
不同饲养温度对草地贪夜蛾过冷却点和体液冰点的影响
2020-06-08 09:41谢殿杰张蕾程云霞
植物保护 2020年2期
关键词:草地贪夜蛾
谢殿杰 张蕾 程云霞


摘要 为明确不同温度条件下草地贪夜蛾耐寒能力的差异,本研究利用过冷却点仪对不同温度条件下饲养的草地贪夜蛾各虫态的过冷却点和体液冰点进行了测定,并进行了频次分布分析。结果显示:在不同温度(16、20、24、28、32、36℃)条件下饲养的草地贪夜蛾,各虫态过冷却点和体液冰点存在显著差异。其中,3龄幼虫、6龄幼虫以及2日龄蛹的过冷却点和体液冰点均为在16℃下饲养时最低,3龄幼虫分别为(-13.48±0.53)℃和(-8.97±0.35)℃;6龄幼虫分别为(-8.94±0.28)℃和(-4.99±0.28)℃;2日龄蛹分别为(-18.09±0.50)℃和(-12.17±0.37)℃。5日龄蛹过冷却点在16℃饲养时最低,为(-17.76±0.49)℃,体液冰点在32℃和16℃饲养时最低,分别为(-11.72±0.56)℃和 (-11.02±0.39)℃;成虫过冷却点在16℃饲养时最低,为(-17.87±0.55)℃,体液冰点在28℃和16℃饲养时最低,分别为(-11.23±0.61)℃和 (-10.98±0.44)℃。本研究结果表明低温对草地贪夜蛾的耐寒能力具有一定的驯化作用,同时适当的高温也可以提高草地贪夜蛾的耐寒能力。该研究结果有助于预测草地贪夜蛾在我国的越冬分布区域,对其监测预警及防控具有重要意义。
关键词 草地贪夜蛾; 饲养温度; 过冷却点; 体液冰点
中图分类号: S 435.132 文献标识码: A DOI: 10.16688/j.zwbh.2019654
Abstract In order to investigate the cold tolerance of Spodoptera frugiperda under different feeding temperatures, the supercooling point and freezing point were measured by the supercooling point meter, and the frequency distribution was analyzed. The results showed that there were significant differences in the supercooling and freezing points of each age at different feeding temperatures (16, 20, 24, 28, 32, 36℃). The supercooling points and freezing points of 3-instar, 6-instar and 2-day-old pupae were the lowest at 16℃, which were (-13.48±0.53)℃ and (-8.97±0.35)℃ for 3-instar, (-8.94±0.28)℃ and (-4.99±0.28)℃ for 6-instar, (-18.09±0.50)℃ and (-12.17±0.37)℃ for 2-day-old pupae, respectively. The supercooling point of 5-day-old pupae was the lowest at 16℃, which was (-17.76±0.49)℃, and the freezing point was the lowest at 32℃ and 16℃, which were (-11.72±0.56)℃ and (-11.02±0.39)℃, respectively. The adult supercooling point was the lowest at 16℃, which was (-17.87±0.55)℃, and the freezing point were the lowest at 28℃ and 16℃, which were (-11.23±0.61)℃ and (-10.98±0.44)℃, respectively. The results of this research indicated that low temperature had a certain role in acclimating the cold tolerance of the S. frugiperda, and the appropriate high temperature could also improve the cold tolerance. The results are helpful to predict the overwintering distribution of S. frugiperda in China, and are of great significance for its monitoring, early warning, prevention and control.
Key words Spodoptera frugiperda; feeding temperature; supercooling points; freezing points
草地貪夜蛾Spodoptera frugiperda (J.E.Smith)属鳞翅目Lepidoptera,夜蛾科Noctuidae,灰翅夜蛾属 Spodoptera,也称秋黏虫,原产于美洲热带和亚热带地区[1]。草地贪夜蛾具有寄主范围广,为害严重,繁殖能力强,适应性强等特点,在中、南美洲可周年发生[2-3]。成虫具有迁飞习性,在中、南美洲可向南迁飞至阿根廷,向北迁飞至加拿大[4-6]。2016年1月,草地贪夜蛾首次入侵非洲,并迅速蔓延到非洲大部分地区[7],以及部分亚洲国家[8]。2019年1月草地贪夜蛾首次入侵我国云南,随后迅速蔓延,截至目前,已经有25个省(市)受到草地贪夜蛾为害[9-12]。
温度是影响昆虫发育历期、存活率、繁殖力及分布范围的环境因素[13]。昆虫是一种变温动物,温度的变化会引起昆虫体内发生一系列生理生化反应,温度过高或过低都会对昆虫种群繁衍产生不利影响[14]。在长期的生物进化过程中,昆虫种群形成了对不利温度条件的适应机制[15]。昆虫对温度变化的响应分为抗寒能力和高温耐受能力,其中过冷却点和体液冰点是评价昆虫耐寒性的重要指标[16]。研究者对黏虫Mythimna separata、花绒寄甲Dastarcus helophoroides、梨小食心虫Grapholitha molesta、马铃薯麦蛾Phthorimaea operculella、劳氏黏虫 Mythimna loreyi等昆虫已经做了大量的耐寒性研究[17-21]。
关于草地贪夜蛾耐寒能力的研究国内已经有少量报道,如张智等测定了草地贪夜蛾不同龄期的抗寒能力[22];张悦等测定了取食不同寄主植物的草地贪夜蛾的过冷却点和体液冰点[23]。但是,温度作为草地贪夜蛾耐寒性的重要影响因素,相关研究还未见报道。
草地贪夜蛾在我国蔓延迅速,已经对我国的粮食安全产生了巨大威胁[24],亟待建立完善的预测预报体系对其进行防控。温度作为其种群分布区域的决定因素之一,有必要进行深入探究。本试验通过设定不同的饲养温度,测定了在不同温度条件下饲养的草地贪夜蛾各虫态的过冷却点和体液冰点,明确其抗寒能力,为预测其可能的分布范围提供科学依据。
1 材料与方法
1.1 供试虫源
试验用草地贪夜蛾幼虫于2019年4月采自广西壮族自治区南宁市(108.37°E,22.82°N),在室内人工饲养繁殖,建立种群。饲养条件为:温度(28±1)℃、相对湿度60%±5%、光周期L∥D=16 h∥8 h、光照强度18 000 lx。幼虫使用新鲜幼嫩玉米苗饲养,饲养密度为10头/瓶(750 mL,高14 cm,直径8 cm),在含水量为15%的灭菌土中化蛹,成虫用10%蜂蜜水补充营养。
1.2 仪器设备
仪器:RXM-508C 恒温培养箱,宁波江南仪器厂;亚都超声波加湿器,北京亚都环保科技有限公司;智能昆虫过冷却点测定仪(SUN-V 型),北京鹏程电子科技中心;高低温试验箱(BPH-060C 型),上海一恒科技有限公司。
1.3 草地贪夜蛾过冷却点和体液冰点测定
选取正常温度(28℃)条件下饲养的草地贪夜蛾同一天产的卵,分别放入不同的恒温培养箱,设定16、20、24、28、32、36℃共6个温度梯度,温差范围为±1℃,试虫整个世代发育均在恒温培养箱中完成。相对湿度为60%±5%,光周期为L∥D=16 h∥8 h、光照强度18 000 lx。从不同温度梯度的恒温培养箱中随机选取发育一致的草地贪夜蛾3龄、6龄幼虫,2日龄、5日龄蛹,3日龄成虫,测定过冷却点和体液冰点,每个处理测定30头以上。其中,3龄幼虫代表低龄幼虫;6龄幼虫代表高龄幼虫;2日龄蛹代表化蛹初期;5日龄蛹代表成熟的蛹;3日龄成虫为羽化后完成成虫期发育,即将进行生殖或迁飞的时期。测定时,把虫体置于1.5 mL离心管中,将过冷却点测定仪的热敏电阻感温探头与虫体充分接触,然后塞入棉花固定,注意不要伤害虫体。最后将离心管放入变温冰箱中,以每分钟1℃的速率从室温开始降温,最低温设置为-40℃。每秒记录1次虫体温度,通过配套软件记录数据,并绘制虫体温度变化曲线,确定其过冷却点和体液冰点[25]。
1.4 数据分析
试验中所有数据的平均值和标准误用SPSS 20软件计算。不同虫态的过冷却点和体液冰点首先进行正态性和方差齐性检验,经方差分析确定差异显著性后再用Tukeys HSD法进行差异显著性检验。利用单样本Kolmogorov-Smirnov 法检验不同虫态过冷却点和体液冰点的频次分布特征,并利用Origin 9.1绘图。
2 结果与分析
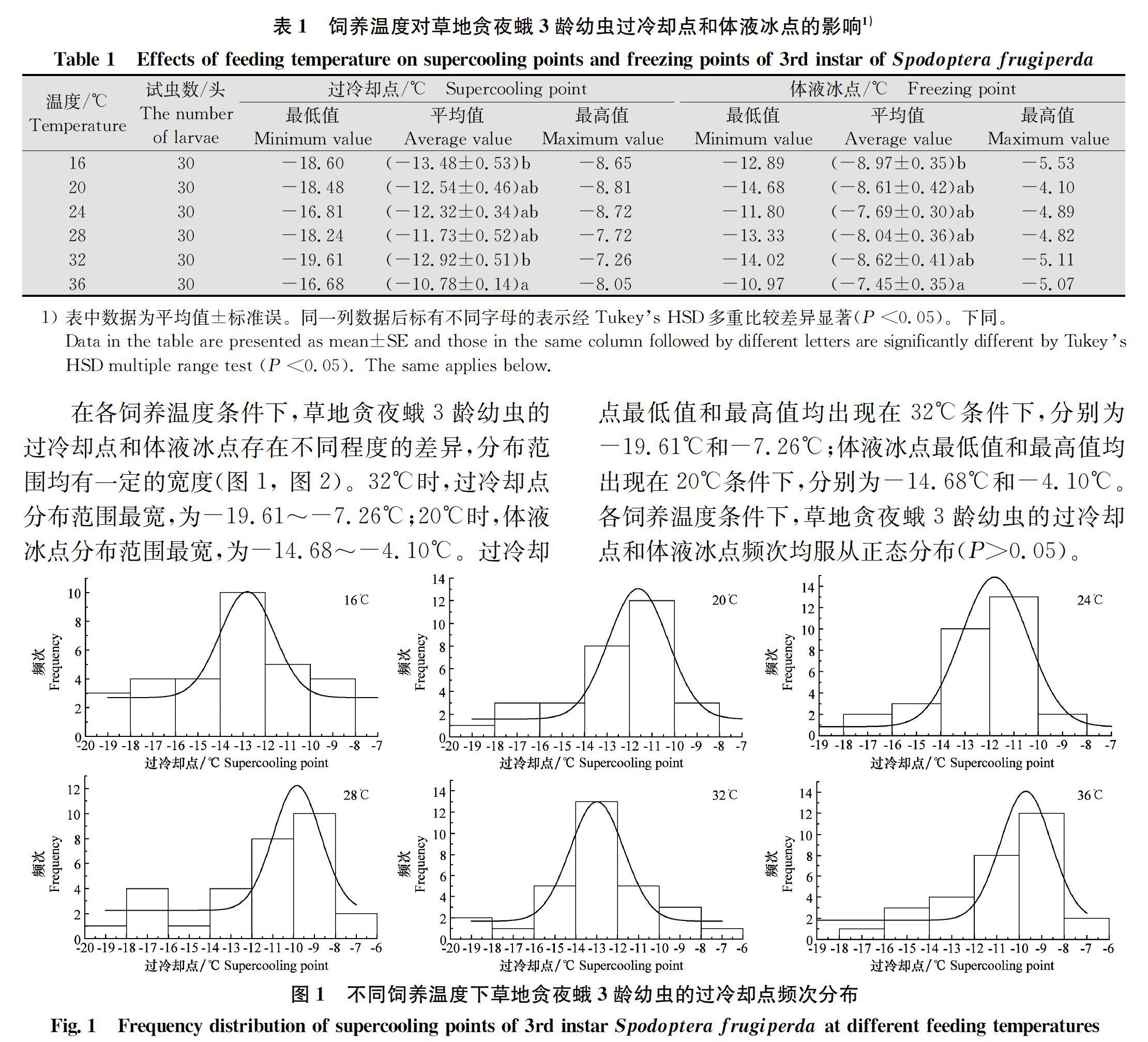
2.1 不同饲养温度下草地贪夜蛾3龄幼虫的过冷却点和体液冰点 不同饲养温度下草地贪夜蛾3龄幼虫的过冷却点和体液冰点存在显著差异(表1)。各饲养温度条件下草地贪夜蛾3龄幼虫的过冷却点由低到高的饲养温度依次为:16℃<32℃<20℃<24℃<28℃<36℃,其中,16℃条件下过冷却点最低,为(-13.48±0.53)℃,其次为32℃。体液冰点由低到高的饲养温度依次为:16℃<32℃<20℃<28℃<24℃<36℃,其中,16℃条件下体液冰点最低,为(-8.97±0.35)℃,其次为32℃。结果表明,低温(16℃)和适当高温(32℃)可以提高草地贪夜蛾3龄幼虫的抗寒能力,温度进一步升高,其抗寒能力下降。
在各饲养温度条件下,草地贪夜蛾3龄幼虫的过冷却点和体液冰点存在不同程度的差异,分布范围均有一定的宽度(图1, 图2)。32℃时,过冷却点分布范围最宽,为-19.61~-7.26℃;20℃时,体液冰点分布范围最宽,为-14.68~-4.10℃。过冷却点最低值和最高值均出现在32℃条件下,分别为-19.61℃和-7.26℃;体液冰点最低值和最高值均出现在20℃条件下,分别为-14.68℃和-4.10℃。各飼养温度条件下,草地贪夜蛾3龄幼虫的过冷却点和体液冰点频次均服从正态分布(P>0.05)。
2.2 不同饲养温度下草地贪夜蛾6龄幼虫的过冷却点和体液冰点 不同饲养温度下草地贪夜蛾6龄幼虫的过冷却点和体液冰点存在显著差异(表2)。其过冷却点由低到高的饲养温度依次为:16℃<36℃<32℃<24℃<28℃<20℃,其中,16℃条件下过冷却点最低,为(-8.94±0.28)℃,其次为36℃。体液冰点由低到高的饲养温度依次为:16℃<36℃<32℃<24℃<28℃<20℃,其中,16℃条件下体液冰点最低,为(-4.99±0.28)℃,其次为36℃。6龄幼虫的过冷却点和体液冰点均高于3龄幼虫,说明低龄幼虫具有更强的耐寒性,高龄幼虫对高温的耐受力更强。
在各饲养温度条件下,草地贪夜蛾6龄幼虫个体的过冷却点和体液冰点存在不同程度的差异,分布范围均有一定的宽度(图3和图4)。24℃饲养时,过冷却点和体液冰点分布范围均最宽,分别为-13.23~-5.11℃和-8.33~-1.24℃。过冷却点最低值出现在24℃条件下,最高值出现在20℃条件下,分别为-13.23℃和-4.16℃;体液冰点最低值出现在16℃饲养条件下,最高值出现在24℃条件下,分别为-8.57℃和-1.24℃。各饲养温度条件下,草地贪夜蛾6龄幼虫的过冷却点和体液冰点频次均服从正态分布(P>0.05)。
2.3 不同饲养温度下草地贪夜蛾2日龄蛹的过冷却点和体液冰点 不同饲养温度下草地贪夜蛾2日龄蛹的过冷却点和体液冰点存在显著差异(表3)。其过冷却点由低到高的饲养温度依次为:16℃<28℃<36℃<24℃<32℃<20℃,其中,16℃条件下过冷却点最低,为(-18.09±0.50)℃。体液冰点由低到高的饲养温度依次为:16℃<28℃<36℃<24℃<32℃<20℃,其中,16℃条件下体液冰点最低,为(-12.17±0.37)℃。2日龄蛹的过冷却点和体液冰点均低于幼虫期,具有更强的耐寒性。
在各饲养温度条件下,草地贪夜蛾2日龄蛹的过冷却点和体液冰点存在不同程度的差异,分布范围均有一定的宽度(图5和图6)。36℃时,过冷却点分布范围最宽,为-21.63~-6.39℃;36℃时,体液冰点分布范围最宽,为-16.39~-2.15℃。过冷却点最低值出现在28℃条件下,最高值出现在36℃条件下,分别为-21.99℃和-6.39℃;体液冰点最低值出现在28℃条件下,最高值出现在36℃条件下,分别为-18.38℃和-2.15℃。各饲养温度条件下,草地贪夜蛾2日龄蛹的过冷却点和体液冰点频次均服从正态分布(P>0.05)。
2.4 不同饲养温度下草地贪夜蛾5日龄蛹的过冷却点和体液冰点 不同饲养温度下草地贪夜蛾5日龄蛹的过冷却点和体液冰点存在显著差异(表4)。其过冷却点由低到高的饲养温度依次为:16℃<32℃<28℃<36℃<24℃<20℃,其中,16℃条件下过冷却点最低,为(-17.76±0.49)℃。体液冰点由低到高依次为:32℃<16℃<28℃<36℃<24℃<20℃,其中,32℃饲养条件下体液冰点最低,为(-11.72±0.56)℃。5日龄蛹与2日龄蛹相似,具有较强的耐寒性。
草地贪夜蛾5日龄蛹在各温度条件下个体的过冷却点和体液冰点存在不同程度的差异,分布范围均有一定的宽度(图7和图8)。36℃饲养时,过冷却点分布范围最宽,为-20.80~-6.33℃;体液冰点分布范围最宽,为-16.43~-3.16℃。過冷却点最低值出现在32℃条件下,最高值出现在36℃条件下,分别为-23.07℃和-6.33℃;体液冰点最低值出现在32℃条件下,最高值出现在36℃条件下,分别为-19.52℃和-3.16℃。各饲养温度条件下,草地贪夜蛾5日龄蛹的过冷却点和体液冰点频次均服从正态分布(P>0.05)。
2.5 不同饲养温度下草地贪夜蛾成虫的过冷却点和体液冰点 不同饲养温度下草地贪夜蛾成虫的过冷却点和体液冰点存在显著差异(表5)。其过冷却点由低到高的饲养温度依次为:16℃<28℃<24℃<20℃<36℃<32℃,其中,16℃条件下过冷却点最低,为(-17.87±0.55)℃。体液冰点由低到高的饲养温度依次为:28℃<16℃<24℃<32℃<20℃<36℃,其中,28℃条件下体液冰点最低,为(-11.23±0.61)℃。
在各饲养温度条件下,草地贪夜蛾成虫个体的过冷却点和体液冰点存在不同程度的差异,分布范围均有一定的宽度(图9和图10)。32℃时,过冷却点分布范围最宽,为-19.78~-5.63℃;28℃时,体液冰点分布范围最宽,为-18.32~-5.17℃。过冷却点最低值出现在16℃条件下,最高值出现在36℃条件下,分别为-22.06℃和-3.9℃;体液冰点最低值出现在28℃条件下,最高值出现在36℃条件下,分别为-18.32℃和-2.28℃。各饲养温度条件下,草地贪夜蛾成虫的过冷却点和体液冰点均服从正态分布(P>0.05)。
3 讨论
抗寒能力一定程度上决定了昆虫的分布范围,过冷却点作为昆虫能够存活的极限温度,是评价昆虫抗寒性的重要指标[26]。过冷却点越低,昆虫的抗寒性越强[27]。本文测定了不同温度条件下饲养的草地贪夜蛾不同虫态的过冷却点和体液冰点,结果显示:16℃条件下各龄期草地贪夜蛾的过冷却点显著低于其他温度;随着温度的升高,各龄期的过冷却点先升高后降低,温度继续升高,过冷却点再次降低。结果表明,低温对草地贪夜蛾的耐寒性具有明显的驯化作用,较低的温度或适当的高温可以提高其耐寒能力。
低温驯化可以启动昆虫应对低温的生理对策,包括过冷却点、小分子抗冻保护剂、抗冻蛋白和热休克蛋白等[28]。温度低于昆虫体液冰点而体液不凝固的现象称作过冷却现象。进入过冷却状态是北半球温带和寒温带地区昆虫采取的主要越冬策略[29]。本研究显示,16℃条件下饲养的草地贪夜蛾的过冷却点显著降低,2日龄蛹过冷却点-18.09℃低于黏虫的-14.05℃[20],甜菜夜蛾的-12.47℃[30],劳氏黏虫的-5.79℃[19]。低温驯化对草地贪夜蛾的耐寒性具有更明显的提升,这也预示草地贪夜蛾可能具有更广阔的越冬适宜区域。比较不同虫态的过冷却点发现,草地贪夜蛾蛹和成虫具有较低的过冷却点,因此推测蛹和成虫可能是其越冬虫态。但是,该推测还应结合潜在越冬区的实际虫情普查和寄主调查,最终确定其越冬虫态。
热休克蛋白(heat shock proteins, HSPs)是一种抗逆蛋白,通常被认为具有保护细胞不受伤害的作用。在大部分昆虫中,冷休克可以诱导热休克蛋白家族基因的表达[31]。本研究的结果显示,随着温度的升高,草地贪夜蛾的过冷却点和体液冰点在32℃时显著降低,这可能与热休克蛋白的功能有关。后续应对该现象进行深入研究。
已有研究表明,较低温度(15℃)下草地贪夜蛾成虫不能正常发育[32],因此本试验选择16℃作为最低处理温度。在储粮害虫[33]、甜菜夜蛾[30]耐低温能力的研究中研究者设置更低的温度(5、0、-10℃和-5℃)对昆虫进行驯化后,结果表明其耐低温能力表现出显著差异。苹浅褐卷蛾Epiphyas postvittana幼虫经冷驯化处理后,过冷却点和死亡率均明显降低[34]。美洲芹凤蝶Papilio zelicaon越冬蛹过冷却点为-20.5℃,当环境温度为-30℃时蛹依然能够存活,但经春季暖温暴露后,过冷却点升为-17℃,蛹在-20℃仅能存活1 h[35]。由此说明低温驯化对降低昆虫过冷却点至关重要。影响昆虫耐寒的因素主要包括生物因素和非生物因素。生物因素主要包括寄主营养、发育历期及雌雄差异等;非生物因素主要有季节性气温变化、快速冷驯化、环境湿度、光周期及不同地理位置等。昆虫的耐寒能力是各种因素相互作用的结果[36],是由多种抗寒物质组成的复杂系统[37-38]。因此,后续对于草地贪夜蛾耐寒性的研究也应综合考虑各种生物及非生物因素的影响,以期更全面地阐明其耐寒性,为其越冬区域的预测提供依据。
我们前期已经就温度对草地贪夜蛾的影响做了很多研究,包括不同温度下的飞行能力、不同温度下的两性生命表构建等[39-40],该试验是对草地贪夜蛾温度试验的补充,进一步阐明温度对草地贪夜蛾的影响。但是,在室内稳定条件下得出的温度对草地贪夜蛾的影响与自然环境中的可能会存在一定差异,需要进一步进行田间试验,从而更深入地了解野外环境下草地贪夜蛾的生物学习性。草地贪夜蛾从2019年1月入侵我国以来已经扩散到除新疆、西藏、内蒙古、东北以外的大部分地区,预计2020年其为害范围还将进一步扩大[41]。因此,急需对其生物学习性进行深入的研究,摸清其生长发育、繁殖、分布及为害规律,为虫情预测预报和虫害控制提供理论依据。
参考文献
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738.
[2] MUR U 'A G, MOLINA-OCHOA J, COVIELLA C. Population dynamics of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and its parasitoids in northwestern Argentina [J]. Florida Entomologist, 2006, 89(2): 175-183.
[3] LUGINBILL P. The fall army worm [R]. USDA Technology Bulletin, 1928, 34: 91.
[4] SPARKS A N. A review of the biology of the fall armyworm[J]. Florida Entomologist, 1979, 62(2): 82-87.
[5] JOHNSON C G. Migration and dispersal of insects by flight[M]. London: Methuen, 1969: 745-763.
[6] JOHNSON S J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere [J]. International Journal of Tropical Insect Science, 1987, 8(4/5/6): 543-549.
[7] DAY R, ABRAHAMS P, BATEMAN M, et al. Fall armyworm: impacts and implications for Africa[J]. Outlooks on Pest Management, 2017, 28(5): 196-201.
[8] 劉刚. 全国农技中心通报: 我国及周边国家草地贪夜蛾发生为害情况[J]. 农药市场信息, 2019(9): 59.
[9] 杨学礼, 刘永昌, 罗茗钟, 等. 云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J]. 云南农业, 2019(1): 72.
[10] 吴秋琳, 姜玉英, 吴孔明. 草地贪夜蛾缅甸虫源迁入中国的路径分析[J]. 植物保护, 2019, 45(2): 1-6.
[11] 郭井菲, 赵建周, 何康来, 等. 警惕危险性害虫草地贪夜蛾入侵中国[J]. 植物保护, 2018, 44(6): 1-10.
[12] 农业农村部就草地贪夜蛾防控工作举行新闻发布会[J]. 中国农技推广, 2019, 35(9): 101-103.
[13] BROUFAS G D, PAPPAS M L, KOVEOS D S. Effect of relative humidity on longevity, ovarian maturation, and egg production in the olive fruit fly (Diptera: Tephritidae) [J]. Annals of the Entomological Society of America,2009,102(1):70-75.
[14] FORSTER J, HIRST A G, WOODWARD G. Growth and development rates have different thermal responses [J]. The American Naturalist, 2011, 178(5): 668-678.
[15] 欧阳芳, 戈峰. 昆虫抗冻耐寒能力的测定与分析方法[J]. 应用昆虫学报, 2014, 51(6): 1646-1652.
[16] 景晓红, 康乐. 昆虫耐寒性的测定与评价方法[J]. 昆虫知识, 2004(1): 7-10.
[17] 罗立平,党英侨,张彦龙,等.不同寄主对花绒寄甲生长发育和耐寒性的影响[J].中国生物防治学报,2019,35(6):835-840.
[18] 孔维娜,王怡,郭永福,等. 梨小食心虫过冷却点及体液冰点测定[J]. 植物保护, 2019, 45(3): 102-105.
[19] 秦建洋, 兰建军, 张蕾, 等. 劳氏黏虫幼虫和蛹过冷却点及体液冰点的测定[J]. 中国植保导刊, 2018, 38(8): 33-38.
[20] 段云, 巩中军, 李慧玲, 等. 黏虫幼虫和蛹过冷却点及体液冰点的测定[J]. 植物保护, 2016, 42(4): 147-150.
[21] 薛冬, 陈丹, 范秀娟, 等. 烟草潜叶蛾的过冷却点测定[J]. 环境昆虫学报, 2014, 36(5): 860-864.
[22] 张智, 郑乔, 张云慧, 等. 草地贪夜蛾室内种群抗寒能力测定[J]. 植物保护,2019,45(6):43-49.
[23] 张悦, 邓晓悦, 张雪艳, 等. 取食不同食物的草地贪夜蛾的过冷却点和体液冰点[J/OL]. 植物保护: 1-6[2019-11-26]. https:∥doi.org/10.16688/j.zwbh.2019608.
[24] 郭井菲, 何康来, 王振营. 草地贪夜蛾的生物学特性、发展趋势及防控对策[J]. 应用昆虫学报, 2019, 56(3): 361-369.
[25] 秦玉川, 杨建才. 一种便携式测定昆虫过冷却点的方法[J]. 昆虫知识, 2000(4): 236-238.
[26] ASAHINA E. Frost resistance in insects [J].Advances in Insect Physiology, 1970, 6: 1-49.
[27] 鹿金秋, 王振营, 何康来, 等. 桃蛀螟越冬老熟幼虫过冷却点测定[J]. 植物保护, 2009, 35(2): 44-47.
[28] FULLER B J. Cryoprotectants: the essential antifreezes to protect life in the frozen state [J]. CryoLetters, 2004, 25(6): 375-388.
[29] 史彩華, 胡静荣, 李传仁, 等. 环境胁迫下昆虫的耐寒适应机制研究进展[J]. 植物保护, 2016, 42(6): 21-28.
[30] 江幸福, 罗礼智, 李克斌, 等. 甜菜夜蛾抗寒与越冬能力研究[J]. 生态学报, 2001(10): 1575-1582.
[31] JENSEN P, OVERGAARD J, LOESCHCKE V, et al. Inbreeding effects on standard metabolic rate investigated at cold, benign and hot temperatures in Drosophila melanogaster [J]. Journal of Insect Physiology, 2014, 62: 11-20.
[32] 何莉梅,葛世帅,陈玉超,等.草地贪夜蛾的发育起点温度、有效积温和发育历期预测模型[J].植物保护,2019,45(5):18-26.
[33] 霍鸣飞,吕建华,刘淑丽.不同低温驯化处理对重要储粮害虫过冷却点和冰点的影响[J].河南大学学报(自然科学版),2016,46(6):665-669.
[34] BRGI L P, MILLS N J. Cold tolerance of the overwintering larval instars of light brown apple moth Epiphyas postvittana [J]. Journal of Insect Physiology, 2010, 56(11): 1645-1650.
[35] WILLIAMS C M, NICOLAI A, FERGUSON L V, et al. Cold hardiness and deacclimation of overwintering Papilio zelicaon pupae [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2014, 178: 51-58.
[36] WILLIAMS C M, CHICK W D, SINCLAIR B J. A cross-seasonal perspective on local adaptation: metabolic plasticity mediates responses to winter in a thermal-generalist moth [J]. Functional Ecology, 2015, 29(4): 549-561.
[37] 张徐,吕宝乾,金启安,等.低温对椰心叶甲成虫体内几种抗寒物质含量的影响[J].热带作物学报,2013,34(5):942-946.
[38] 岳雷,周忠实,刘志邦,等.不同强度快速冷驯化对广聚萤叶甲成虫耐寒性生理指标的影响[J].昆虫学报,2014,57(6):631-638.
[39] 谢殿杰,张蕾,程云霞,等.不同温度下草地贪夜蛾年龄-阶段实验种群两性生命表的构建[J].植物保护,2019,45(6):20-27.
[40] 谢殿杰,张蕾,程云霞,等.温度对草地贪夜蛾飞行能力的影响[J].植物保护,2019,45(5):13-17.
[41] 吴秋琳,姜玉英,吴孔明.草地贪夜蛾缅甸虫源迁入中国的路径分析[J].植物保护,2019,45(2):1-6.
(责任编辑: 田 喆)
猜你喜欢
农家科技下旬刊(2019年10期)2019-11-29
植物保护(2019年5期)2019-11-27
植物保护(2019年5期)2019-11-27
现代农业科技(2019年19期)2019-11-21
现代农业科技(2019年19期)2019-11-21
农家科技(2019年10期)2019-11-13
农家科技(2019年7期)2019-10-08
农家科技(2019年7期)2019-10-08
农家科技(2019年8期)2019-09-19
南方农业·下旬(2019年4期)2019-08-26
