
牛磺酸对HepG2细胞甘油三酯合成的影响研究
2020-06-17 08:29马于巽郭俊霞夏天则
天然产物研究与开发 2020年5期
马于巽,宋 琪,张 静,,郭俊霞,,夏天则,陈 文,*
1北京联合大学 生物活性物质与功能食品北京市重点实验室,北京,100191;2北京联合大学生物化学工程学院食品科学系,北京,100023
牛磺酸(2-氨基乙磺酸,taurine)是一种内源性含硫氨基酸,可由甲硫氨酸和半胱氨酸合成,也可由饮食提供,在鱼贝类等海产品中含量丰富[1]。牛磺酸结构简单且不参与蛋白质合成,主要以游离形式存在于哺乳动物的肝脏、肾脏、脂肪组织、视网膜等组织器官,发挥维持视觉、调节渗透压、抗氧化、调节脂类代谢等作用[1,2]。动物实验表明牛磺酸可减少实验动物体脂,降低血液中甘油三酯水平,改善脂肪肝[3,4]。体外实验显示,在大鼠H4IIE肝细胞和原代肝细胞中,牛磺酸减少了棕榈酸和油酸导致的甘油三酯过度积累[5],而Yanagita等[6]通过在HepG2细胞培养液中添加14C标记的油酸,发现牛磺酸抑制了50%14C标记的油酸合成甘油三酯,提示牛磺酸可抑制HepG2细胞甘油三酯合成。牛磺酸抑制甘油三酯合成相关酶的报道较少,并且关于牛磺酸降低细胞脂质的有效浓度报道不一,需要进一步深入的研究[5-8]。
固醇调节元件结合蛋白(sterol-regulatory element binding proteins,SREBPs)是一种调节脂肪酸、甘油三酯和胆固醇合成代谢的关键核转录因子,有三种亚型:SREBP-1a、SREBP-1c和SREBP-2。SREBP-2存在于大多数组织细胞中,主要参与调节胆固醇的合成;SREBP-1a 在肝脏及肾上腺较丰富,可调节脂肪酸、甘油三酯、胆固醇、磷脂等多种脂质的生物合成,高浓度时促进胆固醇的合成,低浓度时促进脂肪酸的合成[9]。SREBP-1c主要表达在肝脏和脂肪细胞中,主要参与肝脏脂肪酸合成和脂肪前体细胞的分化,是脂肪酸和甘油三酯合成中关键调控因子,可调控乙酰辅酶 A合成酶(acetyl-CoA synthetase,AceCS)、乙酰辅酶 A 羧化酶(acetyl-CoA carboxylase,ACC)、脂肪酸合成酶(fatty acid synthetase,FAS)、硬脂酰 CoA 去饱和酶1(stearyl CoA desaturase 1,SCD-1)等脂肪酸合成相关的酶[10,11]。
本实验以油酸(oleic acid,OA)建立HepG2细胞脂肪变性模型,探讨牛磺酸对正常与高脂条件下HepG2细胞的甘油三酯水平、SREBP-1c及FAS等脂肪合成相关酶蛋白表达的影响,以期为牛磺酸预防/改善机体高脂状态的深入研究提供参考。
1 材料与方法
1.1 材料与试剂
HepG2细胞购自协和医科大学基础医学研究所细胞中心; DMEM细胞基础培养基、ECL显影试剂盒、CCK-8试剂盒购自美国GenView公司;胎牛血清购自美国Gibco公司;双抗(青霉素、链霉素)、RIPA裂解液、PMSF蛋白酶抑制剂、二喹啉甲酸(bicinchoninic acid,BCA)试剂盒购自北京鼎国昌盛生物技术有限公司;牛磺酸(纯度>99%)、油酸(纯度>99%)购自美国Sigma公司;甘油三酯(TG)测定试剂盒购自南京建成生物工程研究所;SREBP-1c一抗(AF4782,兔抗)购自美国Affinity公司;FAS(3180)、AceCS1(3658)、ACSL1(9189)、ACC(3676)、p-ACC(11810)、GAPDH(2118)等一抗(兔抗)、二抗(7074S,抗兔IgG-HRP)购自美国CST(Cell Signaling Technology)公司。
1.2 仪器与设备
酶标仪购自美国Thermo Fisher公司;电泳仪、转膜仪购自美国Bio-Rad公司;凝胶成像系统购自北京Thmorgan生物科技有限公司。
1.3 方法
1.3.1 细胞培养及处理
细胞培养:用含10%胎牛血清的DMEM培养基,37℃、5% CO2恒温培养HepG2细胞,以1×105~2×105个/孔接种于6孔板中。
牛磺酸对HepG2细胞甘油三酯的影响:在培养液中加入以PBS溶解的终浓度分别为1、5、10、20 mmol/L的牛磺酸,培养24、48、72 h。
高脂模型的建立:以无水乙醇为溶剂,在培养液中加入终浓度分别为0.05、0.1、0.2 mmol/L的油酸,培养6、12、24 h后测定细胞内甘油三酯水平。
牛磺酸对高脂HepG2细胞甘油三酯的影响:设立对照组(N),高脂模型组(OA),在模型基础上加入1、5、10、20 mmol/L牛磺酸即牛磺酸组(O+T),共同培养24、48、72 h。
1.3.2 细胞内甘油三酯测定[12]
细胞培养一定时间后弃上清,用PBS洗2次,每孔加入1mL正己烷-异丙醇(2∶1,V/V)置于摇床上慢摇1 h,以提取细胞内脂类物质;4 ℃、10 000 rpm离心10 min,上清通氮气除去有机溶剂;以80 μL含10% TritonX-100的异丙醇涡旋震荡溶解脂提取物后即得到待测样品。以甘油三酯测定试剂盒检测样品甘油三酯浓度。
1.3.3 细胞内蛋白含量的测定
细胞完成脂类物质提取后加入裂解液冰上提取细胞内总蛋白,4 ℃、12 000 rpm离心10 min,上清液即为待测样品。以BCA蛋白测定试剂盒测蛋白样品的浓度。
细胞内甘油三酯含量以甘油三酯与蛋白质比值(μmol/g·pro)表示。
1.3.4 细胞存活率测定
取2×104~4×104个/孔细胞铺于96孔板中,设立:对照组(N),1、5、10、20 mmol/L牛磺酸组(T),无水乙醇对照组(E),油酸组(OA),在油酸基础上加入1、5、10、20 mmol/L牛磺酸(O+T)。培养24、48 h后用CCK-8试剂盒在450 nm测定吸光度。以正常对照为100%,计算细胞存活率。
1.3.5 Western blot
加入PMSF终浓度为1 mmol/L的裂解液提取细胞内总蛋白,以BCA蛋白测定试剂盒测定蛋白浓度,加入SDS蛋白电泳上样缓冲液后沸水浴5 min使蛋白充分变性。每孔上样30 μg样品蛋白进行聚丙烯酰胺凝胶电泳,转至PVDF膜后用5%脱脂奶粉封闭1 h,加入不同目的蛋白的一抗和内参GAPDH抗体4 ℃孵育过夜。TBST洗三次后加入二抗,常温孵育2 h,TBST洗三次后用ECL显影试剂盒显影。条带结果用凝胶图像分析系统进行目的蛋白及内参(GAPDH)灰度值分析,以目的蛋白灰度值与内参蛋白灰度值之比作为所测目的蛋白的相对含量。
1.4 数据处理
2 结果与分析
2.1 牛磺酸对HepG2细胞甘油三酯水平的影响
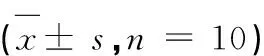
表1 牛磺酸对HepG2细胞甘油三酯水平的影响Table 1 Effect of taurine on triglyceride level in HepG2 ± s,n = 10)(μmol/g·pro)
由表1可知,在HepG2细胞中添加1、5、10、20 mmol/L牛磺酸分别作用24、48、72 h,甘油三酯水平有随牛磺酸浓度升高而降低的趋势,但是牛磺酸各剂量组与正常对照组相比无显著差异,提示牛磺酸对正常HepG2细胞甘油三酯水平无显著影响(P>0.05)。
2.2 油酸对HepG2细胞甘油三酯水平的影响
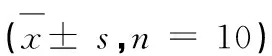
表2 油酸对HepG2细胞甘油三酯水平的影响Table 2 Effect of oleic acid on triglyceride level in HepG2 ± s,n = 10) (μmol/g·pro)
注:*与对照组相比,P<0.05。
Note:*Compared with control,P<0.05.
表2显示:0.025 mmol/L油酸作用24 h,细胞内甘油三酯水平升高22.77%(P<0.05);0.05 mmol/L油酸作用6 h,细胞内甘油三酯水平升高29.28%(P<0.05),作用12和24 h甘油三酯含量均升高50%以上(P<0.05);0.1、0.2 mmol/L油酸作用12和24 h甘油三酯含量均升高80%以上。故本实验选择0.05 mmol/L油酸来建立高脂模型。
2.3 牛磺酸对油酸处理的HepG2细胞甘油三酯水平的影响
表3 牛磺酸对油酸处理的HepG2细胞甘油三酯水平的影响Table 3 Effect of taurine on triglyceride level in HepG2 cells treated with oleic ± s,n = 10)(μmol/g·pro)
注:*与对照组相比P<0.05;#与模型组相比P<0.05。
Note:*Compared with control,P<0.05;#Compared with model,P<0.05.
由表3可知,与对照组相比,0.05 mmol/L油酸作用24和48 h细胞内甘油三酯水平升高约50%(P<0.05)。但培养72 h,细胞内甘油三酯水平仅升高35.25%(P<0.05),提示随培养时间延长,细胞内甘油三酯水平并不持续升高。与高脂模型组相比:1 mmol/L牛磺酸需作用72 h才使甘油三酯水平明显下降(P<0.05),5和10 mmol/L牛磺酸作用24、48、72 h以及20 mmol/L牛磺酸作用24和48 h细胞内甘油三酯水平均明显下降(P<0.05),20 mmol/L牛磺酸作用72 h对细胞内甘油三酯水平无明显影响(P>0.05)可能是由于随培养时间延长,细胞衰老,模型组甘油三酯水平下降。
2.4 牛磺酸和油酸对HepG2细胞存活率的影响
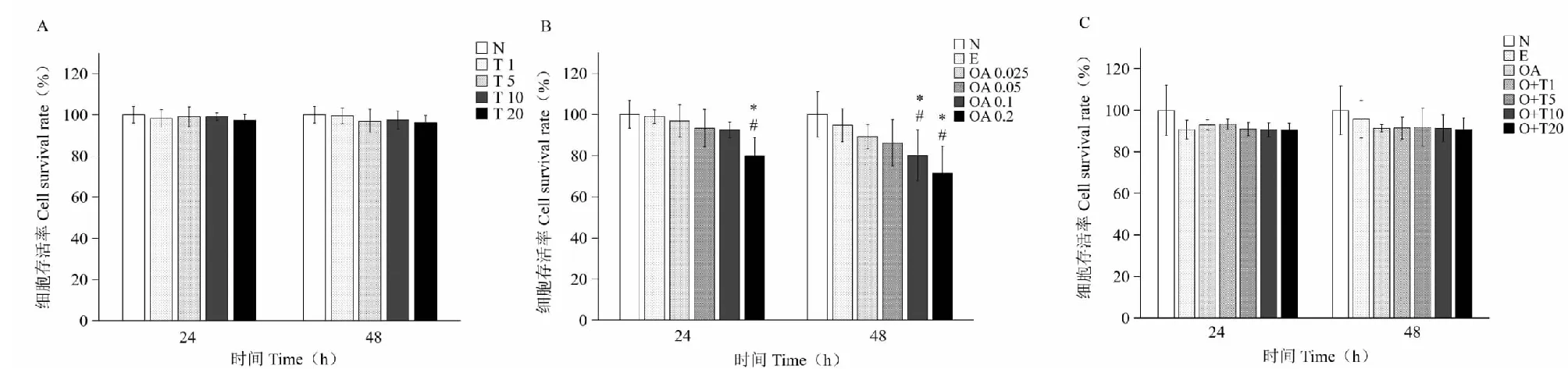
图1 牛磺酸和油酸对HepG2细胞存活率的影响Fig.1 The effect of taurine and oleic acid on the viability of HepG2 cells注:A:牛磺酸对细胞存活率的影响;B:油酸对细胞存活率的影响;C:油酸与牛磺酸共同作用对细胞存活率的影响;N:对照组;T1、T5、T10、T20表示牛磺酸终浓度为1、5、10、20 mmol/L;E:无水乙醇对照组;OA 0.05、0.1、0.2表示油酸终浓度为0.05、0.1、0.2 mmol/L;OA:油酸对照组;O+T1、O+T5、O+T10、O+T20表示在油酸基础上添加终浓度为1、5、10、20mmol/L的牛磺酸;*与对照组相比P<0.05;#与乙醇对照组相比P<0.05。Note:A,B,C represent the effect of taurine,oleic acid and co-action of oleic acid and taurine on cell survival,respectively.N:Control;T1,T5,T10,T20:The concentration of taurine is 1,5,10,20 mmol/L;E:Absolute ethyl alcohol control group;OA 0.05,0.1,0.2:The concentration of oleic acid is 0.05,0.1,0.2 mmol/L;OA:Oleic acid control group;O+T1,O+T5,O+T10,O+T20:0.05mmol/L oleic acid with 1,5,10,20 mmol/L taurine;*Compared with control,P<0.05;#Compared with absolute ethyl alcohol control,P<0.05.
由图1A可知1~20 mmol/L牛磺酸作用24和48 h,各组细胞存活率无明显差异。由图1B可知与正常对照和乙醇对照比0.1 mmol/L油酸作用48 h细胞存活率显著下降(P<0.05),0.2 mmol/L油酸作用24和48 h细胞存活率均显著下降(P<0.05)。由图1C可知在0.05 mmol/L油酸基础上添加1~20 mmol/L牛磺酸作用24和48 h,各组细胞存活率无明显差异(P>0.05)。
综合以上结果(表3、图1),0.05 mmol/L油酸作用24h,细胞甘油三酯水平升高约50%,表示模型建立成功。油酸是脂溶性物质,易进入细胞,并在较短时间内合成甘油三酯。随培养时间延长,细胞衰老,细胞内甘油三酯水平下降,所以细胞甘油三酯含量并不随着油酸作用时间的延长而继续升高,而是维持在一定水平并略呈下降趋势。5 mmol/L牛磺酸作用24 h细胞甘油三酯水平即显著下降(P<0.05),所以本实验以0.05 mmol/L油酸建立高脂模型,以5 mmol/L牛磺酸作用24 h进行后续SREBP-1c及脂肪合成相关酶蛋白表达的实验,此浓度和时间下牛磺酸和油酸对细胞存活率均无明显影响(P>0.05)。
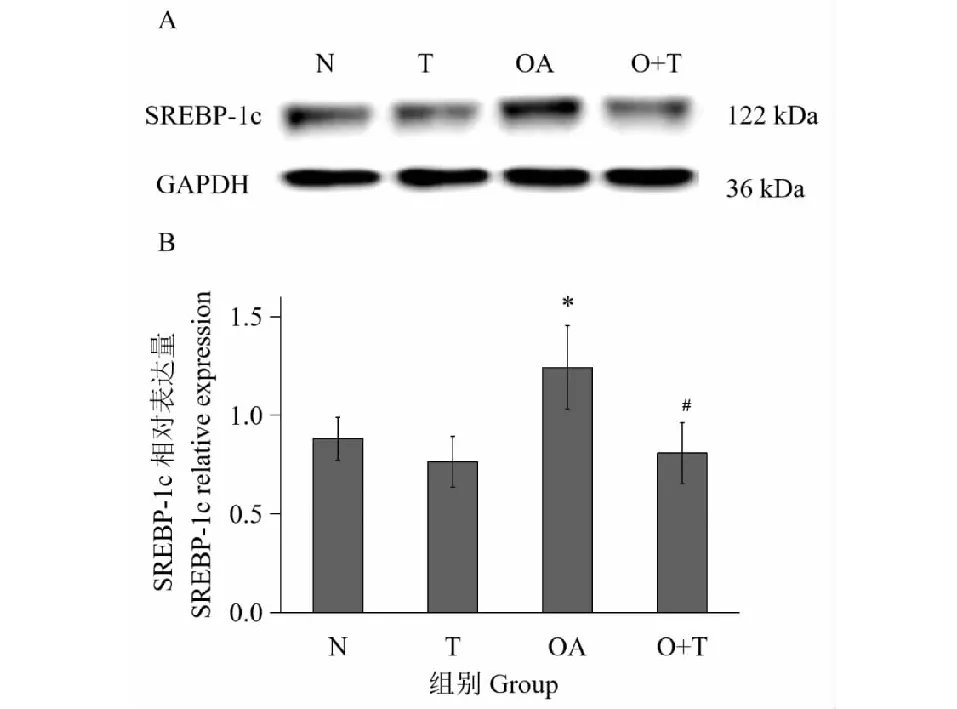
图2 牛磺酸对HepG2细胞SREBP-1c表达的影响Fig.2 The effect of taurine on the protein expression of SREBP-1c in HepG2 cells注:A:SREBP-1c和GAPDH蛋白条带图;B:SREBP-1c相对表达柱状图;N:对照组;T:牛磺酸组;OA:油酸组;O+T:油酸+牛磺酸组;*与正常对照组相比P<0.05;#与模型对照组相比P<0.05。Note:A:SREBP-1c and GAPDH protein bands picture;B:SREBP-1c relative expression histogram;N:Control;T:Taurine;OA:Oleic acid;O+T:Oleic acid+taurine;*Compared with control,P<0.05;#Compared with oleic acid group,P<0.05.
2.5 牛磺酸对HepG2细胞SREBP-1c表达的影响
由图2可知,与对照组相比,5 mmol/L牛磺酸对SREBP-1c表达无明显影响(P>0.05),高脂模型组SREBP-1c表达升高(P<0.05)提示油酸可增加HepG2细胞SREBP-1c的表达;与模型组相比,牛磺酸使SREBP-1c表达下降(P<0.05),提示牛磺酸可能通过降低SREBP-1c表达而调控甘油三酯合成。
2.6 牛磺酸对HepG2细胞中甘油三酯合成相关酶表达的影响
图3显示:与对照相比,牛磺酸组ACC表达下降(P<0.05),FAS、AceCS1、ACSL1表达无明显变化,而高脂模型组FAS、ACC、AceCS1、ACSL1表达上升(P<0.05);与模型组相比,油酸+牛磺酸组FAS、ACC、AceCS1、ACSL1表达下降(P<0.05)。以上结果表明油酸可增加细胞FAS、ACC、AceCS1、ACSL1的表达,增加甘油三酯的合成;而牛磺酸能降低细胞ACC的表达,降低高脂HepG2细胞FAS、ACC、AceCS1、ACSL1的表达,提示牛磺酸通过抑制脂肪酸及甘油三酯合成相关酶的表达而抑制甘油三酯合成。图3B显示:与对照相比牛磺酸使P-ACC表达上升(P<0.05),与模型组相比油酸+牛磺酸组P-ACC表达增加(P<0.05),提示牛磺酸除影响ACC的表达外,还通过增加ACC磷酸化抑制其活性,从而抑制脂肪酸合成。
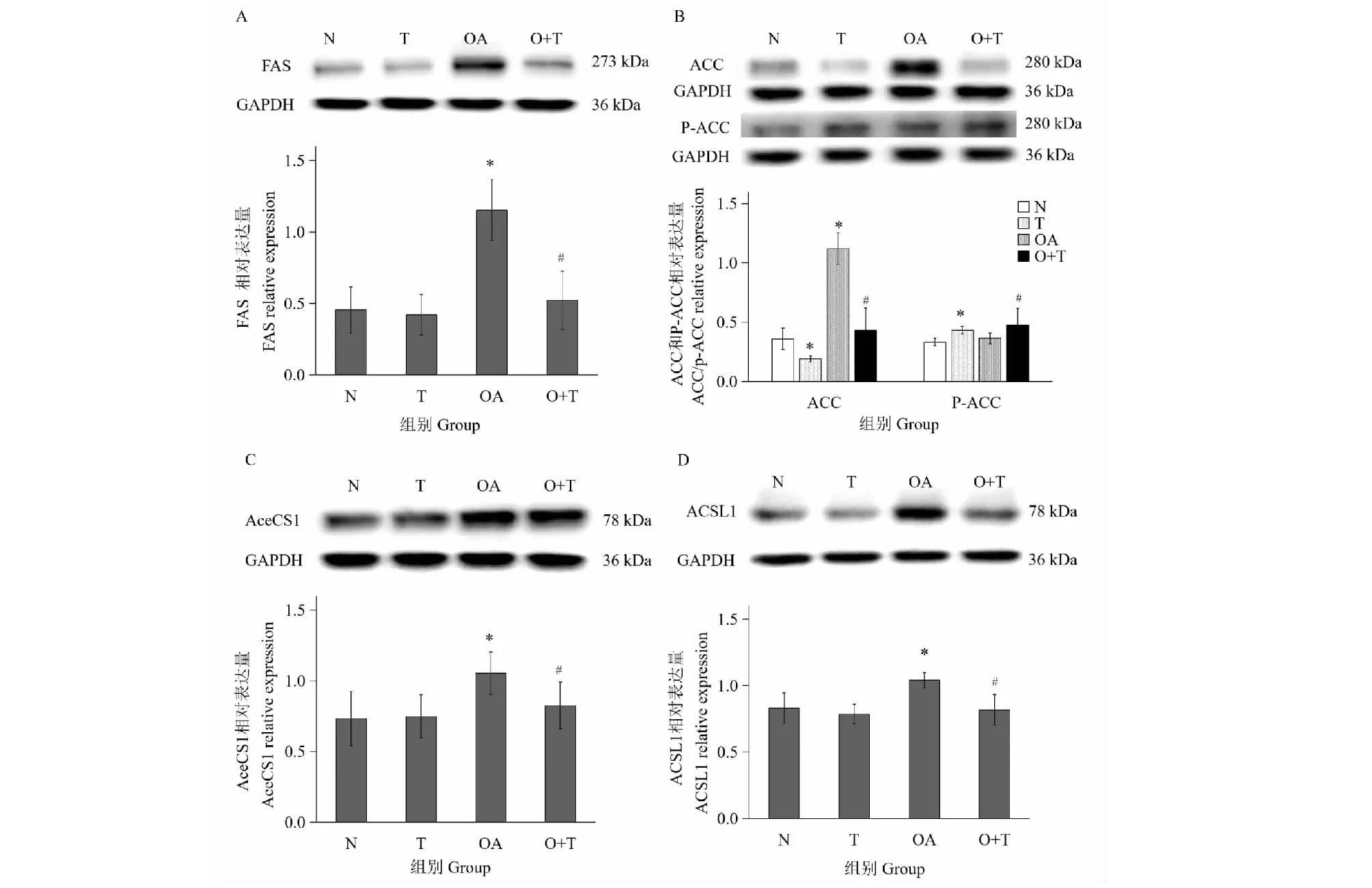
图3 牛磺酸对HepG2细胞中甘油三酯合成相关酶表达的影响Fig.3 Effect of taurine on the protein expression of TG synthesis related enzymes in HepG2 cells注:A、B、C、D分别为FAS、ACC/P-ACC、AceCS1、ACSL1蛋白表达结果及目的蛋白相对表达量柱状图;N:对照组;T:牛磺酸组;OA:油酸组;O+T:油酸+牛磺酸组;*与正常对照组相比P<0.05;#与模型对照组相比P<0.05。Note:A,B,C,D represent the protein expression of FAS,ACC /p-ACC,AceCS1,ACSL in Hepg2 cells,respectively.N:Control;T:Taurine;OA:Oleic acid;O+T;Oleic acid + taurine;*Compared with control,P<0.05;#Compared with oleic acid group,P<0.05.
3 讨论
研究显示,在大鼠[13]、小鼠[14]、豚鼠[4]、线虫[15]等模式生物中,牛磺酸均表现出降低甘油三酯、减少体脂肪/肝脏脂肪蓄积的作用;临床实验[16]和流行病学调查[17]也显示口服牛磺酸或饮食习惯中富含牛磺酸能在一定程度上改善高血脂、肥胖,降低BMI以及动脉粥样硬化指数, 降低高血脂症等代谢疾病和心血管疾病的发生率。
牛磺酸降甘油三酯作用的体外实验及机制研究较少。Gentile等[5]报道,牛磺酸能减少大鼠H4IIE肝细胞和原代肝细胞中甘油三酯过度积累,并降低棕榈酸介导的caspase-3活性、细胞凋亡、内质网应激和氧化应激,该研究提示在饮食中添加牛磺酸对预防非酒精性脂肪肝病(nonalcoholic fatty liver disease,NAFLD)的发生有良好作用。Yanagita等[6]的研究显示,在培养基中添加1 mmol/L牛磺酸作用24 h可以使HepG2细胞内甘油三酯水平下降近25%,而Hoang等[7]报道0.01 mmol/L牛磺酸使正常HepG2细胞甘油三酯水平减少近22%。两个研究团队的结果虽相似但在牛磺酸浓度上却相差了百倍之多。与他们的研究不同,本实验以1、5、10、20 mmol/L牛磺酸处理HepG2细胞24、48、72 h,细胞内甘油三酯水平虽有下降趋势,但并无显著降低(P>0.05);以0.05 mmol/L油酸建立高脂模型后,对于油酸处理的HepG2细胞,牛磺酸则显示出明显的降低甘油三酯水平的作用,其中1 mmol/L牛磺酸作用72 h、5/10 mmol/L牛磺酸作用24、48、72 h、20 mmol/L牛磺酸作用24/48 h都可使细胞内甘油三酯含量显著下降(P<0.05)。
机体甘油三酯代谢的主要途径有合成代谢和分解代谢,但是关于牛磺酸促进甘油三酯代谢的作用机制研究,主要集中在促进脂肪分解代谢和脂肪酸β氧化,此外还有抑制炎症反应和提高抗氧化能力等方面[5,13]。牛磺酸抑制甘油三酯合成的具体机制尚需深入研究。
SREBP-1c主要参与肝脏脂肪酸合成和脂肪前体细胞的分化,是脂肪酸和甘油三酯合成中关键调控因子。SREBP-1c的调控除了受细胞胆固醇水平影响外还受机体营养条件和激素水平的影响,它在营养过剩时表达升高,在饥饿禁食状态表达降低[10,11]。有研究表明,无论是ob/ob小鼠还是高脂饮食诱导的肥胖小鼠,均存在肝脏SREBP-1c 水平异常升高的现象,SREBP-1c 过度表达会引起糖脂代谢紊乱,导致非脂肪组织脂质积聚,引起组织病变,因此SREBP-1c 过度表达与肥胖相关疾病如糖尿病、高脂血症、代谢综合征的发生和发展有着密切关系[10,11]。Hoang等[7]曾报道,0.01 mmol/L牛磺酸可抑制HepG2细胞SREBP-1c表达但不影响其下游靶基因FAS表达。与Hoang的结果不同,本研究显示牛磺酸对正常HepG2细胞SREBP-1c表达无影响,但当以油酸建立高脂模型时,0.05 mmol/L油酸可增加细胞SREBP-1c的表达,5 mmol/L牛磺酸则抑制了油酸诱导的SREBP-1c表达增加,提示正常状态下牛磺酸不影响细胞SREBP-1c表达,仅在高脂状态下抑制SREBP-1c表达。
脂肪酸从头合成的二碳单位是乙酰辅酶A,合成起始于ACC催化乙酰辅酶A转化为丙二酸单酰辅酶A,乙酰辅酶A和丙二酸单酰辅酶A在FAS催化下经缩合、还原、脱水、再还原四步循环反应完成脂肪酸的合成[18]。ACC的活性受可逆磷酸化快速调控,磷酸化使ACC失活,去磷酸化使ACC被激活。FAS和ACC都是脂肪酸生物合成过程中的关键酶,AceCS可催化醋酸盐和辅酶A 转变为乙酰辅酶A,由 AceCS1 合成的乙酰CoA 用于脂肪酸和脂类生物合成[19]。FAS、ACC、AceCS1的表达均受到SREBP-1c的调控[10,11]。本研究显示:0.05mmol/L油酸诱导了HepG2细胞FAS、ACC、AceCS1的表达;5 mmol/L牛磺酸作用24 h可减少正常细胞/高脂细胞ACC的表达、增加p-ACC的表达,并抑制高脂细胞FAS、AceCS1的表达。提示牛磺酸通过抑制SREBP-1c的表达抑制脂肪酸合成相关酶的表达,并且通过增加ACC磷酸化水平抑制其活性从而减少脂肪酸的合成。
长链酰基辅酶A合成酶(long chain acyl-CoA synthetases,ACSL)催化脂肪酸与辅酶A 的连接,形成脂酰辅酶A。ACSL1主要表达在肝脏和脂肪细胞中,由ACSL1催化形成的脂酰辅酶A与L-α-磷酸甘油是甘油三酯合成的前体。ACSL1过度表达会导致脂代谢紊乱和甘油三酯沉积,Li[20]用腺病毒转染大鼠原代肝细胞过度表达ACSL1,检测到14C标记的油酸盐与甘油二酯的结合增加,细胞内甘油三酯水平增加。本研究显示,油酸增加了HepG2细胞ACSL1表达,牛磺酸不影响正常细胞ACSL1表达,但可抑制高脂HepG2细胞的ACSL1表达。ACSL1是PPARγ的靶基因,这一结果提示牛磺酸除通过SREBP-1c途径抑制甘油三酯合成外还可能通过PPARγ抑制甘油三酯合成。
4 结论
牛磺酸对HepG2细胞甘油三酯水平无影响,但能显著降低油酸诱导的高脂细胞的甘油三酯水平,明显减少高脂细胞SREBP-1c、FAS、ACC、AceCS1、ACSL1的表达,提高p-ACC的表达。这一结果提示牛磺酸通过调控SREBP-1c及其下游靶基因而抑制高脂细胞脂肪酸/甘油三酯的合成。深入的研究将进一步揭示在高脂膳食状态下牛磺酸营养的作用。
猜你喜欢
中国畜牧杂志(2022年5期)2022-11-06
生物技术进展(2022年5期)2022-10-11
中老年保健(2022年4期)2022-08-22
电脑报(2022年25期)2022-07-05
新农业(2020年18期)2021-01-07
食品与健康(2020年7期)2020-07-09
现代农村科技(2020年2期)2020-03-24
华声文萃(2019年4期)2019-09-10
文萃报·周二版(2019年10期)2019-09-10
阅读与作文(小学高年级版)(2019年2期)2019-03-27
