
有瓣蝇类触角感受器形态与功能研究进展
2020-05-11 08:59庞秀楠刘贤慧刘根廷李心钰王企珂
环境昆虫学报 2020年2期
庞秀楠,刘贤慧,刘根廷,李心钰,王企珂,张 东*
(1.北京林业大学生态与自然保护学院,北京 100083;2.美国加州大学戴维斯分校,戴维斯 95616;3.墨尔本大学生物科学学院,维多利亚 3010)
有瓣蝇类Calyptratae隶属于昆虫纲Insecta四大超适应辐射类群之一的双翅目Diptera,是短角亚目Brachycera环裂次目Cyclorrhapha有缝组Schizophora的重要组成类群,其主要特征为触角鞭节较短,触角梗节背外侧具一纵贯全长的裂缝等(范滋德,1992;范滋德,1997;薛万琦和赵建铭,1998;Papeetal.,2011;Wiegmannetal.,2011)。最早的有瓣蝇类起源于6 500万年前(Wiegmannetal.,2011),目前已记述的有瓣蝇类物种数量为23 000余种,约占双翅目昆虫物种数量的20%,是双翅目物种多样性最丰富类群的之一(Papeetal.,2011)。有瓣蝇类包括虱蝇总科Hippoboscoidea、蝇总科Muscoidea和狂蝇总科Oestroidea共3个总科,涵盖舌蝇科Glossinidae、蝇科Muscidae、丽蝇科Calliphoridae、麻蝇科Sarcophagidae等18个科,分布遍及除南极大陆外的各个动物地理界。有瓣蝇类生态类型丰富:一些尸食、粪食性类群(如厩腐蝇Muscinastabulans)在生态系统中扮演着分解者的重要角色;一些专性寄生类群(如羊狂蝇Oestrusovis、黑须污蝇Wohlfahrtiamagnifica)能够造成人与动物的蝇蛆病;一些植食性类群(如地种蝇Deliaspp.、球果花蝇Strobilomyiaspp.)能够影响甚至破坏植物的生长;一些吸血性类群(如舌蝇Glossinaspp.、厩螫蝇Stomoxyscalcitrans)是传播多种疾病的重要媒介生物;一些捕食性类群(如秽蝇Coenosiaspp.、溜蝇Lispespp.)是维系食物链的关键类群(范滋德,2008;Kuttyetal.,2010)。快速的适应辐射、极高的物种多样性、广泛的分布和丰富的生态类型使得有瓣蝇类成为开展昆虫适应演化研究的理想类群。
触角是昆虫感知外界环境的重要感觉器官,在寻找食源或寄主、识别异性、定位产卵位置等生活史关键环节中均起到作用,是形态学和生理学领域长期以来的研究热点(Schneider,1964;Zacharuk,1980;Wiegmannetal.,2011;Elgaretal.,2018;Wangetal.,2018;Pangetal.,2020)。昆虫通过其体表分布的大量特化感受结构来获取外界信息,这些结构统称为感受器(Snodgrass,1926)。例如,在昆虫产生嗅觉反应的过程中,气流中的化学信号分子通过与触角上分布的嗅觉感受器直接接触而引起感受器内的嗅觉神经产生神经冲动(Greenfield,2002),进而引发大脑的嗅觉反应。对昆虫触角感受器的研究能够揭示昆虫的感觉机制,探究昆虫的行为如何受到外界因子的影响,从而深入了解生物与环境的相互作用关系。本文综合有瓣蝇类触角感受器的现有研究,统一了之前研究中有瓣蝇类触角上各类感受结构的不同名词,并结合其它昆虫类群的相关研究,综述了各类感受器形态与功能等方面的研究进展,旨在为进一步开展昆虫触角感受器结构的适应演化研究提供参考。
1 有瓣蝇类触角形态结构概况
昆虫为了有效感知不同的外界信号,演化出了多样的触角形态,包括丝状、羽状、鳃状、念珠状、具芒状等10余种类型(彩万志等,2011)。有瓣蝇类触角属于具芒状触角,位于头部前侧复眼中间(图1A),由柄节(Scape,Sc)、梗节(Pedicel,Pd)和具触角芒(Arista,Ar)的鞭节(Funiculus,Fn)共3节组成(图1B,图1C)(Kirk-Spriggs and Sinclair,2017)。一般柄节最短,梗节长度中等,鞭节长度最长(图1D),但有瓣蝇类中一些类群因生存环境不同,触角各节发生适应性演化,外形存在明显差别(Elgaretal.,2018):如黑角胃蝇Gasterophilusnigricornis梗节缩短近盘状(Zhangetal.,2012a)(图1E),紫鼻狂蝇Rhinoestruspurpureus触角鞭节缩短(Liuetal.,2015b)(图1F),纹皮蝇Hypodermalineatum鞭节缩短被梗节半包裹(Lietal.,2015)(图1G),壮小头皮蝇Portschinskiamagnifica鞭节完全被梗节包裹(Zhangetal.,2012b)(图1H,图1I)。鞭节横截面通常为三角形,分为3个面:前背面(Anterodorsal surface,Ad)、背侧面(Dorsolateral margin,Dl)和后腹面(Posteroventral surface,Pv)(图1B,图1C)。触角芒着生于鞭节背侧面,一般分为3节,前2节较短,第3节细长。有瓣蝇类的触角芒根据形态特征可分为:裸、具毳毛、具短纤毛、栉状、羽状(图1J)。
有瓣蝇类的具芒触角长度短却具有极其敏锐的嗅觉功能,是因为其表面分布了大量的感受器。触角感受器通常由神经元(Neuron)、鞘原细胞(Thecogen cell)、毛原细胞(Trichochen cell)、膜原细胞(Tormogen cell)、感器液腔(Sensillum-lymph cavity)、表皮细胞(Epidermal cell)和最外部的角质层(Cuticle)组成(图2)(Kelling,2001;王荫长,2004)。触角感受器在有瓣蝇类获取机械震动、温湿度、化学信号等外界信息的过程中起直接作用。有瓣蝇类因涵盖病媒、天敌、传粉昆虫以及农林害虫等与人类关系密切的类群,对其触角感受器的形态与功能研究均取得了良好的进展。

图1 有瓣蝇类触角结构Fig.1 Features on the antenna of Calyptratae注:A-H为扫描电子显微镜(SEM)图像。A,内蒙古溜蝇Lispe neimongola头部(Zhang et al.,2013b),示有瓣蝇类触角着生位置;B,叉丽蝇Triceratopyga calliphoroides触角前背面(Zhang et al.,2014);C,叉丽蝇触角背侧面和后腹面(Zhang et al.,2014);D,灰地种蝇Delia platura触角(Wang et al.,2014b);E,黑角胃蝇Gasterophilus nigricornis触角,示梗节缩短近盘状(Zhang et al.,2012a);F,紫鼻狂蝇Rhinoestrus purpureus触角,示鞭节缩短(Liu et al.,2015b);G,纹皮蝇Hypoderma lineatum触角,示鞭节缩短被梗节半包裹(Li et al.,2015);H,壮小头皮蝇Portschinskia magnifica触角鞭节(Zhang et al.,2012b);I,壮小头皮蝇触角体视显微镜图像,示鞭节缩短被梗节包裹(Zhang et al.,2012b);J,有瓣蝇类触角触角芒类型,J1-J5分为裸、具毳毛、具短纤毛、栉状、羽状,H6为一种舌蝇Glossina tachinoides的触角,示触角芒上侧具羽状分枝的小毛(范滋德,1992)。比例尺:A=500 μm,B-I=200 μm。Ad,前背面;Ar,触角芒;Dl,背侧面;Fn,鞭节;Pd,梗节;Pv,后腹面;Sc,柄节。Note:SEM micrographs of A,head of Lispe neimongola,showing the position of the antenna of Calyptratae (Zhang et al.,2013b);B,antenna of Triceratopyga calliphoroides,showing the anterodorsal surface (Zhang et al.,2014);C,antenna of Triceratopyga calliphoroides,showing the dorsolateral margin and posteroventral surface (Zhang et al.,2014);D,antenna of Delia platura (Wang et al.,2014b);E,antenna of Gasterophilus nigricornis,showing the shortened antennal pedicel (Zhang et al.,2012a);F,antenna of Rhinoestrus purpureus,showing the shortened antennal funiculus (Liu et al.,2015b);G,antenna of Hypoderma lineatum,showing the shortened antennal funiculus half-covered by antennal pedicel (Li et al.,2015);H,antennal funiculus of Portschinskia magnifica (Zhang et al.,2012b);I,Stereo microscope photograph of the antenna of Portschinskia magnifica,showing the shortened antennal funiculus fully covered by antennal pedicel (Zhang et al.,2012b);J,Types of arista on Calyptratae antenna:bare,pubescent,short ciliated,pectinate,plumose,and J6 shows the antenna of Glossina tachinoides (Fan,1992).Scale bars:A=500 μm,B-I=200 μm.Ad,Anterodorsal surface;Ar,Arista;Dl,Dorsolateral margin;Fn,Funiculus;Pd,Pedicel;Pv,Posteroventral surface;Sc,Scape.

图2 昆虫触角感受器结构(仿Larter et al.,2016)Fig.2 Structure of insect antennal sensilla (Adapted from Larter et al.,2016)注:每个触角感受器通常由2个神经元、1个鞘原细胞、1个毛原细胞、1个膜原细胞、感器液腔、表皮细胞和最外部的角质层组成,通过表皮细胞与相邻感受器相间隔。C,角质层;E,表皮细胞;N,神经元;SLC,感器液腔;Th,鞘原细胞;To,膜原细胞;Tr,毛原细胞。Note:Each antennal sensilla usually consists of two neurons,one thecogen cell,one trichochen cell,one tormogen cell,sensillum-lymph cavity,epidermal cells,and the outermost cuticle.They separated from each other by epidermal cells.C,Cuticle;E,Epidermal cell;N,Neuron;SLC,Sensillum-lymph cavity;Th,Thecogen cell;To,Tormogen cell;Tr,Trichochen cell.
2 有瓣蝇类触角的形态与功能研究进展
Hauser(1880)对昆虫嗅觉的研究中,最早详细描述了直翅目、脉翅目等6个目昆虫的触角解剖结构,证实了昆虫的嗅觉器官存在于触角上。Smith和Lefroy(1919)利用石蜡切片技术,对双翅目多个科的物种进行了触角解剖研究,并通过显微镜观察,发现了多种在其他昆虫类群中极为罕见的,有瓣蝇类所特有的复杂内部结构,包括感觉窝和感觉囊这类感受器复合体,由此引发了昆虫形态学家的长期关注。随着电子显微镜技术的发展,应用扫描电子显微镜(Scanning Electron Microscope,SEM)和透射电子显微镜(Transmission Electron Microscope,TEM)的研究将触角感受器形态的研究推进到微米级的超显微结构水平。Slifer和Sekhon(1964)首次使用了透射电子显微镜观察了银口红麻蝇Sarcophagaargyrostoma的触角表面和感觉窝内部的感受器,Greenberg(1970)首次使用扫描电镜开展了有瓣蝇类触角感受器超显微结构的详细研究,Zacharuk(1980)总结了当时采用扫描电镜和透射电镜技术对昆虫的触角感受器形态开展研究的成果。至今,在舌蝇科(Isaacetal.,2015)、虱蝇科(Zhangetal.,2015;Andreanietal.,2019)、厕蝇科(任宏伟等,2011;Wangetal.,2012;Zhangetal.,2013c)、蝇科(Beenetal.,1988;Sukontasonetal.,2004;Sukontasonetal.,2007;Smallegangeetal.,2008;Tangtrakulwanichetal.,2011;Zhangetal.,2013b;Wangetal.,2014a;Carricoetal.,2015;Horeetal.,2018)、花蝇科(Hondaetal.,1983;Ross and Anderson,1987;Ross and Anderson,1991;Ross,1992;Wangetal.,2014b)、粪蝇科(Liuetal.,2016)、墨丽蝇科(Caetanoetal.,2018)、麻蝇科(Sukontasonetal.,2004;Abouziedetal.,2008b;Pezzietal.,2016)、狂蝇科(Hunter and Adserballe,1996;Poddigheetal.,2010;Zhangetal.,2012a;Zhangetal.,2012b;Lietal.,2015;Liuetal.,2015b;Zhangetal.,2016)、寄蝇科(Giangiulianietal.,1994;Rahaletal.,1996;Abouziedetal.,2008a;Liuetal.,2013;Rohetal.,2020)和丽蝇科(Sukontasonetal.,2004;Sukontasonetal.,2007;Sukontasonetal.,2008;Setzuetal.,2011;Zhangetal.,2013a;Zhangetal.,2014;Horeetal.,2017;Yanetal.,2019)这些有瓣蝇类科级阶元中,触角及触角感受器的形态得以深入研究。
对有瓣蝇类触角形态的认知促进了有瓣蝇类触角功能的研究。Forel(1900)通过比较丝光绿蝇Luciliasericata触角各节的外部形态,推测蝇类触角的柄节和梗节没有嗅觉感受功能,而相对较大的鞭节才是产生嗅觉的部位,甚至推测嗅觉功能与鞭节表面的感觉毛(即感受器)密切相关。20世纪中期电生理学相关技术的进步,为昆虫触角感受器功能的研究带来了突破性发展。Schneider和Hecker(1956)利用电生理学方法首次成功记录到触角电位图(Electroantennography,EAG)。Morita和Yamashita(1961)对Schneider的触角电位技术进行改进,研发了能够观察到单细胞脉冲电位变化的单细胞记录技术。Boeckh等(1963)用钨丝电极插入毛形感受器基部,成功获得了关于毛形感受器的单细胞记录。Schneider(1964)对昆虫触角感受器的形态、功能、定位及神经传导方面做了详细的综述。至今,应用这些技术的研究已经逐渐在有瓣蝇类中得到开展(诸葛洪祥等,1999;Wasserman and Itagaki,2003;Tangtra-kulwanichetal.,2011)。
有瓣蝇类触角表面分布有多种感受结构(Shanbhagetal.,1995;Shanbhagetal.,1999;Shanbhagetal.,2000),主要类型如下:机械感受器(Mechanoreceptor,Mr)、毛形感受器(Trichoid sensilla,Tr)、锥形感受器(Basiconic sensilla,Ba)、腔锥形感受器(Coeloconic sensilla,Co)、棒状感受器(Clavate sensilla,Cl),此外还在部分物种中发现了板形感受器(Plaque,Pl)(Greenberg,1970)、钟形感受器(Pedicellar button,PB)(McAlpine,2008)和耳形感受器(Auriculate sensilla,Au)(Setzuetal.,2011)。部分感受器聚集分布于感觉窝(Sensory pit,SP)或感觉囊(Sacculus,Sa)内。
2.1 机械感受器
机械感受器(Mr)是一类常见的感受器,几乎在有瓣蝇类全身都有分布(薛万琦和赵建铭,1998)。有瓣蝇类触角上的机械感受器主要分布于柄节和梗节,数目从几个到十几个。机械感受器一般表面具纵沟,呈长鬃状,基部着生于1个突起的光滑基座中,基座周围通常具1簇微毛(图3A)(Wangetal.,2014a;Pezzietal.,2016),这类基座被认为是机械感受器的标志特征(Lietal.,2014)。机械感受器能够感受接触物体的压力,外界空气、水或介质产生的振动,肌肉活动产生的张力,地球引力等机械刺激(王荫长,2004;Klowden,2007)。
2.2 板形感受器
板形感受器(Pl)最初由Greenberg(1970)在丽蝇科的丝光绿蝇中发现,在丽蝇科和麻蝇科中广泛分布,而对于蝇科,仅于部分物种(Greenberg and Ash,1972)中存在。板形感受器主要分布于触角梗节背面(图3B),数目从几个到十几个不等。板形感受器在有瓣蝇类不同类群间形态差异较大,有些呈扁平的盖状,基部具毛簇,端部急尖(图3C),有些呈膨胀的球根状(图3D),可作为分类依据(Greenberg,1970;Greenberg and Ash,1972;Sukontasonetal.,2004;Zhangetal.,2013a;Zhangetal.,2013c;Wangetal.,2014b;Liuetal.,2016)。Sukontason等(2004)对蝇类触角的比较形态学研究认为其可能具有机械或者化学感受功能。
2.3 钟形感受器
钟形感受器(PB)广泛分布于昆虫外骨骼上,在不同类群、不同部位上具有不同的形态结构及功能(Chapman,1971;McIver,1975)。有瓣蝇类触角的钟形感受器着生于梗节与鞭节连接处的凹陷内(图3E),因其着生位置隐蔽,直到McAlpine(2008)通过对触角梗节和鞭节的拆分才得以发现。有瓣蝇类每个触角一般仅有1个钟形感受器,其边缘明显,中间呈圆顶状的凸起,通常表面光滑,无孔或具孔(图3F)。此后McAlpine(2011)又发现此类感受器广泛存在于环裂亚目各类群中,甚至发现部分物种的每个触角上具有2个钟形感受器。
McAlpine(2011)认为钟形感受器属于压力感受器,能感受身体内外部的压力变化,行使本体感觉功能,其隐蔽的位置可能是一种保护策略。在膜翅目的部分物种中发现钟形感受器表面具孔,表明其可能为化学感受器(Dietz and Humphreys,1971;Ochiengetal.,2000)。在丝光绿蝇(Zhangetal.,2013a)和小黄粪蝇Scathophagastercoraria(Liuetal.,2016)中也发现具微孔的钟形感受器(图3F),说明化学气味分子有可能通过微孔进入其内部,从而行使化学感受功能,因此有瓣蝇类钟形感受器的功能还有待进一步探究。
2.4 毛形感受器
毛形感受器(Tr)在除虱蝇科物种外其他已研究的有瓣蝇类的触角上均有分布(Zhangetal.,2015),是有瓣蝇类触角感受器中分布最广且长度最长的感受器。毛形感受器主要分布于触角鞭节的中部和端部(图4O),其长度从十几μm到几十μm,一些较大的毛形感受器在光学显微镜下就可以观察到。毛形感受器一般从1个较明显隆起的光滑基部发生,紧密地嵌在表皮形成的槽中,向上逐渐变细,末端一般较尖(图4A),但也有些具有钝的末端(Smallegangeetal.,2008)。毛形感受器的基部横切显示其内部具有1或2个神经树突(图4B),处于感受器内腔的淋巴液中,而其近端部的横切则显示神经树突逐渐分为细枝(图4C)(Kelling,2001;Klowden,2007;刘贤慧等,2015a)。有瓣蝇类触角上毛形感受器的数量因物种而异,有的仅有几十个(Porter,1998),有的达到几百个(Ross,1992)。
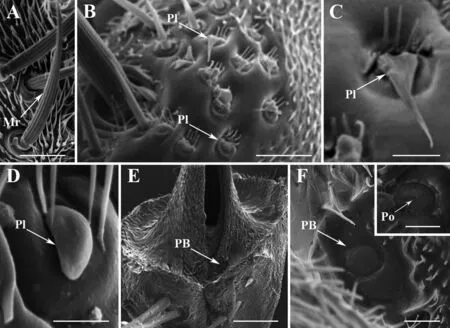
图3 有瓣蝇类触角梗节感受器类型Fig.3 Types of antennal sensilla on antennal pedicel of Calyptratae注:A-F为扫描电子显微镜(SEM)图像。A,刺足齿股蝇Hydrotaea armipes机械感受器(Wang et al.,2014a);B,丝光绿蝇Lucilia sericata触角梗节,示板形感受器分布位置(Zhang et al.,2013a);C,丝光绿蝇板形感受器(Zhang et al.,2013a);D,灰地种蝇Delia platura板形感受器(Wang et al.,2014b);E,蓝黑栉寄蝇Pales pavida触角梗节,示钟形感受器分布位置(Liu et al.,2013);F,蓝黑栉寄蝇钟形感受器(Liu et al.,2013),框内为小黄粪蝇Scathophaga stercoraria具孔的钟形感受器(Liu et al.,2016)。比例尺:A、B=20 μm,C、D=5 μm,E=50 μm,F=5 μm(框内5 μm)。Mr,机械感受器;PB,钟形感受器;Pl,板形感受器;Po,微孔。Note:SEM micrographs of A,mechanoreceptor of Hydrotaea armipes (Wang et al.,2014a);B,antennal pedicel of Lucilia sericata,showing the position of plaques (Zhang et al.,2013a);C,plaque of Lucilia sericata (Zhang et al.,2013a);D,plaque of Delia platura (Wang et al.,2014b);E,antennal pedicel of Pales pavida,showing the position of pedicellar button (Liu et al.,2013);F,pedicellar button of Pales pavida (Liu et al.,2013),and pored pedicellar button of Scathophaga stercoraria (in box) (Liu et al.,2016).Scale bars:A,B=20 μm,C,D=5 μm,E=50 μm,F=5 μm (5 μm in box).Mr,Mechanoreceptor;PB,Pedicellar button;Pl,Plaque;Po,Pore.

图4 有瓣蝇类触角鞭节感受器类型Fig.4 Types of antennal sensilla on antennal funiculus of Calyptratae注:小黄粪蝇Scathophaga stercoraria 毛形感受器(A),锥形感受器(D),腔锥形感受器(G),棒状感受器(K)和感觉窝(M)扫描电子显微镜(SEM)图像(Liu et al.,2016);丝光绿蝇Lucilia sericata触角毛形感受器(B),锥形感受器(E)和腔锥形感受器(H)基部横切透射电子显微镜(TEM)图像,箭头示感受器神经树突(Liu et al.,2015a);丝光绿蝇触角毛形感受器(C),锥形感受器(F)和腔锥形感受器(I)近端部横切TEM图像,箭头示神经树突分枝(Liu et al.,2015a);黑角胃蝇Gasterophilus nigricornis 触角芒腔锥形感受器( J)和耳形感受器(L)SEM图像(Zhang et al.,2012a);夏厕蝇Fannia canicularis触角感觉窝和感觉囊(N)纵切光学显微镜(LM)图像(Zhang et al.,2013c);黑腹胃蝇Gasterophilus pecorum触角(O)形态模式图,示四种触角感受器分布位置(Zhang et al.,2016)。比例尺:A=5 μm,B-L=2 μm,M=10 μm,N=50 μm。Au,耳形感受器;Ba,锥形感受器;Cl,棒状感受器;Co,腔锥形感受器;Pit,感觉窝;Sacculus,感觉囊;Tr,毛形感受器。Note:SEM micrographs of trichoid sensilla(A),basiconic sensilla(D),coeloconic sensilla(G),clavate sensilla(K),and sensory pit(M) of Scathophaga stercoraria (Liu et al.,2016).TEM micrographs of Lucilia sericata,showing cross sections through the base of trichoid sensilla(B),basiconic sensilla(E),and coeloconic sensilla(H),arrows point to the nerve dendrites (Liu et al.,2015a).TEM micrographs of Lucilia sericata,showing cross sections through the tip of trichoid sensilla(C),basiconic sensilla(F),and coeloconic sensilla(I),arrow points to the nerve dendritic branches (Liu et al.,2015a).SEM micrographs of coeloconic sensilla(J) without longitudinal grooves,and auriculate sensilla(L) of Gasterophilus nigricornis (Zhang et al.,2012a).Light microscope photograph of Fannia canicularis,showing longitudinal section through sensory pits and sacculus(N) (Zhang et al.,2013c).Distribution of four types of sensilla(O) on antennal funiculus of Gasterophilus pecorum (Zhang et al.,2016).Scale bars:A=5 μm,B-L=2 μm,M=10 μm,N=50 μm.Au,Auriculate sensilla;Ba,Basiconic sensilla;Cl,Clavate sensilla;Co,Coeloconic sensilla;Pit,Sensory pit;Tr,Trichoid sensilla.
毛形感受器是研究最多的感受器类型。早期的研究根据毛形感受器表面分布微孔的事实,推断其应该属于化学感受器(Clyneetal.,1997)。随后的电生理学研究证明了毛形感受器对性外激素的敏感性,从而确认了其化学感受功能,并认为毛形感受器对寻找异性具有关键作用(Clyneetal.,1999)。气味受体蛋白(Odorant binding protein)对昆虫产生嗅觉反应不可或缺(Sachse and Krieger,2011),最近的研究表明毛形感受器内存在气味受体蛋白(Gomezdiazetal.,2018),更加明确了毛形感受器的化学感受功能。
2.5 锥形感受器
锥形感受器(Ba)在已研究的全部有瓣蝇类的触角上都有分布,是触角上数量和分布密度仅次于毛形感受器的类型。锥形感受器的分布没有明显的特异性,在触角表面均匀分布或集中分布于感觉窝或感觉囊中(图4O)。锥形感受器呈从底部向端部逐渐变细的锥状,有时沿鞭节的长轴方向具不同程度的弯曲,外壁密布微孔,密度较毛形感受器更大(图4D)。锥形感受器内基部具有2或3个神经树突(图4E),向端部产生大量分枝,甚至可充满整个感受器内腔(图4F)(Kelling,2001;Klowden,2007;刘贤慧等,2015a)。一般同一物种的锥形感受器形态多样,根据不同的形态结构,常分为2到3种亚型(Wangetal.,2012;Zhangetal.,2012a),部分有瓣蝇类的锥形感受器仅具有1种类型(Zhangetal.,2012b)。
根据有瓣蝇类触角锥形感受器表面具孔的特征和透射电子显微镜的拍摄结果,以及Clyne等(1997)、Riesgo-Escovar等(1997)使用电生理学方法观察到锥形感受器对气味物质能够产生反应的现象,加之最近的研究也表明锥形感受器内存在气味受体蛋白(Gomezdiazetal.,2018),目前普遍认为锥形感受器具有化学感受功能。这个结论与鳞翅目(Shields and Hildebrand,1999)、鞘翅目(Lopesetal.,2002)、半翅目(Broeckling and Salom,2003)等类群昆虫中的研究类似。
2.6 腔锥形感受器
腔锥形感受器(Co)有时也被称为具沟感受器(Grooved sensilla)。目前除蝇科溜蝇属的内蒙古溜蝇Lispeneimongola外,腔锥形感受器在已有研究的有瓣蝇类的触角上均有分布(Zhangetal.,2013b)。腔锥形感受器数量较少,均匀分布或集中分布于鞭节表面(图4O)。腔锥形感受器通常着生于1个表皮凹陷中,长度短于微毛,表面有数道沿轴方向的纵沟(图4G)(Shanbhagetal.,1999)。腔锥形感受器具有双层壁结构,外层壁厚于内层壁,感受器基部具有2个神经树突(图4H),向端部逐渐分枝(图4I)(Kelling,2001;Klowden,2007;刘贤慧等,2015a)。腔锥形感受器的形态变化较大,在不同物种间甚至同一物种内可能存在不同的形态,为方便研究,常将其分为几种亚型(Fernandesetal.,2004;Sukontasonetal.,2004)。除了触角鞭节,有些物种的触角芒上也具有腔锥形感受器,不同于触角鞭节上的类型,这些腔锥形感受器表面通常光滑而无纵沟(图4J)(Zhangetal.,2012a;Zhangetal.,2016)。
Schneider和Steinbrecht(1968)用电生理学的实验方法证明了腔锥形感受器具有感觉CO2浓度、温度和湿度的功能,Ochieng等(2000)同样提出腔锥形感受器为温湿度感受器。也有一些学者认为该感受器可能为嗅觉感受器(Olson and Andow,1993;Shanbhagetal.,1995;Clyneetal.,1997),且有研究表明腔锥形感受器内存在气味受体蛋白(Gomezdiazetal.,2018),因此有瓣蝇类腔锥形感受器的功能还有待进一步探究。
2.7 棒状感受器
棒状感受器(Cl)在已有研究的有瓣蝇类的触角上均有分布。棒状感受器主要分布于触角鞭节的基部(图4O),着生于略深的表皮凹陷或位于感觉窝内,顶端膨大,形状类似棒球棒(图4K)(Slifer,1954),长度与触角表面的微毛相似。
棒状感受器表面分布微孔,有研究根据部分物种中棒状感受器表面微孔较少的特征,推测其功能很可能是感知CO2(Lewis,1971),但也有研究推测其主要作用是感受温度和湿度(Ross and Anderson,1987)。
2.8 耳形感受器
耳形感受器(Au)在有瓣蝇类触角感受器中相对少见,目前仅在丽蝇(Setzuetal.,2011)和狂蝇(Zhangetal.,2012a;Zhangetal.,2016)物种中有所发现。Setzu(2011)等人在新陆原伏蝇Protophormiaterraenovae中首次提到耳形感受器,随后,在黑角胃蝇、红尾胃蝇Gasterophilushaemorrhoidalis、肠胃蝇Gasterophilusintestinalis、鼻胃蝇Gasterophilusnasalis和黑腹胃蝇Gasterophiluspecorum中也发现了这类感受器(Zhangetal.,2012a;Zhangetal.,2016)。耳形感受器均匀分布于触角鞭节表面或位于感觉窝内,形状类似耳形或勺形,表面具大量微孔,中部较宽而扁平,端部急尖,长度略长于微毛(图4L)。
Shields和Hildebrand(1999,2001)切片研究烟草天蛾Manducasexta的耳形感受器显示其为单层壁,表面密布微孔,推测其具有嗅觉感受功能。目前在有瓣蝇类中发现的耳形感受器表面同样密布微孔,显示其可能为一种嗅觉感受器。
2.9 感觉窝
除上述单个分布的感受器以外,在有瓣蝇类触角表面还存在一些称为感觉窝(SP)的表皮凹陷结构(图4M)。感觉窝呈单室凹陷,内部为聚集在一起的多个同种类型的感受器(Zhangetal.,2014)。感觉窝内感受器的数量可从几个至几十个不等(Zhangetal.,2014;Liuetal.,2016)。不同类群有瓣蝇类的触角上,感觉窝的数量差别较大,开口大小的变化也很大(Zhangetal.,2013c;Zhangetal.,2014;Liuetal.,2016;Pezzietal.,2016)。
感觉窝的感受功能是通过其内部的感受器来完成的。有时在同一个触角上可能看到几种不同的感觉窝,其中通常具有锥形感受器、腔锥形感受器、棒状感受器、耳形感受器等(Sukontasonetal.,2004;Zhangetal.,2012a;Zhangetal.,2016)。Wasserman和Itagaki(2003)使用NADPH-硫辛酰胺脱氢酶的组织化学方法,通过观察感觉细胞染色的方式证明了感觉窝具有嗅觉功能。
2.10 感觉囊
通过对有瓣蝇类触角进行纵切,发现有瓣蝇类触角内存在复杂的多室凹陷结构,这些结构称为感觉囊(Sa)(图4N)。与感觉窝不同,感觉囊内部有多种不同类型的感受器(Zhangetal.,2013c)。温度感受器、湿度感受器与化学感受器在感觉囊中同时存在,表明感觉囊为一类感受器复合体,其内部各种不同的感受器同时执行各自的功能(Zhangetal.,2014;刘贤慧等,2015a)。感觉囊能够使气味分子聚集,从而提高化学感受器的灵敏度;能够有效保护其内部的感受器免于机械损伤;能够为这些感受器提供相对稳定的微环境,减缓感受器的水分散失(Ross,1992;Shanbhagetal.,1995;Hunter and Adserballe,1996;Sukontasonetal.,2004;Zhangetal.,2012a;Zhangetal.,2013c)。
3 问题与展望
虽然Schneider(1964)已在关于昆虫触角的综述中对感受器类型及形态做出了较为全面的总结,但因有瓣蝇类物种数量大,生态类型多样,触角上各种类型的感受器间存在较大的变化,导致现有研究对触角感受器的类型定义和形态描述之间存在诸多差异、混乱和歧义。仅利用感受器的外部形态特征很难对感受器进行准确区分,现有研究中存在毛形感受器与锥形感受器混淆的现象(Fernandesetal.,2004;Sukontasonetal.,2004)。不同的研究角度采用不同的定义方式,从形态特征出发可将触角感受器分为毛形感受器、锥形感受器、腔锥形感受器等;从感受器的表面结构出发可将感受器分为无气孔类,壁具孔类(此类又可细分为单壁型和双壁型)和顶端具孔类(Altneretal.,1977;Altneretal.,1983);而从感受器的功能出发可以将感受器分为机械感受器、化学感受器、温度感受器和湿度感受器(余海忠,2007)。透射电子显微镜、激光共聚焦显微镜(Laser Scanning Confocal Microscopy,LSCM)能够对触角切片样品进行精细观察,已逐渐在有瓣蝇类触角感受器的研究中得到应用(刘贤慧等,2015)。触角电位技术(EAG)是通过测量昆虫触角对特定物质刺激产生反应的电生理学方法(田厚军等,2011)。触角电位技术对实验操作水平的要求较高,目前对相关研究仅限于毛形感受器、锥形感受器和腔锥形感受器3种(Clyneetal.,1997;Van Naters and Carlson,2007),存在大量研究空白。将触角感受器的外部形态与内部结构相结合,并参考触角感受器电生理学研究,从而确定各个触角感受器的功能,是更加准确定义不同类型触角感受器的基础。
另一方面,长期以来对有瓣蝇类触角感受器的研究主要集中于蝇科、丽蝇科、花蝇科等与人类关系密切的类群。对寄生性有瓣蝇类的研究多为狂蝇、皮蝇、胃蝇等脊椎动物寄生蝇,而对寄蝇科这类以节肢动物为寄主的重要天敌昆虫的触角形态研究较为匮乏,目前仅有对Trichopodapennipes(Giangiulianietal.,1994)、稻苞虫赛寄蝇Pseudoperichaetanigrolineata(Rahaletal.,1996)、追寄蝇Exoristasp.(Abouzied,2008a)、蓝黑栉寄蝇Palespavida(Liuetal.,2013)和普通球腹寄蝇Gymnosomarotundatum(Rohetal.,2020)5个物种的研究。有瓣蝇类触角感受器类型丰富,在不同科之间甚至同一科不同属的不同物种间存在较大的形态差异,涉及不同类群和不同生态型大范围开展有瓣蝇类触角感受器的研究仍需加强。
近年来,随着显微CT(Micro-CT)和三维重建技术的发展,越来越多的先进手段应用到昆虫形态学研究领域。显微CT不仅已应用于昆虫的肌肉系统、神经系统、消化系统的三维结构重建(Geetal.,2012),在颚器、疣足等一些昆虫外部器官的三维结构重建中也取得了良好的效果(Faulwetteretal.,2013)。有瓣蝇类具芒触角的内部结构较为复杂,尤其是感觉囊和感觉窝的组成与相互关系较难通过传统的形态学手段研究。利用这些技术的优势进行有瓣蝇类触角的高精度三维结构重建,并结合各物种的生物学特征从形态学角度揭示触角的适应演化过程,将是下一阶段研究的重点之一。
致谢:在此由衷感谢对本研究提供长期支持的史军娜老师和张辉老师(北京林业大学公共分析测试中心)。
猜你喜欢
南京林业大学学报(自然科学版)(2022年3期)2022-11-29
汽车实用技术(2022年14期)2022-07-30
口腔医学(2021年10期)2021-12-02
新世纪智能(英语备考)(2021年11期)2021-03-08
昆明医科大学学报(2021年1期)2021-02-07
华人时刊(2020年19期)2021-01-14
中国现代医药杂志(2020年10期)2020-12-14
扬子江(2019年1期)2019-03-08
百科知识(2016年21期)2016-12-24
作文评点报·作文素材小学版(2016年9期)2016-03-18
