
水稻磷脂酶PLD家族生物信息学分析
2019-11-14 08:30康伟伟易自力陈智勇谭炎宁
西南农业学报 2019年10期
康伟伟,张 超,2,易自力,陈智勇,谭炎宁
(1. 湖南农业大学生物科学技术学院,湖南 长沙 410128;2. 湖南杂交水稻研究中心杂交水稻国家重点实验室,湖南 长沙 410125)
【研究意义】磷脂酶广泛存在于动植物中,具有磷脂酶A1、A2、B、C和D等类型,它们能特异地作用于磷脂分子的各个酯键,将其水解为脂肪酸及亲脂性物质[1]。植物磷脂酶D(Phospholipase D,PLD)即磷脂酰胆碱磷脂水解酶(EC3. 1. 4. 4),其作用底物较为宽广(如底物有磷脂酰胆碱、磷脂酰丝氨酸等),能专一性水解磷脂中的4位酯键生成磷脂和有机碱[2],在脂质代谢、生物膜形成、信号转导以及逆境响应方面发挥着重要作用[3]。【前人研究进展】PLD作为植物磷脂酶家族的重要组成亚类,其结构和功能越来越受到关注。拟南芥基因组中共存在12个PLD成员,根据其序列相似性及蛋白结构域的不同被分成α、β、δ、γ和ζ等5种不同的亚型[4]。水稻PLD含有17个成员,预测其中16个为拟南芥PLD的同源蛋白,剩余一个在水稻中未被编码;水稻PLD比拟南芥增加了φ和κ两种亚型[5]。有趣的是,φ型PLD并不具有C2结构域或PX/PH结构域而含有一段信号肽,因此也被称为信号肽PLD(SP-PLD)[4-5];φ型PLD已在葡萄、杨树等多种植物中相继被发现[6]。目前,多项研究发现拟南芥PLD参与干旱、盐和低温等胁迫应答。过表达AtPLDα1基因提高了植株对脱落酸ABA的敏感性,促进气孔关闭,减少水分蒸腾散失,从而增强了植株的抗旱能力,而敲除株系则表现出相反的性状[7-8]。在盐胁迫条件下,AtPLDα1和AtPLDδ基因被激活;AtPLDα1的突变株出现了生长缺陷,AtPLDα1、AtPLDδ的单突变体及其双突变体对盐敏感性增加[11,14];类似地,番茄LePLDα1在盐胁迫下被诱导表达,其突变细胞在盐胁迫下活性显著降低[11,15];拟南芥AtPLDα3也被报道参与了盐、脱水和ABA的响应[10]。AtPLDδ与低温响应有关,AtPLDδ突变株在-7 ℃或更低温度条件下其存活能力明显低于野生型[11,16]。【本研究切入点】在水稻中,OsPLDζ1受盐诱导表达,OsPLDζ1的突变株在盐胁迫条件下生长发育明显缓慢,植株矮化且存活率下降[17];OsPLDβ1通过激活ABA信号通路,抑制种子的萌发[18]。与之相反,OsPLDα3突变株在盐胁迫条件下比野生型呈现出更好的生长状况,表明OsPLDα3在盐胁迫响应中具有负调控作用[17]。但是目前,水稻PLD家族基因功能的研究相对较少。【拟解决的关键问题】为了加强对该基因家族的深入认识,本研究利用生物信息学手段分析了水稻16个磷脂酶PLD家族成员的基因结构、蛋白域基序、时空表达以及激素诱导表达模式,旨在为进一步研究该家族的生物学功能提供参考。
1 材料与方法
1.1 水稻PLD家族基因结构分析方法
从Amarjeet Singh(2012)文献中获得水稻PLD家族基因的登录号,通过在RGAP网站上搜寻并下载其基因组序列和CDS序列,然后在GSDS(Gene Structure Display Server 2.0)网站分析基因结构。
1.2 水稻PLD家族蛋白基序分析方法
在Ensembl Plants网站上搜寻并下载水稻OsPLD家族蛋白序列,利用在线分析网站MEME(http://meme-suite.org/)分析基序种类与特征。
1.3 水稻PLD家族时空表达与激素诱导表达分析方法
从RiceXPro网站所公布4X44K Microarray芯片数据中提取OsPLD家族基因在叶、叶鞘、茎、根、花药、子房等不同组织部位及营养时期、生殖时期和成熟期等不同发育时期的表达数据,进行标准化处理;此外,利用该数据库调取以种子萌发至生长7 d的水稻嫩芽为样本的激素(如脱落酸、赤霉素、生长素、油菜素内酯、细胞分裂素和茉莉酸)处理表达数据,经归一标准化处理后使用MeV4.6作图,得到PLD家族的时空表达谱热图及激素诱导表达谱。
2 结果与分析
2.1 水稻PLD家族基因结构分析
经GSDS分析发现,水稻PLD家族成员基因长度在2982 bp(OsPLDα5)~11 305 bp(OsPLDζ1)范围内(图1),绝大多数基因长度大于4 kb,只有OsPLDα3、OsPLDα5和OsPLDα8等少数基因长度少于4 kb,它们的长度分别为3671、3793和2982 bp(图1)。不同亚类基因可能具有不同的外显子数目,但同亚类基因的外显子数大致一致;α型外显子数目最少,OsPLDα1、OsPLDα2、OsPLDα3、OsPLDα4、OsPLDα5和OsPLDα8的外显子数量均只有3个(图1),β型(OsPLDβ1和OsPLDβ2)与δ型(OsPLDδ1、OsPLDδ2和OsPLDδ3)的外显子数目相当,均达到了10个,ζ型(OsPLDζ1)外显子个数最多,达到了21个;而水稻中2个拟南芥不含有的φ和κ亚型成员(OsPLDφ和OsPLDκ)的外显子数分别为7和6个。
2.2 水稻PLD家族蛋白基序分析
MEME在线分析蛋白序列基序特征发现,水稻PLD家族含有的氨基酸残基数在511(OsPLDφ)~1164(OsPLDζ1)之间,大部分蛋白的基序集中分布在500~800位残基区域;除OsPLDφ和OsPLDκ不含有共同基序外,其它成员要都含有2或3种共同基序,如,OsPLDζ1只含1、3基序而不包含基序2,而另14个成员都具有1、2和3基序。基序1、2、3具体对应的蛋白序列见图2。
2.3 水稻PLD家族基因时空表达分析
通过提取水稻PLD家族基因的时空表达芯片数据发现,该基因家族在不同组织部位和不同发育时期呈现不同的表达特性,一般在根、茎、叶、花药、子房等部位表达相对较高,在胚芽、胚乳等部位表达较低。在α亚型中,除OsPLDα7在胚乳发育晚期表达较高而在叶中表达较低外,其它7个成员的表达趋势整体一致,都在茎、叶、根中的表达相对较高;成员OsPLDα4和OsPLDα6表现出较为相似的表达规律,在胚和胚乳中表达较低。在β亚类中,OsPLDβ1和OsPLDβ2的表达差异较大,OsPLDβ1在各部位中的表达水平普遍较低,而OsPLDβ2在各部位中的表达量要高于OsPLDβ1,特别是在子房和花药中具有较高的表达量。对于δ型成员而言,其表达趋势也不尽一致,OsPLDδ1在根中高表达,在胚乳和叶中表达较低;OsPLDδ2在花药和子房中表达较高,在胚和胚乳表达较低;而OsPLDδ3在幼穗、子房和花药等部位及胚胎整个发育阶段的表达量较高,在胚乳和叶中表达相对较低;另外,OsPLDφ、OsPLDζ1和OsPLDκ这3个成员分别在胚、子房、茎中表达相对较高(图3)。
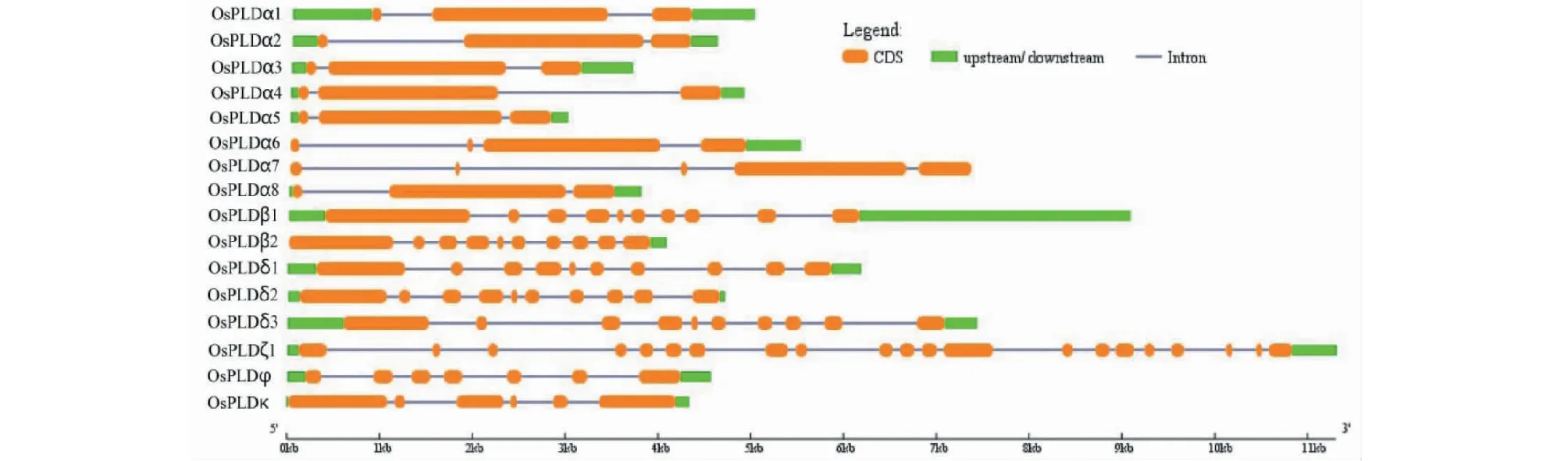
图1 水稻PLD家族基因结构分析Fig.1 Structural analysis of rice PLD-family genes
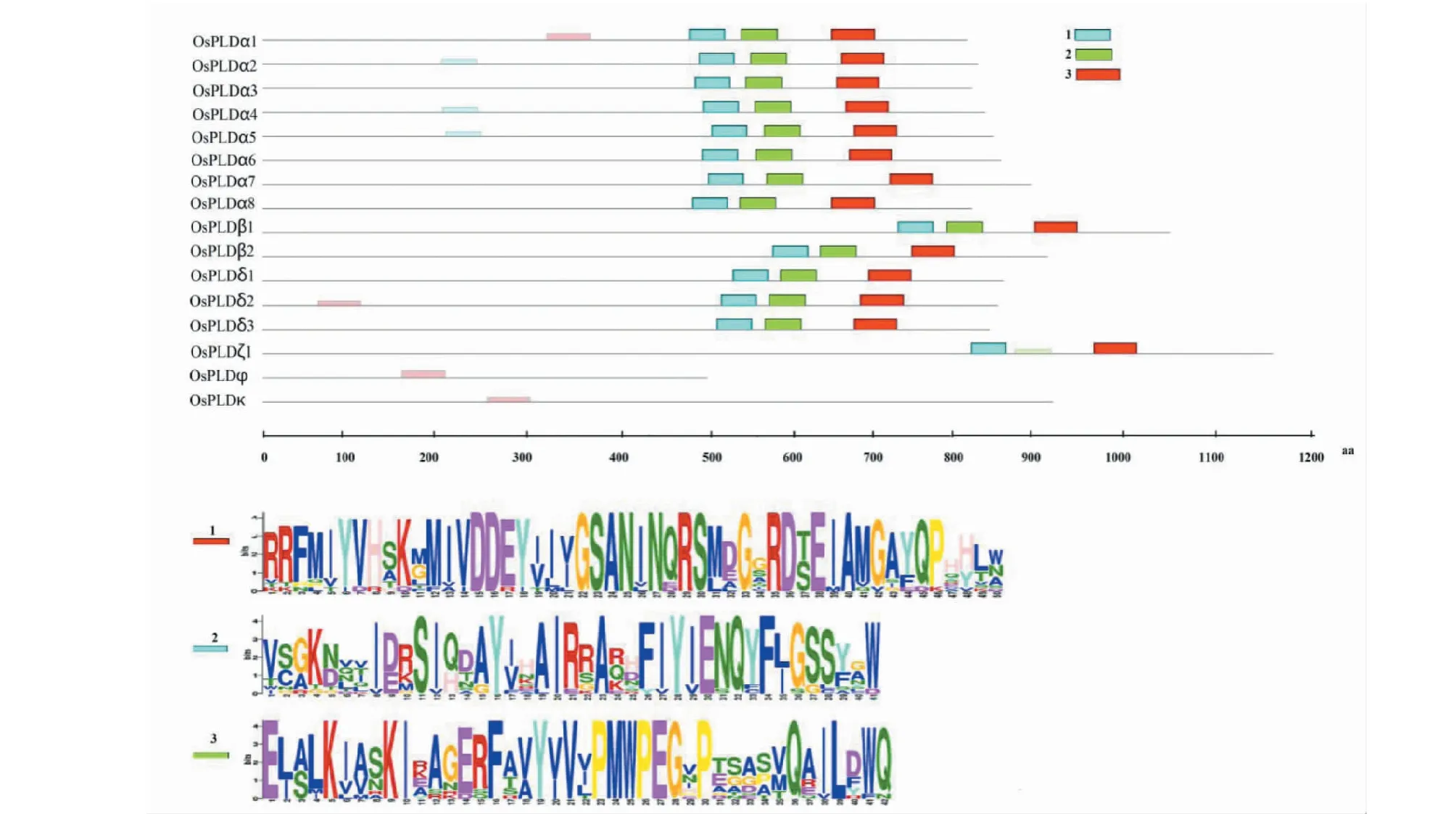
图2 水稻PLD家族蛋白基序分析Fig.2 Protein motifs analysis of PLD-family in rice
2.4 水稻PLD家族激素诱导表达分析
诱导表达谱热图分析发现,水稻PLD家族在各种激素诱导下呈现出不同的表达响应模式,一般受JA(茉莉酸)和ABA(脱落酸)诱导后表达水平发生的变化较明显,而对其它激素的敏感性相对较低。在Dα亚型中,OsPLDα3、OsPLDα4、OsPLDα5、OsPLDα8能够受JA诱导表达上调,在处理1 h后加即有较高的表达量并能维持至12 h;在ABA处理下,整体表达量偏低,处理后期的表达量(如6~12 h)要低于前期(如1 h);OsPLDα1、OsPLDα2和OsPLDα7对JA的敏感性要低于其它α亚型成员,OsPLDα1和OsPLDα2分别在ABA处理后的3~12 h和6~12 h进入高表达状态。在β类型中,OsPLDβ1、OsPLDβ2都受ABA诱导而增强表达,其中OsPLDβ1在IAA(吲哚乙酸)处理3 h时表达量明显上升,OsPLDβ2在JA处理1 h时表达量明显上调,处理6~12 h后表达下降。在δ型成员中,OsPLDδ2在JA处理下的表达受明显抑制,在BR(油菜素内酯)处理3 h时表达量较高,其它时间无明显变化;OsPLDδ1和OsPLDδ3对各种激素的诱导似乎都不太敏感,OsPLDζ1在6~12 h、OsPLDζ3在12 h处受ABA微弱的诱导作用,但不明显。对于OsPLDζ1而言,其表达受CTK(细胞分裂素)的微弱诱导,在其它激素处理下表达无明显变化。另外,特有的φ型成员OsPLDφ在各种激素作用下表达无明显变化,而κ亚型成员OsPLDκ在GA(赤霉素)和JA处理后都出现先升后降的趋势。
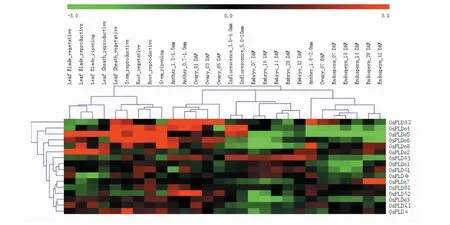
Leaf Blade: 剑叶; Leaf Sheath: 叶鞘; Root: 根; Stem: 茎; Inflorescence: 花序; Anther: 花药; Ovary: 子房; Embryo: 胚; Endosperm: 胚乳; Vegetative: 营养生长; Reproductive: 生殖; ripening: 成熟; DAF: Day after flowering, 开花后天数图3 水稻PLD家族基因组织表达分析Fig.3 Expression analysis of PLD-family genes in rice tissue

ABA:脱落酸;GA:赤霉素;IAA:生长素;BR:油菜素内酯;CTK:细胞分裂素;JA:茉莉酸图4 水稻PLD家族响应激素诱导表达分析Fig.4 Expression for rice PLD-family genes induced by hormone
3 讨 论
目前,在拟南芥和番茄等植物中已鉴定了多个PLD家族成员[19-21],发现PLD不仅能调控植物生长发育而且还能响应逆境胁迫[22-23];但是,水稻中的相关报道较少。
生信分析是预测基因功能的基本研究手段。本研究发现,尽管水稻PLD家族的基因长度不论是在亚类间还是同一亚类中都存在着较大差异,但是同一亚型内的各成员基本上都具有一致的外显子数目,比如α亚型多数成员都具有3个外显子,而β亚型和δ亚型都含有10个外显子(图1);这表明所属同一亚型的各成员具有进化和功能上的保守性,而不同亚型间所呈现的外显子数量多样性暗示了它们可能在生长发育和逆境响应过程中发挥着不同的作用。经蛋白基序分析发现,该家族整体上具有较高的相似性,除拟南芥不含有而水稻所特有的两种新型成员OsPLDφ、OsPLDκ型在基序方面各含有一段信号肽序列外,其余成员都具有2种或3种共同的结构域(图2),这表明大多数PLD可能具有相同的底物催化功能[20],而OsPLDφ、OsPLDκ的蛋白产物在细胞内的作用部位可能区别于其它成员。
有研究指出,PLD可以在各个植物组织器官(种子、果实、根、茎、叶等)中广泛表达,并且集中在生长代谢旺盛的部位,如成熟初期的种子和萌芽初期的幼苗[24-25]。本研究发现,水稻PLD的大部分成员在多数营养器官中都有较高的表达水平,而在生殖器官中的表达较低,只有OsPLDα7、OsPLDδ3和OsPLDδ1等少数基因在在生殖生长期的花序、胚或胚乳中较高表达;这说明PLD成员通过多样化的时空表达行为影响不同组织器官中的脂质代谢及信号转导途径,进而调节水稻生长发育。
在激素诱导表达中,发现该家族成员能够受不同激素影响表现出不同的应答方式,整体对ABA和JA的响应较为敏感,对BR和CTK无显著变化。ABA是植物在逆境胁迫下诱导产生的激素分子,拟南芥AtPLDα1、AtPLDα3等都与ABA有紧密联系,过表达AtPLDα1株系对ABA敏感性提高,而AtPLDα3突变株对ABA敏感性增加[7-8,11],这说明PLD从不同方向响应逆境。水稻OsPLDα3、OsPLDα4、OsPLDα5和OsPLDα8等成员在ABA处理后有被抑制倾向,而OsPLDα1、OsPLDβ1、OsPLDζ1有增强表达趋势,这可能与其基因启动子区域存在ABA和逆境响应元件有关[25]。此外,水稻PLD基因对JA具有较强的反应,如OsPLDα3、OsPLDα4、OsPLDα5、OsPLDα8、OsPLDζ1和OsPLDκ等成员均受JA影响有一定的被诱导表达的倾向,而OsPLDα2、OsPLDβ2和OsPLDδ2等受JA影响有明显的被抑制作用,故推测OsPLD成员很可能参与了JA介导的一些生物胁迫响应途径,但有待进一步验证。
本研究对水稻PLD家族进行了蛋白产物特征分析、组织时空以及响应激素的表达谱分析,这些信息将为深入研究基因家族的生物学功能提供依据。
4 结 论
水稻磷脂酶PLD家族族成员的基因长度和外显子数呈现多样化特点,所属同一亚型的各成员拥有较一致的外显子数目;大部分成员含有相同类型的基序,表明它们在功能上具有保守性和一致性。大多数水稻PLD成员在胚和胚乳中表达量低,而在各个植物组织器官(种子、果实、根、茎、叶等)中广泛表达;它们受ABA和JA诱导表达,而对BR和CTK处理不敏感,各成员在不同激素处理后表现出不同的应答方式,暗示水稻PLD成员在逆境响应中执行不同的功能。本研究结果为深入认识水稻PLD家族的生物学功能提供了信息与参考。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
南方医科大学学报(2022年3期)2022-04-13
中国医科大学学报(2021年8期)2021-09-02
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国生殖健康(2020年4期)2021-01-18
园艺与种苗(2020年12期)2021-01-08
三农资讯半月报(2020年15期)2020-08-25
肿瘤预防与治疗(2019年6期)2019-07-30
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
