
闽楠微扦插繁育体系的建立
2020-01-18 02:16肖业玲童再康吴建辉
浙江农林大学学报 2020年1期
肖业玲, 童再康, 吴建辉
(1. 浙江农林大学 省部共建亚热带森林培育国家重点实验室, 浙江 杭州311300; 2. 浙江省建德市三都镇人民政府, 浙江 建德311605)
闽楠Phoebe bournei为樟科Lauraceae 楠属Phoebe植物, 常绿乔木, 分布于江西、 福建、 浙江南部、广东、 广西等地的亚热带常绿阔叶林地带, 为中国特有的国家二级保护植物。 闽楠材质优良、 用途广泛, 有很高的经济价值, 是优良珍贵用材和园林绿化树种[1-2]。 然而, 由于长期人为破坏, 闽楠自然繁育率低, 自然资源已接近枯竭[3]。 种子繁殖是楠木常规的繁殖方式[4], 但是种苗繁殖效率低, 生长周期长, 具有很高的杂合性, 很难保持其优良性状。 目前, 在扦插繁殖中, 基质、 生根促进剂质量浓度、 季节等3 种因子都会影响闽楠嫩枝扦插生根率及幼根生长[5-6]。 植物组织培养技术是建立珍贵树种无性繁殖体系的重要手段, 这一技术不仅能缩短育种周期, 在短时间内快速繁育良种, 还能为转基因平台的搭建打下基础。 以闽楠近成熟种子为外植体进行组培快繁技术研究未见报道。 因此, 本研究以闽楠近成熟胚为外植体建立闽楠微扦插繁殖体系, 探讨抗褐化剂和生长素对近成熟胚萌发以及微扦插生根的影响,以期获得闽楠微扦插繁殖的最佳培养条件, 为闽楠良种的快速繁殖、 后续生理及分子研究提供参考。
1 材料与方法
1.1 试验材料
闽楠果实在未成熟时为青绿色, 质地较硬, 等到发育至成熟时果皮变为紫黑色, 同时果肉变软, 挤压有汁液流出。 由于未成熟的闽楠种子组培萌发率较低, 而成熟种子外部的果肉易霉烂, 组培污染率较高, 因此以果皮颜色变为紫黑色而果肉未明显变软的近成熟果作为试验材料。
2017 年11 月19 日于江西婺源上梅洲塘村和马家村采集闽楠近成熟果实。 选取长势良好、 树干通直、 结实量较多的7 个株系, 采集光滑饱满、 无病虫害的果实, 沙藏保存, 每隔5 d 喷水1 次。
1.2 试验方法
1.2.1 外植体消毒处理 取闽楠近成熟果实于组培瓶中, 流水冲洗2 h 后, 用体积分数为75%的乙醇浸泡30 s, 无菌水冲洗3~4 次后, 加入质量分数为0.1%的氯化汞(HgCl2)溶液消毒8~10 min, 期间摇动组培瓶2~3 次, 使消毒液与果实充分接触, 随后用无菌水冲洗4~5 次。 在超净工作台上, 用无菌滤纸吸干表面水分, 用镊子将果皮拨开, 用解剖刀将种皮划开。 将去掉种皮的胚快速接种至培养基中, 每瓶接种2~4 个。 培养温度为(25±2) ℃, 光照/黑暗周期为18 h/6 h, 光照强度为2 000~2 500 lx。
1.2.2 胚褐化试验 以1/2 MS 为基础培养基, 添加0.2 mg·L-1的IBA。 将种子消毒后去掉种皮, 分别切去全部子叶、 切去一半子叶, 与保留全部子叶(ck)作为对照组, 每组试验至少接种5 瓶, 每瓶4~6 个胚, 重复3 次, 统计褐化率。 褐化率=褐化植株数量/接种植株数量×100%。
1.2.3 褐化程度和褐化多酚氧化酶(PPO)活性 以1/2 MS 为基础培养基, 添加0.2 mg·L-1的IBA, 分别添加1.00、 2.00、 4.00 g·L-1的聚乙烯基吡咯烷酮(PVP)和0.25、 0.50、 1.00 g·L-1的维生素C(VC)。 取接入培养基0, 3 和10 d 的7 号株系近成熟胚, 测定其褐化程度等生理指标。 褐化程度: 取0.5 g 胚, 加入预冷磷酸缓冲液(pH 7.0)3 mL, 快速研磨, 将研磨液倒入离心管中, 用7 mL 缓冲液冲洗研钵, 合并后4 ℃离心(1 200×g)10 min, 取上清液3 mL 于比色皿中, 在475 nm 测光密度, 重复3 次。 以光密度代表褐化程度, 光密度D(475)越大代表褐化程度越大[7]。 PPO 活性: 取0.5 g 胚, 加入预冷的磷酸缓冲液(pH 7.0)3 mL, 研磨, 转移至离心管中, 再用7 mL 缓冲液冲洗研钵, 合并提取液, 4 ℃离心(1 200×g)10 min, 取上清液为粗酶液, 将0.5 mL 邻苯二酚加入2 mL 磷酸缓冲液(pH 7.0)中, 加入0.5 mL 酶提取液, 立即于波长410 nm 下测定光密度, 2 min 后再计光密度, 以不加酶提取液的反应液为对照, 重复3次, 以2 min 内光密度上升1 为1 个酶活单位[8]。
1.2.4 不同抗褐化剂对胚萌发试验 采用的实验材料为3 号株系, 以1/2 MS 为基础培养基, 添加0.2 mg·L-1IBA, 分别添加1.00、 2.00、 4.00 g·L-1PVP 和0.25、 0.50、 1.00 g·L-1VC, 每组培养基均添加30 g·L-1蔗糖, 7 g·L-1琼脂, pH 调至6.0, 每个处理接种15 瓶, 接种2~4 个胚·瓶-1, 重复3 次。 培养45 d后统计萌发情况, 并计算每个处理相应的萌发率。 萌发率=萌发植株数量/接种植株数量×100%。
1.2.5 不同吲哚丁酸(IBA)质量浓度对胚萌发试验 以1/2 MS 为基础培养基, 设计单因素试验, 分别添加0.2、 0.5、 1.0、 1.5 mg·L-14 个质量浓度的IBA, 以不添加植物生长素为对照(ck), 附加2 g·L-1PVP,7 g·L-1的琼脂, 蔗糖30 g·L-1, pH 调至6.0, 每个处理接种15 瓶, 重复3 次。 培养45 d 后统计萌发情况, 并计算出每个处理相应的萌发率。
1.2.6 不同抗褐化剂对微扦插生根试验 采用5 号株系作为实验材料, 以1/2 MS+1.5 mg·L-1IBA 为基础培养基, 分别加入0.25、 0.50、 1.00 g·L-1VC 和1.00、 2.00、 4.00 g·L-1PVP, 以不添加任何抗褐化剂的基础培养基作为对照(ck), 每个处理接种10 瓶, 接种3~5 个芽·瓶-1, 重复3 次。 30 d 后统计生根率、 平均生根数和平均根长。 生根率=生根植株数量/接种植株数量×100%; 平均生根数=所有生根植株总的生根数量/生根植株数量; 平均根长=所有生根的长度总和/生根植株数量。
1.2.7 不同质量浓度IBA 对无菌苗微扦插试验 以1/2MS 为基础培养基, 采用6 号株系, 分别添加0.2和1.5 mg·L-12 个IBA 质量浓度, 以不添加IBA 的培养基为对照(ck), 附加2 g·L-1PVP, 7 g·L-1的琼脂, 蔗糖30 g·L-1, pH 调至6.0, 每个处理接种15 瓶, 重复3 次。 培养45 d 后统计生根情况, 并计算出每个处理相应的生根率。
1.2.8 繁殖系数 闽楠近成熟胚接种30 d 后去掉顶芽, 60 d 后侧芽可用于微扦插, 微扦插苗生长60~90 d 后可进行移栽, 每年每株无菌苗最多可剪取4 次芽用于微扦插。 去掉顶芽60 d 后统计长度大于1 cm 芽的数量, 每个株系至少统计5 株。 繁殖系数=平均新生侧芽数量。
1.3 数据统计与处理
采用Excel 2007 与SPSS 20.0 对数据进行统计分析。
2 结果与分析
2.1 子叶切除处理下的胚褐化
从表1 可见: 虽然不同株系胚褐化率有所差异, 但是整体表现出相似的规律, 即保留全部子叶时褐化率最高, 其中1 号株系褐化率最高, 达60.81%, 而不留子叶时褐化率最低, 在6 号株系中褐化率仅为13.84%。 然而, 对萌发后无菌苗后续生长观察发现, 切除全部子叶的无菌苗较细长, 长势较差, 真叶无法正常展开。 因而, 后续研究中采用保留全部子叶、 通过抗褐化剂控制子叶褐化程度的方法来提高胚的萌发率并保证无菌苗的生长。
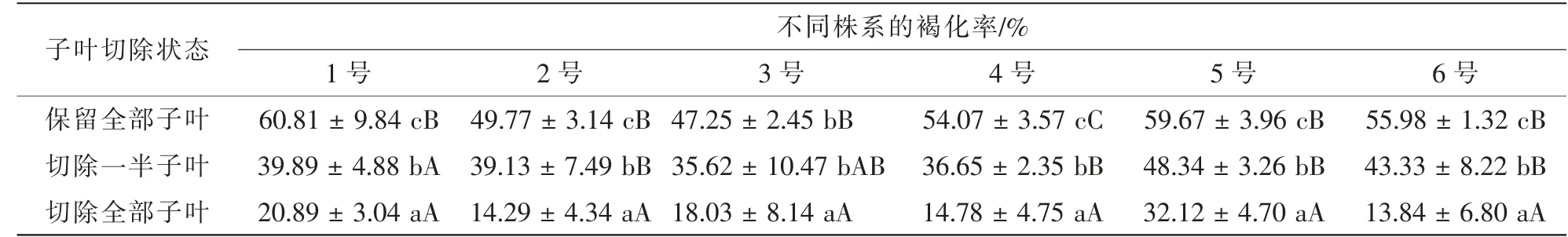
表1 子叶切除对胚褐化的影响Table 1 Effect of cotyledonectomy on embryo browning
2.2 不同抗褐化剂下子叶褐化和PPO 活性
2.2.1 子叶褐化 如图1A 所示: 闽楠近成熟胚在1/2 MS 培养基中培养3 d 时开始发生褐变, 在未加入任何抗褐化剂时, 种子褐化程度较重, 10 d 时光密度D(475)达1.2, 且萌发较为迟缓。 而添加VC 和PVP 的种子褐化程度降低, 生长状况渐好。 不同种类和质量浓度的抗褐化剂对抑制褐化的效果不同, 添加了PVP 或VC 时在第3 天时都有不同程度褐化, 其中褐化程度为添加1.00 g·L-1PVP>2.00 g·L-1PVP>4.00 g·L-1PVP。 VC 褐化值在0~10 d 均低于PVP 和ck, 到第3 天时, 添加0.50 和1.00 g·L-1VC 的褐化值均下降, 到了第10 天时, VC 褐化程度均呈上升趋势, 当VC 的添加量为1.00 g·L-1, 培养10 d 时,种子褐化率最低, 为25%。
2.2.2 PPO 活性 分析PPO 活力(图1B)可见: 在添加了PVP 的培养基中, 子叶中PPO 活性在第3 天时与ck 大致相等, 呈平稳状态, 到第10 天时, PVP 组的活性高于ck 组, 其中, PPO 活性为添加4.00 g·L-1PVP>1.00 g·L-1PVP>2.00 g·L-1PVP。 VC 组在整个过程中, 子叶中PPO 活性一直呈下降趋势, 第3天时, PPO 活性为添加1.00 g·L-1VC>0.25 g·L-1VC>0.50 g·L-1VC, 到了第10 天, PPO 活性为添加0.25 g·L-1VC>0.50 g·L-1VC>1.00 g·L-1VC, 且一直低于PVP 组和ck 组。 因此, 添加的最适抗褐化剂为2.00 g·L-1PVP。
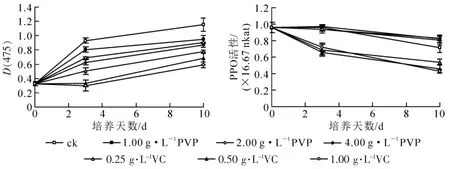
图1 不同抗褐化剂对子叶褐化及PPO 活性的影响Figure 1 Influence of different anti-browning agents on browning and PPO activity
2.3 不同抗褐化剂对胚萌发的影响
从表2 可见: 以1/2 MS 为基础培养基, 添加0.2 mg·L-1IBA, 3 号株系在添加PVP 诱导近成熟种子胚的萌发效果较好, 而添加2.00 g·L-1PVP 的效果最好, 萌发率为83.57%, 添加VC 时胚萌发则较差,在添加0.25 g·L-1时, 胚萌发率为53.06%。
2.4 不同质量浓度IBA 对胚萌发的影响
从表3 可以看出: 闽楠近成熟胚萌发对IBA 较为敏感, 较低质量浓度的IBA 诱导即可萌发, 且萌发率较高。 IBA 质量浓度为0.2 mg·L-1时效果最好, 萌发率最高的6 号株系可达89.71%。 随着IBA 质量浓度的升高萌发率下降, IBA 质量浓度达1.0 mg·L-1及以上时, 萌发率低于ck。 闽楠近成熟胚萌发较适IBA 质量浓度为0.2 mg·L-1。

表2 不同抗褐化剂下胚萌发的变化Table 2 Change of different anti-browning agents on cotyledon embryo germination

表3 不同质量浓度IBA 下胚萌发的变化Table 3 Change of different concentrations of IBA on cotyledon embryo germination
2.5 不同抗褐化剂对无菌苗微扦插生根的影响
从表4 可见: 以添加1.5 mg·L-1IBA 的1/2 MS 培养基为基础培养基, 添加PVP 诱导闽楠无菌苗微扦插生根效果较好, 顶芽生根率均达100%。 其中, 添加2.00 g·L-1PVP 在接种15 d 开始生根, 至30 d 时,顶芽平均生根数为3.86 条, 平均根长1.56 cm; 侧芽生根率为60.27%, 平均生根数为1.55 条, 平均根长0.85 cm。 添加VC 时生根效果较差, 仅在添加0.25 g·L-1时有66.81%的顶芽生根, 但是生根数量与平均根长均明显低于添加PVP 的培养基, 而随着质量浓度的升高, 顶芽微扦插后无法生根。 在添加了VC 的生根培养基中, 侧芽均无法生根。 在不添加任何抗褐化剂(ck)中顶芽生根情况也没有添加PVP 效果好, 闽楠近成熟种子微扦插生根较适用的抗褐化剂为2.00 g·L-1PVP。
2.6 不同质量浓度IBA 对无菌苗微扦插的影响
如图2 所示: 闽楠微扦插生根较慢, 平均15 d 左右开始生根。 从表5 可以看出: 在无菌苗继代生根培养中, 1.5 mg·L-1IBA 效果最好, 生根率最高, 顶芽生根率均达100%, 15 d 可发新根, 平均生根数约2.85 条, 平均根长为1.07 cm, 侧芽生根率为52.24%, 平均生根数约1.48 条, 平均根长为0.56 cm。闽楠无菌苗微扦插生根较适培养基为1/2 MS+1.5 mg·L-1IBA+2.00 g·L-1PVP。

表4 不同抗褐化剂下微扦插生根的变化Table 4 Change of different anti-browning agents and concentrations on rootage after microcuttage

表5 不同质量浓度IBA 处理下无菌苗微扦插的变化Table 5 Change of different concentrations of IBA on micro-cutting of sterile seedlings
2.7 不同株系的繁殖系数
从表6 可见: 4 号、 5 号、 6 号株系生根率均为100%。 其中4 号株系的无菌苗芽小而短, 可用于微扦插的芽较少, 因而繁殖系数偏低, 而3 号株系有较多丛生芽现象出现(图2G~I), 繁殖系数高。 综上,闽楠无菌苗微扦插体系中, 3 号、 5 号、 6 号株系表现较好, 其中3 号家系繁殖系数为24.40。

表6 不同株系繁殖系数的变化Table 6 Change of reproductive coefficient of different families
3 结论与讨论
PPO 是一类广泛存在于植物中的氧化酶[9], 在褐化过程中起关键作用[10]。 当细胞中的膜结构遭到破坏后, 原本绑定于膜上的PPO 随即被激活[11], 将酚类物质氧化生成醌类物质, 再经非酶促反应聚合形成褐色物质, 对植物产生毒害作用[12]。 褐化发生与外植体体内酚类物质的含量呈正相关[13]。 由于细胞中含有较多的酚类化合物, 闽楠近成熟胚在组培体系中萌发时, 子叶褐化情况较为严重, 对其萌发以及后续的生长均会产生不利影响。 在切除子叶后发现, 胚在培养基中的萌发率明显提高, 但由于缺乏子叶所提供的营养, 其后续生长较为缓慢, 叶片无法正常展开。 因此, 本研究保留子叶以保证无菌苗萌发后的生长, 同时通过添加抗褐化剂的方法来抑制子叶褐化。 抗褐化剂按作用原理可主要分为2 种: 一种是具有还原能力的抗氧化剂, 另一种是具有多孔洞结构的吸收剂。 还原型抗氧化剂主要有维生素C(VC)、 硫代硫酸钠(Na2S2O3)和硝酸银(AgNO3)等, 添加这一类抗褐化剂可以改变外植体周围的氧化还原电势, 从而抑制酚类氧化, 减轻褐变。 而吸收型抗褐化剂主要有聚乙烯吡咯烷酮(PVP)和活性炭(AC)等, 这类抗褐化剂具有特殊的孔洞结构, 能够吸收固定酚类物质, 阻止其与PPO 的接触, 从而起到减轻褐化的效果[14]。 本研究发现: 还原型抗褐化剂VC 能够抑制PPO 活性, 对子叶褐化的抑制能力较强, 但是它同时也影响了其他生理过程, 最终造成胚萌发受阻。 而吸收型抗褐化剂PVP 不影响PPO 活性, 对子叶褐化有一定的抑制能力, 能够显著提高胚的萌发率, 同时对无菌植株的后续生长也无明显抑制。 因此, 本研究中闽楠胚萌发培养基中添加2.00 g·L-1PVP 显著抵制PPO 活性, 这与陈梦倩等[15]研究的抗褐化剂对香港四照花Dendrobenthamia hongkongensis褐化的影响试验结果一致。
适宜质量浓度的抗褐化剂不仅能够缓解子叶的褐化和促进胚的萌发, 同时在微扦插生根时也能起到积极作用。 本研究中, 添加2.00 g·L-1PVP 时顶芽和侧芽的生根情况最好, 与不添加任何抗褐化剂的对照组(ck)相比, 所生出的根较为粗壮, 颜色也较浅。 而VC 作为一种还原剂, 虽然能强烈抑制褐化, 但同时也抑制了植物组织其他正常的生理活动, 导致微扦插生根率降低。
添加适量的IBA 能够显著提高生根率和不定根以及试管苗的生长速度。 本研究表明: 添加0.2 mg·L-1IBA 能够促进闽楠近成熟胚萌发。 随着IBA 质量浓度的升高, 萌发率下降。 IBA 质量浓度达1.0 mg·L-1及以上时, 根粗短、 植株小、 长势慢, 不利于后期生长, 这与二花蝴蝶草Torenia biniflora的组织培养及植株再生现象相似[16]。 添加高质量浓度的1.5 mg·L-1IBA 能够促进闽楠无菌苗生根, 顶芽生根率为100%, 平均生根数为3.86 条, 平均根长为1.56 cm; 侧芽生根率为60.27%, 平均生根数为1.55 条, 平均根长为0.85 cm, 比添加低浓度IBA 效果显著。

图2 闽楠近成熟胚微扦插繁育情况Figure 2 Micro-cutting and breeding of near mature embryos of P. bournei
猜你喜欢
特种经济动植物(2022年2期)2022-02-15
农业与技术(2021年21期)2021-11-17
园艺与种苗(2021年8期)2021-09-23
安徽农业科学(2021年11期)2021-07-16
绿色科技(2021年11期)2021-07-01
安徽农学通报(2021年10期)2021-06-17
新农业(2020年15期)2020-12-19
农民致富之友(2020年22期)2020-09-02
新农业(2020年8期)2020-08-26
现代畜牧科技(2016年5期)2016-10-21
