
重型α-地中海贫血胎儿羊水源诱导多能干细胞的建立及鉴定
2019-11-19 01:31张娟赖春田王海燕邹闻达彭娟朱力宇李辉
中国医药导报 2019年27期
张娟 赖春田 王海燕 邹闻达 彭娟 朱力宇 李辉

[摘要] 目的 探討重型α-地中海贫血胎儿羊水源诱导多能干细胞的建立及鉴定。 方法 对胎儿被诊断为重型α-地中海贫血(--SEA/--SEA)的孕妇(16+周)进行超声引导下羊膜腔穿刺术,抽取10 mL羊水。利用非整合型仙台病毒介导Sox2、Klf4、c-Myc、Oct 3/4 四种转录因子将重型α-地中海贫血胎儿的羊水细胞重编程为诱导性多能干细胞(iPSCs);通过碱性磷酸酶(ALP)染色,免疫荧光标记(IF)检测等对iPSCs进行多能性鉴定;通过拟胚体(EB)的形成和自发分化实验对iPSCs在体内外3个胚层细胞分化的能力进行鉴定;同时进行细胞核型分析及缺失型α-地中海贫血基因诊断。 结果 仙台病毒可以成功地将重型α-地中海贫血胎儿的羊水细胞重新编程为非整合型诱导性多能干细胞(h-AF-SeV-iPSCs);ALP染色呈强阳性;IF检测显示Sox2、Oct4、SSEA-4、Tra-1-81表面特异性蛋白表达为阳性;h-AF-SeV-iPSCs悬浮培养均能形成囊性EB,贴壁培养后3个胚层均能分化;h-AF-SeV-iPSCs第10代及第20代细胞染色体核型均为正常核型(46,XY);且h-AF-SeV-iPSCs及其原代羊水细胞的重型α-地贫基因检测均为--SEA/--SEA缺失型。 结论 h-AF-SeV-iPSCs可作为研究重型α-地中海贫血胎儿宫内治疗的理想细胞模型,羊水细胞可作为iPSCs的理想细胞来源。
[关键词] 重型α-地中海贫血;羊水细胞;诱导多能干细胞;仙台病毒
[中图分类号] R714.5 [文献标识码] A [文章编号] 1673-7210(2019)09(c)-0018-06
Establishment and identification of induced pluripotent stem cells sourced fetal amniotic fluid cells of a fetus with severe α-halassemia
ZHANG Juan LAI Chuntian WAND Haiyan ZOU Wenda PENG Juan ZHU Liyu LI Hui
Department of Reproductive Center, Zhuzhou Central Hospital, Hu′nan Province, Zhuzhou 412007, China
[Abstract] Objective To explore the establishment and identification of induced pluripotent stem cells sourced fetal amniotic fluid cells of a fetus with severe α-halassemia. Methods Ultrasound-guided amniocentesis was performed on pregnant women (16+ weeks) whose fetus was diagnosed with severe α-halassemia(--SEA/--SEA), 10 mL of amniotic fluid was extracted. The amniotic fluid cells of a fetus with severe α-halassemia which were reprogrammed into induced pluripotent stem cells (iPSCs) by non-integrated sendai virus mediated transcription factors of Sox2, Klf4, c-Myc, Oct 3/4. IPSCs were multifunctionally identified by alkaline phosphatase (ALP) test and immunofluorescence (IF) labeling. The ability of iPSCs to differentiate outward from three endodermal cells in vivo was identified by embryoid (EB) formation and spontaneous differentiation test. Cell karyotype analysis and genetic diagnosis of type α-thalassemia were performed. Results Sendai virus could successfully reprogram the amniotic fluid cells of a fetus with severe α-halassemia into non-integrated induced pluripotent stem cells (h-AF-SeV-iPSCs). H-AF-SeV-iPSCs were strongly positive of ALP staining, and embryonic stem cells specific proteins of SOX2, Oct4, SSEA-4 and Tra-1-81 were positive expressed of IF staining. After the suspension culture of h-AF-SeV-iPSCs, the cystic EB body could be formed, and after the adherent culture, it could also differentiate into the three germ layers. The karyotype analysis of chromosome in the 10th and 20th generation of h-AF-SeV-iPSCs were normal (46, XY). H-AF-SeV-iPSCs and its primary amniotic fluid cells were all detected as --SEA/--SEA. Conclusion The h-AF-SeV-iPSCs could be used as an ideal cell model for study of intrauterine treatment of a fetus with severe α-halassemia, and amniotic fluid cells could be used as an ideal cells source for iPSCs.
[Key words] Severe α-halassemia; Amniotic fluid cells; Induced pluripotent stem cells; Sendai virus
地中海贫血(以下简称“地贫”)是由于珠蛋白肽链合成受到阻碍或完全被抑制,导致血红蛋白(Hb)组成异常,引起慢性溶血性贫血。不同类型的珠蛋白基因缺失或缺陷,可引起相应的珠蛋链合成受抑制,可将其分为α-地贫、β-地贫、δ-地贫、γ-地贫,及少见的δβ-地贫[1],前两种类型较为常见。各类地贫内部及与各异常Hb之间可相互组合(如Hb E/β地贫),这一系列病况又称地贫综合征,均属于不完全显性常染色体遗传病。α-地贫(α-mediterraneananemia)是由于α-珠蛋白基因的缺失或功能缺陷(点突变),导致α-珠蛋白链合成障碍所引起的一组溶血性贫血[2]。重型α-地贫如Hb Bart′s胎儿水肿综合征是由于4个α-珠蛋白基因缺少导致,胎儿出现严重的贫血症状,如不及时进行宫内治疗,大多数胎儿在孕晚期会发生死亡[3-4]。诱导性多能干细胞(induced pluripotent stem cells,iPSCs)技术是将体细胞重新编程为PSCs。选择已终末分化的干细胞导入特定转录因子,分化的细胞在特定条件下被逆转后,恢复至全能性状态或形成胚胎干细胞系(ESC)或进一步发育成一个新的个体的过程[5-6]。iPSCs的出现,避免了一些伦理问题及免疫排斥风险。目前,使用iPSCs治疗β-地貧的报道较多,但是将iPSCs用于治疗重型α-地贫的研究刚起步,而且重型α-地贫在生殖领域还没有良好的解决方案。因此,iPSCs对重型α-地贫的治疗研究将是一个重要的研究方向。本课题选择用非整合仙台病毒将重型α-地贫胎儿的羊水细胞转换成iPSCs,为重型α-地贫机制的探讨构建细胞模型。
1 材料与方法
1.1 材料
1.1.1 细胞及实验动物
实验所用的羊水细胞来源自株洲市中心医院(以下简称为“我院”)产前诊断中心。2017年10月,一对携带SEA型地贫DNA的夫妇,到我院行产前介入性检查,女方孕龄16+周,共抽取20 mL羊水,一部分用于地贫DNA验证,结果显示--SEA/--SEA缺失型;另一部分捐赠给本课题组。夫妇均知情同意,本研究经我院医学伦理委员会批准。
实验动物为6~8周龄重症联合免疫缺陷(Server combined immune-deficiency,SCID)小鼠4只,雌雄各半,体重25~33 g,合格证号:SCXK(京)2015-0001;8~10周美国癌症研究所(Institute of Cancer Research,ICR)小鼠4只,雌雄各半,体重26~32 g,合格证号:SCXK(京)2015-0001;均购买于北京维通利华实验动物技术有限公司。本研究通过中南大学实验动物伦理委员会批准。
1.1.2 主要仪器设备与试剂
实验所需仪器设备、试剂均由具有合法资质的企业生产、供应,均检验合格,试剂盒均按照操作说明严格执行。
1.2 实验方法
1.2.1 小鼠胚胎成纤维细胞(mouse embryonic fibroblasts,MEF)的制备
对8~10周ICR小鼠进行激素刺激,使小鼠均在同期发情,且雌鼠超数排卵,为保证交配,按2:1的比例将雌、雄鼠同笼喂养。将带有阴道栓的雌鼠单笼饲养0.5 d。选择12.5~13.5 d的孕鼠经颈椎脱臼后结束妊娠,75%的酒精浸泡消毒后解剖。取出的子宫浸泡在磷酸盐缓冲液(PBS)中(含双抗),然后去除胎鼠的头、尾、内脏和四肢,用PBS(含双抗)润洗躯干。剪刀物理分解躯干后,将碎小组织倒入EP管(50 mL)中,加入10 mL 0.05%胰酶消化组织5 min(晃动,37℃水浴箱),随后使用FM培养基停止消化。取细胞悬液1000 r/min离心5 min,去除上清液,FM培养基重悬细胞沉淀,并接种于10 cm培养皿中,置于37℃、5%CO2的培养箱内培养(P0代)。待MEF汇合度≥100%时,0.05%胰酶消化细胞,随后以1:3的比例传代。把MEF(P2代或P3代)制成小鼠饲养层细胞(Feeder)。
1.2.2 Feeder制备
MEF汇合度为100%时,使用PBS润洗2遍,加入含10 mg/mL丝裂霉素C的MEF培养基4 mL,培养箱内放置3 h后,去除培养基,PBS润洗3次,0.05%胰酶消化细胞,随后1000 r/min离心5 min,去除上清液。将细胞冻存留种,每冻存管106个细胞,可用于5个培养皿(35 mm)的细胞复苏。
1.2.3 羊水细胞采集及培养
B超定位穿刺吸取羊水10 mL,1000 r/min离心5 min,去除上清液,4 mL BIOAMF-2培养基重悬细胞沉淀,接种于60 mm培养皿中,置37℃、5%CO2的培养箱中培养。1周后替换成干净的羊水培养基,去除没有附着在培养皿壁上的细胞。当羊水细胞汇合度为70%~80%时,0.05%胰酶消化,并按1∶4的比例传代,传代细胞置于37℃、5%CO2的培养箱中培养,隔天换液。
1.2.4 羊水细胞重编程
在12孔板上,按每孔1.0×105~1.5×105个羊水细胞接种2孔,其中1孔用0.05%胰酶消化,室温静置直至细胞出现脱落,添加1 mL羊水细胞培养基终止消化,随后倒入15 mL圆锥离心管,1000 r/min离心5 min,计数,然后按照实际细胞的数量衡量亟待添加的仙台病毒的数量(kos MOI=5,hc-Myc MOI=5,hklf MOI=3)。将仙台病毒放在37℃水浴箱中5~10 s,解冻,超速离心后立即放入冰块中。将1 mL含有仙台病毒的羊水细胞培养基升温到37℃,汇入另1孔羊水细胞后置于37℃、5%CO2的培养箱过夜孵育。转导24 h后用新鲜的羊水细胞培养基替换含有仙台病毒的羊水细胞培养基,每隔1 d更新培养液,连续6 d,最后1 d制备Feeder培养皿。
第7天时使用0.05%胰酶0.5 mL消化转导仙台病毒的细胞,1 mL羊水细胞培养基(常温)终止细胞消化,最后全部转至EP管(15 mL),1000 r/min离心5 min,去除上清液后再次悬浮细胞,将其接种于Feeder上,细胞密度为2×104、4×104个。从第2天开始用iPSCs培养液替掉羊水细胞培养基,每天更换培养液,第8天既可隔日在显微镜下观察克隆体形成与否。转导仙台病毒3~4周后,选择物理方法挑选克隆生长的具备核仁明显以及边界清晰的单细胞集落,在覆盖基质胶(matrigel)或Feeder的培养皿中接种培养。
1.2.5 iPSCs的培养以及传代
iPSCs在37℃、5%CO2的培养箱中培养4~5 d后,借助体视学显微镜及1 mL注射器针头把克隆体分成多个小细胞团块,将其以1∶4的比例接种于包被饲养层或覆盖matrigel的培养皿中。前5代的细胞传代均选择机械切割法。从第6代开始,当细胞汇合度≥80%时,使用分散酶(dispase)或0.5 mmol/L乙二胺四乙酸二钠(Ethylenediamine tetraacetate disodium,EDTA)消化细胞,每天以iPSC/mTeSR1培养基更新旧的培养液,保证细胞正常传代。
1.2.6 多能性鉴定
1.2.6.1 碱性磷酸酶(ALP)染色 去除旧培养液,PBS换洗培养皿2、3次,4%多聚甲醛(PFA)固定细胞1~2 min,吸掉PFA后,使用PBS和TBST缓冲液润洗细胞,各2次。将溶液A(瓶盖白)、溶液B(瓶盖绿)、溶液C以50∶12.5∶437.5的剂量在室温、避光前充分混合,随后完全遮住孔底静置15~20 min,弃染色液后可用PBS润洗1次,倒置显微镜下观察染色结果并拍照。活性较好的ALP集中出现在未分化细胞中,故通过ALP染色结果验证干细胞是否发生分化,分型方法包括蓝/紫色表示未分化细胞;无特殊显色表示分化细胞。
1.2.6.2 免疫荧光标记(IF)检测 表面特异性蛋白会在胚胎干细胞表达,通过对Sox2、Oct4、SSEA-4和Tra-1-81的抗原IF染色,完成iPSCs的多能性检测。IF染色需满足iPSCs接种后细胞汇合度长至50%~60%。具体流程见下:
①固定:去掉旧的培养基,常温PBS润洗3次,每次10 min;4% PFA室温下固定细胞20~30 min;②透膜:PBS润洗3次,每次10 min,0.5%细胞通透剂TritonX-100通透细胞10 min;③封闭:PBS润洗3次,10 min/次,室溫下5%牛血清白蛋白(Bovine serum albumin,BSA)封闭细胞60 min;④一抗:加入对应一抗的试剂配比,分别为SSEA4 1∶100、Oct4 1∶500、Tra-1-81 1∶100、Sox2 1∶100。4℃过夜孵育完成抗原抗体反应;⑤二抗:PBS润洗3次,10 min/次,去掉一抗;室温下加入1∶500荧光的二抗(1 μg/mL),避光染色1 h;⑥核染:PBS洗3次,10 min/次,去掉二抗,用l μg/mL 4′,6-二脒基-2-苯基吲哚(4′,6-Diamidino-2-phenylindole,DAPI)染色10 min;⑦拍照:PBS润洗3次,10 min/次,去掉DAPI。将细胞转至专用载玻片上,在激光共聚焦倒置显微镜下拍照、并分析IF染色的结果。
1.2.7 拟胚体(EB)的形成与自发分化
1 mg/mL Dispase液消化iPSCs细胞成小团块,1000 r/min离心5 min,去掉上清液,EB培养液重新悬浮细胞,接种前述团块于低吸附培养皿中,置于37℃、5%CO2培养箱内培养;间隔一日更换培养液,连续7 d;把EB贴壁培养于4孔板(0.1%明胶包被)内,隔日替换培养液至第14天;IF染色观察3个胚层(AFP-内胚层、SMA-中胚层和Nestin-外胚层)的分化情况。
1.3细胞的核型分析
①细胞培养:iPSCs传代3~4 d后,分析在对数生长期时的细胞核型;②秋水仙素的处理过程:将加入秋水仙素(终浓度0.25 μg/mL)的细胞置于培养箱培养3 h后,0.05%胰酶消化成单体细胞,收集至15 mL的EP管中,1000 r/min离心5 min,去除上清液;③低渗:沿EP管侧壁小心加入3、4滴新鲜配制的低渗混合液(0.4%枸橼酸钠∶0.4%KCl=1∶1)共计1 mL轻柔混匀,37℃水浴箱6 min加速反应过程;④预固定:加入7滴即刻配制的固定液(冰醋酸∶甲醇=3∶1),利用吹打动作在水浴箱消掉气泡预固定,1 000 r/min离心5 min;⑤固定:去除上清,进一步打入4 mL前述固定液冲匀气泡,静置40 min,1000 r/min离心5 min。去除上清液,第3次混入2 mL固定液放置20 min,1000 r/min离心5 min,去除上清液;⑥滴片及烤片:从高处滴入固定液到冰水玻片打散气泡,迅捷扫过3遍酒精灯。60℃过夜静待反应;⑦Giemsa染色:室温下,Giemsa染色10 min后,PBS润洗染料晾干备用。⑧结果分析:每个实验组随机选取≥20个细胞中期用Leica染色体软件观察及分析。
1.4 缺失型α-地贫的DNA诊断
冰箱中取出PCR 8连管,“short”离心,赋予编号;用加样枪打入2 μL待测样于PCR 8连管中,阴/阳性对照也需一并打入;将前述8连管进行PCR扩增。配制1%琼脂糖凝胶板,0.3 g琼脂糖粉与30 mL 1× TBE在干净的三角烧瓶中加热混匀,当温度降至50~60℃时加入1.5 μL Gold View后灌胶于胶板中,室温凝固至少30 min。拔出用于制备准确间隔加样槽的梳子,将胶板浸入含1× TBE的电泳槽中,将4 μL PCR产物依次点样于琼脂糖凝胶(DNA大小分别是 0.9、1.2、1.4、1.7和2.0 kb),反应条件为130 V、45 min,用凝胶成像仪观察电泳完的凝胶,根据图像分析实验结果。2.0、1.7、1.4、1.2、0.9 kb扩增带依次对应右侧缺失(-α 3.7)、无缺失、左侧缺失(-α 4.2)、东南亚型缺失(--SEA)、泰国型缺失(--THAI)。
2 结果
2.1 重型α-地贫胎儿羊水中细胞的培养结果
置于37℃、5% CO2培养箱培养7 d后的重症α-地贫胎儿羊水细胞长成了几个相对独立的细胞小岛。更换培养液2 d后,细胞生长更加旺盛形成多个细胞集落。细胞汇合度达到70%~80%时可按1:4传代羊水细胞,后者整體生长形态似梭形或者不规则三角形,细胞核呈现卵圆形,见图1。
2.2仙台病毒诱导下重新的编程结果
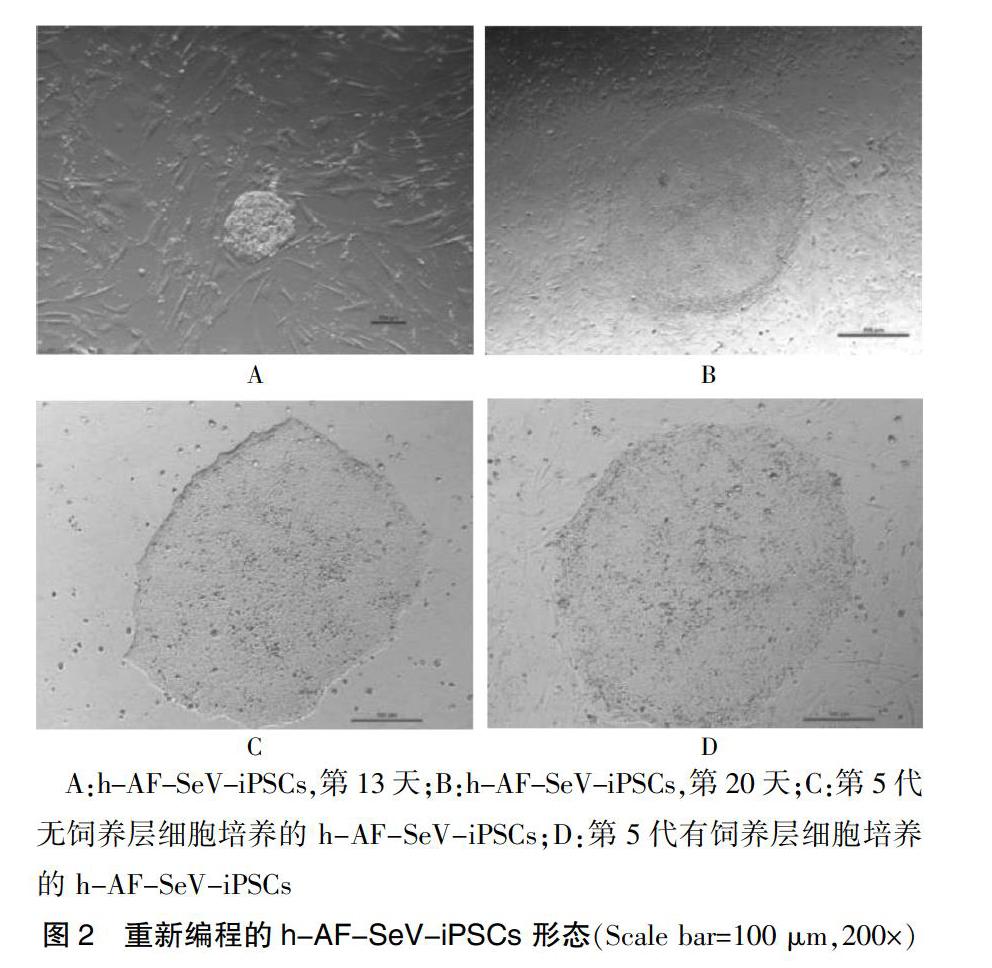
4种转录因子(Sox2、Klf4、c-Myc 及Oct3/4)通过仙台病毒介入第2代羊水细胞中。羊水细胞的胚胎干细胞样克隆现象会出现在转导的第4天。胰酶消化转导第7天的羊水细胞,而后以2×104、4×104个的密度接种,转导后陆续可见克隆样生长细胞(第9天),清晰界限位于克隆与周围饲养层细胞间(第13天),见图2A;克隆体越来越大,紧密排列的细胞间存在无法忽视的核仁且核浆比高,与周围饲养层细胞边限清楚(第20天),见图2B。1 mL注射器针头将克隆体切割接种传代,一部分在Matrigel包被的4孔板中;一部分在饲养层细胞包被的培养皿中,此过程各自使用无动物血清的mTeSR1培养基以及iPSC培养基培养。4~5 d克隆体传代1次,第5代后干细胞形态仍保留,见图2C~D。转导6×104个仙台病毒,接种在饲养层的羊水细胞会显现含有30个胚胎的干细胞样克隆体-非整合型诱导性多能干细胞(h-AF-SeV-iPSCs),经AP染色结果为阳性,转导效率在0.05%左右。
A:h-AF-SeV-iPSCs,第13天;B:h-AF-SeV-iPSCs,第20天;C:第5代无饲养层细胞培养的 h-AF-SeV-iPSCs;D:第5代有饲养层细胞培养的 h-AF-SeV-iPSCs
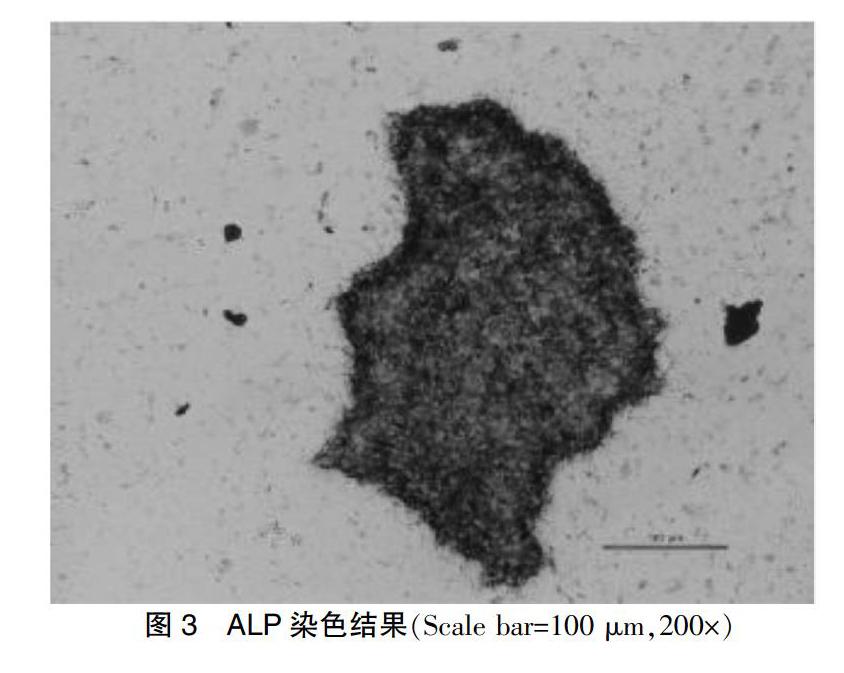
2.3 ALP染色结果
实验发现,无饲养层细胞培养的iPSCs着色明显。见图3。
2.4 IF检测结果
IF检测显示Sox2、Oct4、SSEA-4、Tra-1-81表面特异性蛋白阳性表达。见图4(封三)。
2.5 EB形成与自发分化结果
低吸附皿中,h-AF-SeV-iPSCs悬浮培养均能形成囊性拟胚体,见图5A(封三);贴壁培养后3个胚层均能分化,图5B(封三)。
2.6 染色体核型分析结果
核型分析发现,第10代和第20代 h-AF-SeV-iPSCs染色体核型都是正常的核型(46,XY)。见图6。
2.7 重型α-地贫的DNA诊断结果
h-AF-SeV-iPSCs及其原代羊水细胞的重型α-地贫基因检测均为--SEA/--SEA缺失型。见图7。
3 讨论
越来越多的研究报道指出,虽然宫内干预和产后重症监护能改善重型α-地贫胎儿的存活情况,但较多的存活胎儿仍合并不同程度、不同类型的并发症(呼吸窘迫、水肿、贫血等),通常需要终身治疗,严重影响了新生儿的生长发育[8-9]。有研究显示,宫内治疗能减少重型α-地贫胎儿的并发症,降低水肿的发生情况,从而降低窒息的风险[10]。重型α-地贫的宫内造血干细胞(HSC)移植治疗效果并不是很理想,仍需进行宫内输血治疗。而HSC来源问题是宫内HSC移植疗效不佳的重要因素。因此,采用患者来源的iPSCs对重型α-地贫胎儿进行宫内治疗具有非常重要的意义[11-12]。
现阶段,β-地贫为iPSCs治疗的重点,而α-地贫的研究报道少见。有报道利用质粒诱导技术重新编程α-地贫患者的成纤维细胞株形成iPSCs,使用同源重组技术将外源α-珠蛋白基因嵌入,通过基因修复技术使iPSCs分化成红细胞,继而表达α-珠蛋白链,平衡α/β珠蛋白链的表达[13-14]。或将重型α-地贫胎儿的皮肤成纤维细胞通过质粒转染变为iPSCs[15-16]。因此,对羊水来源的α-地贫特异性iPSCs的建立及修复具有非常重要的意义。
其他来源细胞比羊水细胞在重新编程上存在劣势。第一,羊水细胞含有羊水干细胞,与其他细胞相比,细胞相对“年轻”(胚胎一样的表观遗传学背景)。羊水细胞可快速、高效、简单地诱导成PSCs。第二,微创抽取羊水诊断是行之有效的最佳方案,且10 mL的细胞量足以保证后续的实验研究。第三,由于羊水细胞相对年轻,它引起周围环境突变的可能性微乎其微,而且遗传物质更稳定[17]。研究人员发现,再次激活Oct4基因并经过valproic acid(VPA)修饰的羊水细胞可以表现出类胚胎干细胞活性与功能[18-19]。
本研究使用仙台病毒将重型α-地贫胎儿的羊水细胞诱导成多功能干细胞。经仙台病毒介导的羊水细胞转导4 d后,可以观察到胚胎干细胞样的羊水细胞克隆;羊水细胞经病毒转导后的第7天,胰酶消化完转入饲养层细胞,克隆细胞在培养第2天出现。与文献报道的克隆时间比较[20-21],其他成纤维细胞诱导的PSCs显著减少。因此,iPSCs治疗的理想细胞来源可选羊水细胞。仙台病毒诱导PSCs只需权衡细胞添加转染试剂的量,未整合外源基因到宿主基因组中,操作简单、安全、高效、简便。本课题诱导效率在0.05%左右,与文献报道[14]基本一致。实验构建的h-AF-SeV-iPSCs和胚胎干细胞形态相同,干细胞相关多能性检测显示ALP染色明显;IF检测显示Sox2、Oct4、SSEA-4、Tra-1-81表面特异性蛋白阳性表达;h-AF-SeV-iPSCs悬浮培养均能形成囊性EB,贴壁培养后3个胚层均能分化,说明h-AF-SeV-iPSCs在体外具有向3个胚层分化的能力;核型分析发现,第10代和第20代h-AF-SeV-iPSCs染色体未出现异常,说明仙台病毒转导的羊水细胞即便经过长时间的体外培养,依旧可以通过正常染色体核型的维持来保障稳定的物质遗传过程,足以安全保障治疗未来疾病;h-AF-SeV-iPSCs及其原代羊水细胞的重型α-地贫基因检测均为--SEA/--SEA缺失型,说明长时间体外传代培养,经仙台病毒诱导的羊水细胞基因突变在未出现交叉污染细胞的前提下,维持不变。故本课题构建的重症α-地贫胎儿特异性iPSCs具有类似胚胎干细胞的自我更新和多向分化能力,可作为α-地贫进一步治疗的成熟细胞模型。
综上所述,本研究以仙台病毒为载体,虽然没有被整合,但安全性仍需关注。虽然本课题构建了重型α-地贫特异性iPSCs,但今后深入探讨应集中在利用基因编辑修复iPSCs突变基因,细胞向红系分化修复前后的iPSCs,Western blot与RT-PCR技术分析Hb A的表达及转录情况。
[参考文献]
[1] 胡俊杰,陳鑫苹,符生苗.干细胞治疗地中海贫血:现状及未来[J].中国组织工程研究,2018,22(21):3431-3437.
[2] 严金梅.重型α-地中海贫血胎儿羊水来源的诱导多能干细胞系的建立及鉴定[D].广州:广州医科大学,2017.
[3] 冼业星,严金梅,陈玉嫦,等.血红蛋白Bart′s水肿胎儿综合征诱导多能干细胞系的建立[J].实用医学杂志,2018, 34(7):1123-1127,1131.
[4] 李华晖.重型地中海贫血患者同胞造血干细胞移植后移植物抗宿主病相关危险因素分析[D].广西:广西医科大学,2017.
[5] 李继慧,覃运荣,梁毅.地中海贫血的实验室分子诊断技术研究进展[J].分子诊断与治疗杂志,2018,10(3):202-205.
[6] 刘雅丽.ssODNs结合小分子化合物对β-地中海贫血病人特异iPS细胞系基因修复的研究[D].广州:广州医科大学,2017.
[7] 黄利华,刘冬霞,李伟明,等.广东省清远地区人群α-地中海贫血基因型及临床表型特点[J].第三军医大学学报,2017,39(6):584-588.
[8] 唐燕青,陈秋莉,陈碧艳,等.15969例轻型α地中海贫血的基因型和血液学分析[J].中国计划生育学杂志,2017,25(1):57-60.
[9] 贺钰磊,徐鸣,谢雅梅,等.四川地区α地中海贫血基因检测结果及分布特征分析[J].四川医学,2017,38(11):1304-1306.
[10] 李哲涛,李伍高,唐永梅,等.东南亚缺失型α地中海贫血的胚胎植入前遗传学诊断[J].实用医学杂志,2017, 33(12):2006-2008.
[11] 罗茗月,肖克林,邵聪文,等.α地中海贫血不同基因型携带(患)者红细胞参数的差异性研究[J].中国计划生育学杂志,2017,25(7):474-478.
[12] 林芬,杨立业,林敏,等.华南人群地中海贫血的罕见基因突变[J].中华医学遗传学杂志,2017,34(6):792-796.
[13] 李荣,邓明芬,徐艳文,等.应用全基因组扩增技术对α地中海贫血进行植入前遗传学诊断的效果分析[J].生殖医学杂志,2018,27(2):166-170.
[14] 孕妇七项血液指标在东南亚型α-地中海贫血诊断、筛查中的价值分析[J].现代诊断与治疗,2017,28(2):166-170.
[15] 杜丽,王继成,秦丹卿,等.HKαα地中海贫血的基因诊断及临床表型分析[J].中国产前诊断杂志:电子版,2017, 9(3):20-22.
[16] Kwon EM,Connelly JP,Hansen NF,et al. iPSCs and fibroblast subclones from the same fibroblast population contain comparable levels of sequence variations [J]. Proc Natl Acad Sci USA,2017,114(8):1964-1969.
[17] He S,Qin Q,Huang P,et al. Characterization of a Large Novel α-Globin Gene Cluster Deletion Causing α0-Thalassemia in a Chinese Family [J]. Hemoglobin,2017,41(4-6):297-299.
[18] Para S,Mungkalasut P,Chanda M,et al. An Observational Study of the Effect of Hemoglobinopathy,Alpha Thalassemia and Hemoglobin E on P.Vivax Parasitemia [J]. Mediterr J Hematol Infect Dis,2018,10(1):e2018015.
[19] de Azevedo LA,Bonazzoni J,Wagner SC,et al. Do Alpha Thalassemia,Fetal Hemoglobin,and the UGT1A1 Polymorphism have an Influence on Serum Bilirubin Levels and Cholelithiasis in Patients with Sickle Cell Disease? [J]. Mol Diagn Ther,2017,21(4):437-442.
[20] Hsu SY,Chen SH,Fu RH. Alpha-thalassemia major and restrictive foramen ovale in a preterm neonate without severe hydrops fetalis [J]. Pediatr Neonatol,2017,58(6):546-548.
[21] Nguyen LS,Redheuil A,Mangin O,et al. Sickle-cell and alpha-thalassemia traits resulting in non-atherosclerotic myocardial infarction:Beyond coincidence? [J]. World J Clin Cases,2017,5(12):428-431.
(收稿日期:2019-03-07 本文编辑:任 念)
猜你喜欢
科学大众(2021年6期)2022-01-01
基层中医药(2021年4期)2021-07-22
基层中医药(2021年4期)2021-07-22
阅读(科学探秘)(2021年9期)2021-05-30
考试与评价·高二版(2020年4期)2020-09-10
家庭医学(下半月)(2020年6期)2020-08-24
橡塑技术与装备(2018年14期)2018-07-20
销售与市场(管理版)(2017年3期)2017-03-28
西藏科技(2016年8期)2016-09-26
中国卫生标准管理(2015年7期)2016-01-15
