果,选取2076、2095、2380、2392、2272、2278、2218和2231共8个引物扩增的16个多态性位点,构建了39个玫瑰香系葡萄品种的DNA指纹图谱(图3)。其中,部分品种较难区分,它们之间仅1个位点的差异。如康太与8612[(2272)-1288 bp]、玫瑰香与大粒玫瑰香[(2231)-873 bp]、意大利与博多白[(2231)-873 bp]、泽玉与泽香[(2218)-1296bp]。此图谱可以有效区分39个玫瑰香系葡萄品种。

1-39材料编号同表1。No. 1-39 were the same as table 1。
3 讨论
3.1 玫瑰香系葡萄品种的遗传多样性分析
通过iPBS标记分析可知:39个玫瑰香系葡萄品种的多态性位点百分率(PPL)为86.86%,高于丁洁菲等[16]74%的检测结果,与李琳等[17]、王发明等[18]的研究结果类似,但低于李雪雁等[19]和郭印山等[20]的100%的检测结果。这种现象可能是由于不同研究者采用的分子标记类型不同、研究对象的种类和数量各异等因素所致。同时,iPBS分析得到的PIC平均值较高(0.863 8),说明本研究筛选的引物具有很高的多态性。另外,从评价遗传多样性水平的其他4个主要参数Na、Ne、H和I来看,其平均值分别为1.868 4、1.409 0、0.251 8和0.390 7,也处于较高水平。品种间的遗传相似系数(GS)为0.526 3~0.940 8,变幅(0.413 5)较大。上述结果说明,39个玫瑰香系葡萄品种间具有较为丰富的遗传变异,同时也说明,在玫瑰香系葡萄不断的人工定向杂交选育过程中其遗传多样性仍维持在较高水平。
3.2 玫瑰香系葡萄品种的聚类分析
本研究中的39个玫瑰香系葡萄品种,共涉及欧美杂交种(16个)和欧亚种(23个)2个类群。聚类分析结果表明:在遗传相似系数(GS)为0.72处可将39个玫瑰香系葡萄品种分为4组。其中欧亚种群主要分布在第Ⅰ组,而欧美杂交种群主要分布在第Ⅱ、Ⅲ和Ⅳ组。在第Ⅰ组中除第1小组出现欧美杂交种和欧亚种互插外,其他4个小组全为欧亚种,分布相对集中。因此,从整体来看,2个类群的玫瑰香系葡萄分布呈现一定规律。将此聚类结果与系谱[12]对比可以发现,两者基本吻合。例如玫瑰香与莎巴珍珠的杂交后代郑州早红与早玫瑰聚在了一起;沈阳玫瑰香和大粒玫瑰香是通过玫瑰香芽变选育的品种,三者具有很大的相似性,聚在一起;紫珍香和香悦是沈阳玫瑰香与8001杂交选育的后代,两者聚在一起,且与玫瑰香、沈阳玫瑰香和大粒玫瑰香一起聚在第Ⅰ组第1小组的一个分支;第Ⅰ组第2小组分布7个玫瑰香系葡萄品种,除香妃和甲斐乙女外,其余5个品种(红茧、意大利、红高、红意大利和博多白)是意大利和通过意大利选育的品种,显示出很近的亲缘关系;另外,黑香蕉、贵妃玫瑰和红双味是通过红香蕉选育的后代,这4个品种聚在一起;泽玉和泽香是玫瑰香和龙眼的杂交后代,它们也聚在了一起。因此,该聚类分析能较好地反映玫瑰香系葡萄品种间的遗传关系。说明iPBS分子标记技术在葡萄的遗传多样性研究中是可行的。但是,该聚类结果也出现了与系谱不一致的现象。如爱神玫瑰和早玛瑙是玫瑰香与京早晶的杂交后代,两者未能聚在一起;早玫瑰香、郑州早红和早玫瑰具有相同的亲本却未能聚在一起。另外,从系谱分析,蓓蕾、蓓蕾玫瑰和甲斐乙女应该具有较近的亲缘关系,但聚类分析未能有效显示。出现这种现象可能的原因,一是引物数量较少,检测位点不足;另一方面,iPBS标记自身也有一定的局限性,任何一种分子标记都无法覆盖全基因组序列。因此,需要结合形态学和其他分子标记技术进行深入研究,为玫瑰香系葡萄种质遗传多样性与品种的鉴定提供更多的分子依据。
3.3 iPBS标记在种质资源鉴定上的应用
依据植物的形态学特征很难区分表型特征极为相似的品种,而DNA分子标记可直接反映品种间的遗传差异,已成为品种鉴定的有效方法之一。iPBS标记在植物品种鉴定方面已有成功应用。石艳艳[10]利用15个iPBS引物对巨峰及其芽变品种峰早和京亚及其芽变品种洛浦早生的顶端分生组织细胞原层(LⅡ层)、LⅠ+ LⅡ层进行了比较分析,结果显示,上述2个早熟芽变品种的LⅡ层发生了变化,为亲本及其芽变品种的鉴定提供了一种新的方法。王文婷[21]利用8个iPBS引物产生的特异性条带对12个杨梅品种进行了有效鉴定。李婵等[22]利用11个扩增效果较好的iPBS引物,对47份菌草种质材料进行了有效区分。本研究通过iPBS技术,利用8个引物扩增的16个多态性高、重复性好的特异性位点构建了39个玫瑰香系葡萄品种的DNA指纹图谱,为后续葡萄种质间的大规模鉴定与评价提供了理论支撑。
猜你喜欢
华人时刊(2022年11期)2022-09-15
今日农业(2021年14期)2021-11-25
小哥白尼(军事科学)(2021年8期)2021-11-22
桉树科技(2020年2期)2020-07-16
四川蚕业(2020年2期)2020-07-10
英美文学研究论丛(2019年2期)2019-08-24
科学导报(2018年47期)2018-05-14
海外星云(2016年17期)2016-12-01
华人时刊(2016年17期)2016-04-05
航运交易公报(2015年40期)2015-11-30
