
植物源化合物肉桂醛对单核细胞增生李斯特氏菌的抑制作用
2020-02-22 06:22王帆刘小莉
江苏农业科学 2020年24期
关键词:抑制作用
王帆 刘小莉
摘要:研究了植物源化合物肉桂醛对单核细胞增生李斯特氏菌(Lm)的抑制作用。肉桂醛对Lm的最小抑制浓度和最小杀菌浓度分别为150、300 μg/mL。经过肉桂醛处理的Lm细胞出现菌体细胞变形、膨大的现象,胞外的葡萄糖和核酸含量明显高于对照,胞内蛋白含量降低,说明肉桂醛处理使得Lm细胞通透性增大,并且抑制了Lm的蛋白合成。流式细胞仪分析结果显示,最小杀菌浓度的肉桂醛处理6 h后,细胞死亡率可达到95.6%。
关键词:肉桂醛;单核细胞增生李斯特氏菌;抑制作用
中图分类号: S182 文献标志码: A 文章编号:1002-1302(2020)24-0194-04
单核细胞增生李斯特氏菌(Listeria monocytogenes,Lm)是一种人畜共患的食源性致病菌,主要存在于水产品、乳品、肉品等动物性食品中,对特定人群(孕妇、新生儿、老人以及免疫缺陷者)的潜在威胁很大,致死率高达20%~30%[1-2]。与大多数病原菌不同的是,Lm在低温条件下仍可生长繁殖,在食品加工与贮藏过程中容易造成污染引起食物中毒,随着生活节奏的加快,人们对冷藏以及即食食品的需求比重日益加大,Lm潜在的危险性也逐渐凸显。现有的防控措施中,化学方法可以快速杀菌但也存在安全隐患,因此开发安全高效的新型抑菌剂具有巨大的应用潜力。
肉桂醛(cinnamaldehyde)在自然界主要存在于樟属植物中,工业上通常以化学合成的方法进行制备。它的应用极其广泛,在医药领域被用于抗肿瘤、抗病毒等,在食品领域被用作食用香料以及防腐保鲜剂[3]。近年研究表明,肉桂醛对Lm表现出较好的抑制效果[4-6],但肉桂醛对Lm的作用机制却很少有报道。本研究探索肉桂醛抑制Lm的作用机制,为将肉桂醛开发成为新型高效抑菌剂提供理论依据。
1 材料与方法
1.1 供试菌株和试剂
Lm CICC21532采购自中国工业微生物菌种保藏中心,脑心浸液(BHI)培养基采购自北京陆桥公司,肉桂醛采购自国药集团,羧基荧光素二乙酸酯(cFDA)、碘化丙啶(PI)采购自美国Sigma公司。
1.2 菌液和药液制备
将Lm接种于无菌BHI培养基中,37 ℃培养 16 h 至对数期,以BHI培养基调整菌液浓度至1×107 CFU/mL备用。以乙醇作溶剂将肉桂醛配置成浓度为1×105 μg/mL的母液。
1.3 Lm对肉桂醛的敏感性测定
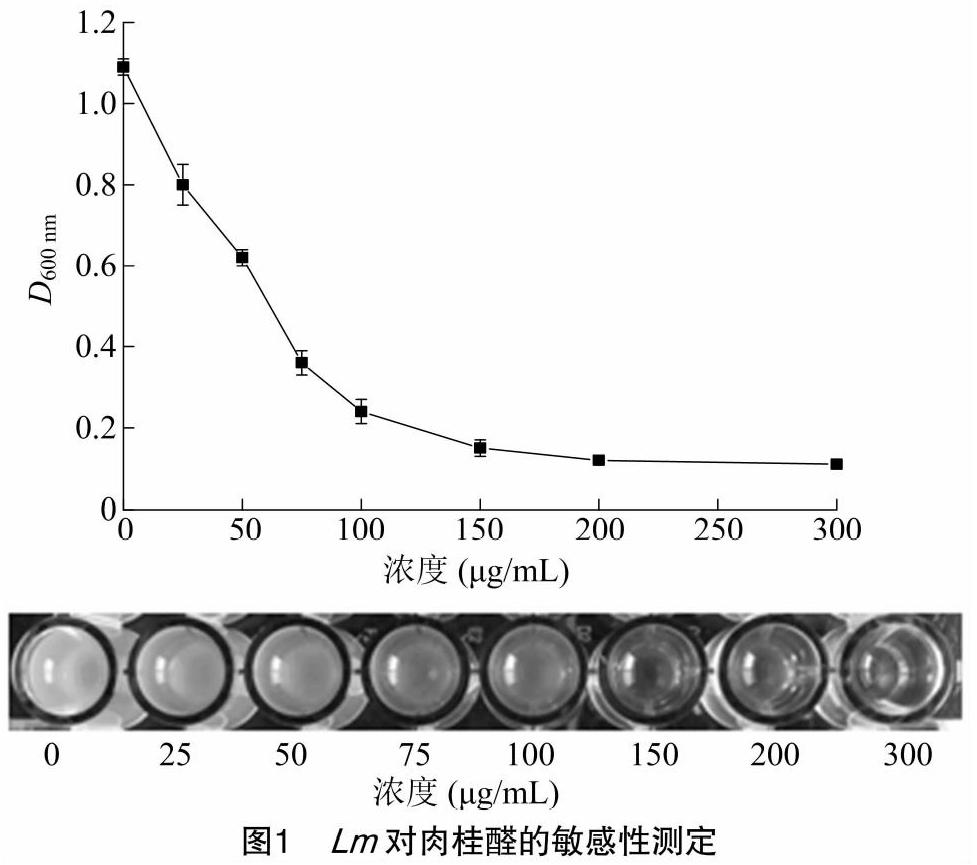
在96孔板上将肉桂醛母液用BHI培养基进行梯度稀释,使其作用浓度分别为0、25、50、75、100、150、200、300 μg/mL,菌液浓度为1×107 CFU/mL,37 ℃培养24 h。采用超微量微孔板分光光度计于600 nm测定96孔板的吸光度,测定肉桂醛对Lm的最小抑制浓度(MIC)。在没有浑浊的微孔中接种50 μL培养液涂布于BHI琼脂培养基上,37 ℃培养24 h,测定肉桂醛对Lm的最小杀菌浓度(MBC)。
1.4 肉桂醛对Lm细胞形态的影响
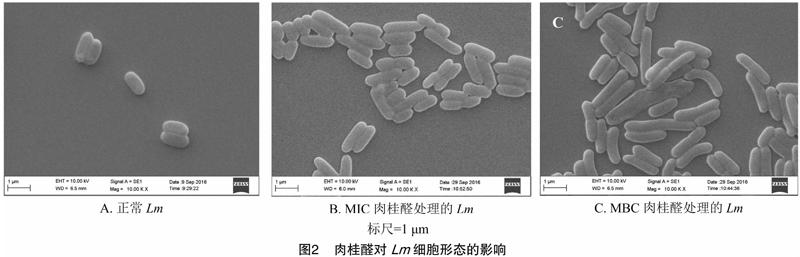
Lm培养至对数期,加入肉桂醛使其作用浓度分别为MIC和MBC,用无菌水作空白对照,37 ℃、120 r/min 处理6 h后离心收集菌体,用0.1 mol/mL pH值7.4的磷酸缓冲液洗涤3次,加入2.5%戊二醛于4 ℃过夜固定。菌体采用体积分数为30%、50%、70%、90%、100%的乙醇溶液脱水置换,每次15 min,脱水后临界点干燥,将干燥的菌体固定到金属台上,用离子溅射仪对金属台喷金、镀膜,扫描电镜观察拍照。
1.5 肉桂醛对Lm细胞通透性的影响
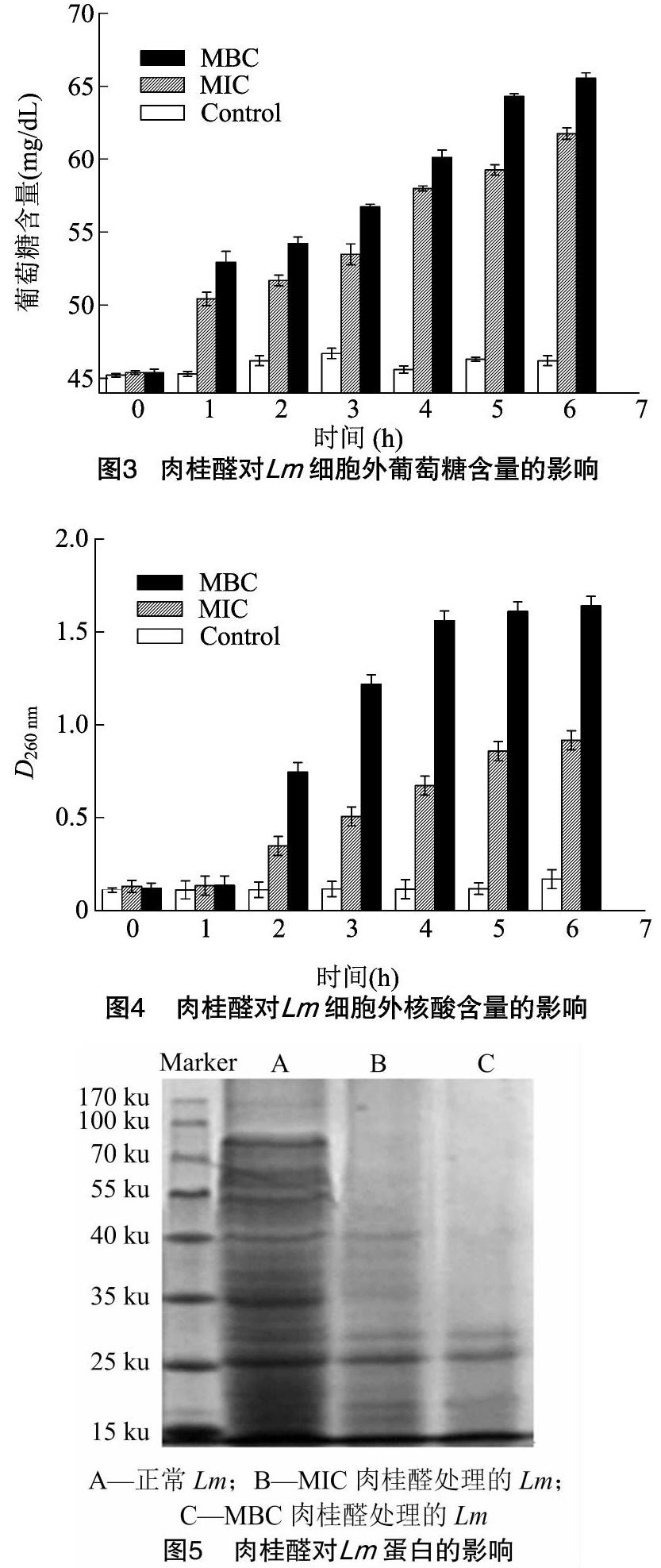
Lm培养至对数期,用无菌磷酸缓冲液(0.1 mol/mL,pH值7.4)洗涤重悬,加入肉桂醛使其作用浓度分别为MIC和MBC,用无菌水作空白对照,置于37 ℃、120 r/min条件下培养,分别于0、1、2、3、4、5、6 h取样,离心收集上清液,采用F006试剂盒(南京建成生物工程研究所)测定葡萄糖含量,260 nm紫外吸光度反映核酸含量的变化。
1.6 肉桂醛对Lm细胞蛋白的影响
Lm培养至对数期,用无菌磷酸缓冲液(0.1 mol/mL,pH值7.4)洗涤重悬,加入肉桂醛使其作用浓度分别为MIC和MBC,用无菌水作空白对照,37 ℃、120 r/min处理6 h,经细胞破碎仪破碎后离心,取沉淀加入等体积上样缓冲液混合,于100 ℃沸水浴中保温5 min,取出冷却至室温后进行SDS-PAGE凝胶电泳(分离胶12%,浓缩胶4%,电压120V)。用染色液染色2~3 h,再用脱色液进行处理后观察拍照。
1.7 Lm细胞死亡的测定
Lm培养至对数期,用无菌磷酸缓冲液(0.1 mol/mL,pH值7.4)洗涤重悬,加入肉桂醛使其作用浓度分别为MIC和MBC,用无菌水作空白对照,37 ℃、120 r/min处理6 h。菌悬液样品加入cFDA荧光探针(终浓度50 μg/mL),37 ℃避光装载10 min,无菌磷酸缓冲液洗涤后加入PI荧光探针(终浓度30 μg/mL),室温避光装载10 min,无菌磷酸缓冲液洗涤重悬后进行流式细胞仪分析检测。流式细胞仪激发波长488 nm,检测器FL1检测波长525 nm,检测器FL3检测波长620 nm,cFDA进入FL1荧光通道,PI进入FL3荧光通道,获取FL1-FL3散点图。
1.8 数据处理与分析
所有试验均进行3次重复,应用SPSS 17.0软件对试验数据进行显著性分析(P<0.05),Origin 8.5软件绘图。
2 結果与分析
2.1 Lm对肉桂醛的敏感性测定
Lm对肉桂醛的敏感性结果见图1。Lm经肉桂醛处理后生长受到明显的抑制作用,随着处理浓度的增大,D600 nm逐渐降低,当浓度达到150 μg/mL时,微孔中观察不到浑浊,因此肉桂醛对Lm的MIC为150 μg/mL。在无浑浊的微孔中接种50 μL培养液涂布于BHI琼脂培养基上,37 ℃培养24 h,完全无细菌生长的最小浓度为300 μg/mL,即为肉桂醛对Lm的MBC。
2.2 肉桂醛对Lm细胞形态的影响
肉桂醛对Lm细胞形态的影响见图2。正常Lm呈短杆状,菌体形态饱满,细胞表面光滑。MIC浓度的肉桂醛处理后,Lm菌体细胞变长,细胞之间的界限变得模糊。MBC浓度的肉桂醛处理后,部分细胞出现变形、膨大的现象。肉桂醛作用后Lm细胞表面并未观察到明显的凹陷和穿孔。
2.3 肉桂醛对Lm细胞通透性的影响
核酸中的嘌呤和嘧啶碱基在260 nm附近有最大吸收峰,通过测定260 nm处的紫外吸光度可以反映细胞中核酸的泄漏情况[7]。从图3、图4可以看出,肉桂醛处理1 h后,Lm细胞外的葡萄糖含量明显(P<0.05)高于空白对照,处理2 h后,核酸含量也明显(P<0.05)升高,且含量随着时间和浓度的增加逐渐升高。说明肉桂醛处理导致Lm细胞的通透性增大,细胞内的物质向细胞外流失。
2.4 肉桂醛对Lm细胞蛋白的影响
肉桂醛对Lm细胞蛋白的影响见图5,对分子量在15~170 ku的Lm细胞蛋白进行SDS-PAGE电泳分析,正常Lm蛋白电泳条带完整清晰,MIC肉桂醛处理后,Lm的电泳条带明显变浅,MBC肉桂醛处理后,部分条带逐渐缺失,缺失部分为分子量在35~170 ku之间的蛋白。 结果表明 肉桂醛处理使得Lm细胞蛋白含量降低,一方面肉桂醛抑制了Lm的蛋白特别是大分子量蛋白的合成,进而导致菌体生长受阻死亡;另一方面反映出细胞膜通透性增大导致细胞内蛋白质流失到细胞外,这与“2.3”节的结果趋势一致。
2.5 Lm细胞死亡的测定
cFDA/PI双染色结果见图6,cFDA是一种渗透性染料,自身不发荧光,穿过细胞膜后被细胞内的酯酶水解,释放出发绿色荧光的羧基荧光素cF,非渗透性的cF只能保留在具有完整细胞膜的活细胞中,死细胞和细胞膜破损的受伤细胞不能被染色[8]。PI不能穿过完整的活细胞膜,只能进入受伤细胞和死细胞破损的细胞膜并与核酸结合发红色荧光,因此PI染色可以反映出细胞膜破损和细胞死亡的程度[9]。采用cFDA/PI双染色结合流式细胞仪可以检测菌体活细胞、受伤细胞和死细胞。流式细胞仪FL1-FL3散点图显示4类不同的细胞,即cFDA+/PI-的活细胞(Q1-LR)、cFDA+/PI+的受伤细胞(Q1-UR)、cFDA-/PI+的死细胞(Q1-UL)以及cFDA-/PI-的细胞碎片(Q1-LL)[10]。试验结果表明,正常Lm大多数细胞处于活细胞状态,所占比例为93.1%;MIC肉桂醛处理6 h后,活细胞比例降低到2.6%,细胞受伤率和死亡率分别升高到73.7%和21.9%,大多数细胞处于膜损伤状态,但是细胞内的酯酶仍然具有活性;MBC肉桂醛处理6 h后,细胞死亡率升高到95.6%,大多数细胞处于死亡状态。
3 结论与讨论
本试验研究了肉桂醛对Lm的抑制作用。肉桂醛对Lm的MIC和MBC分别为150、300 μg/mL。扫描电镜结果显示,肉桂醛处理后Lm出现变形、膨大的现象,细胞表面并未观察到明显的凹陷和穿孔。 肉桂醛处理1 h和2 h 后Lm细胞外的葡萄糖和核酸含量明显高于空白对照,说明肉桂醛处理使得Lm细胞通透性增大,细胞内的物质向细胞外流失。SDS-PAGE电泳分析结果表明,肉桂醛处理后Lm细胞蛋白含量降低,肉桂醛抑制了分子量在 35~170 ku之间蛋白的表达。cFDA/PI双染色结合流式细胞仪检测结果显示,肉桂醛处理后的Lm活细胞比例降低,细胞受伤率和死亡率升高,MIC浓度的肉桂醛处理6 h后,细胞受伤率升高到73.7%,MBC浓度的肉桂醛处理6 h后,细胞死亡率达到到95.6%。综上所述,肉桂醛可对Lm的细胞形态、细胞膜渗透性造成损伤,并且抑制其大分子量蛋白质的合成。有关肉桂醛抑制Lm的特异性靶标和分子机制还有待进一步研究。
参考文献:
[1]Hain T,Chatterjee S S,Ghai R,et al. Pathogenomics of Listeria spp.[J]. International Journal of Medical Microbiology,2007,297(7/8):541-557.
[2]Buchanan R L,Gorris L G M,Hayman M M,et al. A review of Listeria monocytogenes:an update on outbreaks,virulence,dose-response,ecology,and risk assessments[J]. Food Control,2017,75:1-13.
[3]Burt S. Essential oils:their antibacterial properties and potential applications in foods—a review[J]. International Journal of Food Microbiology,2004,94(3):223-253.
[4]Sanla-Ead N,Jangchud A,Chonhenchob V,et al. Antimicrobial activity of cinnamaldehyde and eugenol and their activity after incorporation into cellulose-based packaging films[J]. Packaging Technology and Science,2012,25(1):7-17.
[5]Clemente I,Aznar M,Silva F,et al. Antimicrobial properties and mode of action of mustard and cinnamon essential oils and their combination against foodborne bacteria[J]. Innovative Food Science and Emerging Technologies,2016,36:26-33.
[6]王 帆,杨静东,王春梅,等. 复配植物源杀菌剂的开发研究[J]. 江西农业学报,2010,22(2):87-89.
[7]陆海霞,黄小鸣,朱军莉. 超高压对单增李斯特菌细胞膜的损伤和致死机理[J]. 微生物学报,2014,54(7):746-753.
[8]Ananta E,Heinz V,Knorr D. Assessment of high pressure induced damage on Lactobacillus rhamnosus GG by flow cytometry[J]. Food Microbiology,2004,21:567-577.
[9]Amor K B,Breeuwer P,Verbaarschot P,et al. Multiparametric flow cytometry and cell sorting for the assessment of viable,injured,and dead Bifidobacterium cells during bile salt stress[J]. Applied and Environmental Microbiology,2002,68(11):5209-5216.
[10]葉 雷,陈庆森,阎亚丽,等. 流式细胞术快速检测直投式发酵剂菌体活力[J]. 食品科学,2014,35(10):139-144.
猜你喜欢
现代农业科技(2017年14期)2017-09-06
商情(2017年14期)2017-06-09
商业研究(2017年2期)2017-03-21
山东农业科学(2017年2期)2017-03-15
农业科技与装备(2014年9期)2014-12-26
