
墨兰‘吴字翠’× 墨兰‘红花’杂交后代根状茎增殖及芽分化研究
2020-06-19 08:53谢娟李龙邢玥翁青史樊晚林彭东辉
热带作物学报 2020年5期
谢娟 李龙 邢玥 翁青史 樊晚林 彭东辉

摘 要:墨蘭(Cymbiduim sinense)又称报岁兰,是兰科兰属植物,以其株型健壮、花期在春节深受国人喜爱。鉴于墨兰组培根状茎增殖率低、分化成芽困难,本研究以杂交墨兰根状茎为材料,通过不同浓度的生长素、分裂素以及有机添加物的组合,以期找到适宜墨兰根状茎增殖和分化的培养基配方。结果表明,最适宜杂交墨兰根状茎增殖的培养基配方为MS+ 2 mg/L 6-BA+0.1 mg/L NAA+0.2 mg/L TDZ+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag,增殖系数达5.35,生长速度为3.44。在添加不同激素的情况下,最适宜叶芽分化的培养基为MS+10.0 mg/L 6-BA+0.7 mg/L NAA+0.2 mg/L TDZ+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag,叶芽分化率为52.5%。在添加其他添加物的情况下,墨兰叶芽分化最适宜的培养基为MS+8.0 mg/L 6-BA+0.1 mg/L NAA+7.0 g/L蛋白胨+100.0 mg/L CW+1.0 g/L AC,叶芽分化率为173.30%。本研究探讨了激素和添加物对根状茎增殖分化的影响,可实现杂交品种组培苗大量繁殖。
关键词:墨兰;根状茎;增殖;分化;诱导
中图分类号:S682.31 文献标识码:A
Rhizome Proliferation and Bud differentiation of Cymbiduim sinense ‘Wuzicui × C. sinense ‘Honghua Hybrid
XIE Juan, LI Long, XING Yue, WENG Qingshi, FAN Wanlin, PENG Donghui*
College of Landscape Architecture, Fujian Agriculture and Forestry University / Key Laboratory of Orchidaceous Plant Conservation and Utilization, State Forestry and Grassland Administration, Fuzhou, Fujian 350002, China
Abstract: Cymbiduim sinense is also called Bao Sui Lan (Heralding New Year Orchid), which belongs to Cymbiduim (Orchidaceae). It is loved by Chinese people for its robust plant type and flowering period during the Spring Festival. In view of the low proliferation rate and the difficulty of bud differentiation of the rhizome of C. sinense. In this study, the hybrid rhizomes of C. sinense were used as materials, through the combination of auxin, mitogen and organic additives in different concentrations, in order to find a suitable medium formula for the proliferation and differentiation of the rhizomes of hybrid C. sinense. The results showed that the most suitable medium for rhizomes proliferation was MS+ 8 mg/L 6-BA+0.1 mg/L NAA+0.8 mg/L TDZ+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag, with a proliferation coefficient 5.35 and the growth rate 3.44. The most suitable medium for leaf bud differentiation was MS+10.0 mg/L 6-BA+ 0.7 mg/L NAA+0.2 mg/L TDZ+1.5 g/L AC+ 35.0 g/L Su+6.0 g/L Ag, with a leaf bud differentiation rate 52.5%. The most suitable medium for leaf bud differentiation was MS+8.0 mg/L 6-BA+0.1 mg/L NAA+7.0 g/L peptone+100.0 mg/L CW+1.0 g/L AC, with a leaf bud differentiation rate 173.30%. This study explored the effects of hormones and additives on the proliferation and differentiation of the rhizomes, which could achieve large-scale reproduction of the tissue culture seedlings.
Keywords: Cymbiduim sinense; rhizome; proliferation; differentiation; induction
DOI: 10.3969/j.issn.1000-2561.2020.05.019
墨兰(Cymbidium sinense)又称报岁兰,兰科(Orchidaceae)兰属(Cymbidium)植物,原产于我国、越南和缅甸,因其花期在春节,深受国人喜爱,具有较高的经济价值、观赏价值和药用价值。
墨兰的繁殖方式很多,常见的有分株繁殖和组织培养方式。传统的繁殖方式主要是分株繁殖,但面临繁殖率较低、周期长等问题,无法实现大量生产和满足市场的需求。随着组培技术的发展,国内外学者开展了墨兰组培繁殖研究,其中在墨兰根状茎的增殖研究中,较适合的基本培养基有1/4MS、1/2MS、MS、Hyponex1-2和KC,常用的激素有6-BA、NAA、KT、IBA、TDZ等,但其适宜的浓度并不一致[1]。施福军等[2]研究表明2.0 mg/L 6-BA和1.0 mg/L NAA有利于金嘴墨兰根状茎增殖,增殖系数达8.1。左利娟等[3]研究发现2.0 mg/L 6-BA+0.5 mg/L KT+0.05 mg/L NAA有利于墨兰杂交后代增殖分化,增殖系数达3.96;丁雪珍等[4]发现2.0 mg/L BA和0.1 mg/L NAA有利于金锦凤墨兰根状茎增殖,增殖系数达6.6。
在墨兰芽分化相关研究中发现添加6-BA和NAA、TDZ可促进根状茎的芽诱导[5-6],傅雪琳等[7]研究表明5.0 mg/L 6-BA+0.5 mg/L NAA能够使芽的分化率达180%;朱根发等[8]发现在MS培养基中加入10 mg/L 6-BA和适量无机盐有利于墨兰根状茎芽的分化。許申平等[9]提到5 mg/L 6-BA与0.5 mg/L NAA的组合,墨兰根状茎绿芽数达到6.43;Yan等[10]研究表明4.0 mg/L 6-BA与1.0 g/L NAA组合时,芽数达2.9;Chen等[11]发现大花惠兰(Cymbidium hybrida)×墨兰(C. sinense)和惠兰(C. faberi)根状茎在含有1.0 mg/L BA和0.5 mg/L NAA的培养基中,根状茎顶端80%以上发育成芽;Gao等[12]研究在培养基中添加4 mg/L 6-BA和0.2 mg/L NAA可促进根茎的芽诱导。以上研究结果中发现6-BA的浓度高低对芽分化促进的结果存在较大出入。除了NAA和6-BA对叶芽分化有促进作用,TDZ也有利于芽分化。Chang等[13]提到添加TDZ有利于芽分化。
综上所述,虽然墨兰根状茎的组培增殖及芽分化的相关研究已经取得了较大的进展,但研究结果存在差异,且并不统一。在墨兰组织培养研究中,仍存在繁殖系数偏低、芽分化系数低和繁殖周期长等问题,并且制约了种苗规模化生产。除此之外,不同品种之间存在较大差异,导致其研究结果存在较大差异。本试验以杂交墨兰为材料通过调节培养基和添加外源激素进行根状茎诱导,以期筛选出最佳的培养基配方,提高其根状茎的繁殖系数和芽分化率,从而达到降低成本、提高效率的目的,有利于选育优良品种,促进墨兰种苗生产产业化。
1 材料与方法
1.1 材料
由福建连城兰花股份有限公司提供的墨兰‘吴字翠× 墨兰‘红花杂交种子无菌播种诱导出的根状茎为试验材料。
1.2 方法
1.2.1 6-BA、NAA和TDZ对墨兰根状茎增殖的影响 采用L9(34)正交试验研究不同浓度(2、5、8 mg/L)6-BA、(0.1、0.3、0.5 mg/L)NAA和(0.2、0.5、0.8 mg/L)TDZ对墨兰根状茎增殖的影响。所有处理附加1.5 g/L AC+35.0 g/L Su+ 6.0 g/L Ag(pH 5.6~5.8)。一共9组处理,每组接种20个材料为一个重复,每个材料长度为1.5~ 2.0 cm,3次重复。培养条件:光照时间12 h/d,光照强度1000~1500 lx,培养温度(25±1)℃。180 d后根据统计生长速度、增殖系数。
生长速度=(增殖后根状茎质量?接种时根状茎质量)/接种时根状茎质量
增殖系数=接种根状茎上新增的侧枝数/接种根状茎数(新增根状茎长度>0.5 cm)
1.2.2 高浓度6-BA和低浓度NAA、TDZ对根状茎叶芽分化的影响 采用L9(34)正交试验研究不同6-BA(8、10、12 mg/L)、NAA(0.3、0.5、0.7 mg/L)和TDZ(0.2、0.5、0.8 mg/L)对根状茎叶芽分化的影响。所有处理附加MS+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag(pH 5.6~5.8)。一共9组处理,每组接种20个材料为一个重复,每个材料长度为1.5~2.0 cm,2次重复。每组20个材料,2重复进行计算平均值,并对9组,每组2次重复的正交试验数据进行方差分析和多重比较分析。培养条件:光照时间12 h/d,光照强度1000~ 1500 lux,培养温度(25±1)℃。240 d后基于生长情况统计平均每个根状茎的叶芽分化率和株高。
叶芽分化率=根状茎上诱导出的芽个数/接种根状茎数×100%(芽>0.5 cm)
1.2.3 蛋白胨、椰子汁(CW)、活性炭(AC)对根状茎叶芽分化的影响 将材料采用L9(34)正交试验研究不同浓度蛋白胨(1、2、3 g/L)、活性炭(1.0、1.5、2.0 g/L)、椰子汁(50、100、150 mg/L)对根状茎的影响。基础培养基为MS+ 0.1 mg/L NAA+8 mg/L 6-BA+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag(pH 5.6~5.8)。一共9组,每组接种20个材料为一个重复,每个材料长度为1.5~ 2.0 cm,3次重复。培养条件:光照时间12 h/d,光照强度1000~1500 lx,培养温度(25±1)℃。240 d后记录生长情况并统计每个根状茎的叶芽分化率和株高。
叶芽分化率=根状茎上诱导出的芽个数/接种根状茎数×100%(芽>0.5 cm)
1.3 数据处理
使用Excel 2010软件计算平均值和标准差,SPSS 19.0统计分析软件对数据进行方差分析和Least- Significant Difference(LSD)多重比较,分析各因素对增殖和芽分化的影响。
2 结果与分析
2.1 6-BA、NAA和TDZ对根状茎增殖的影响
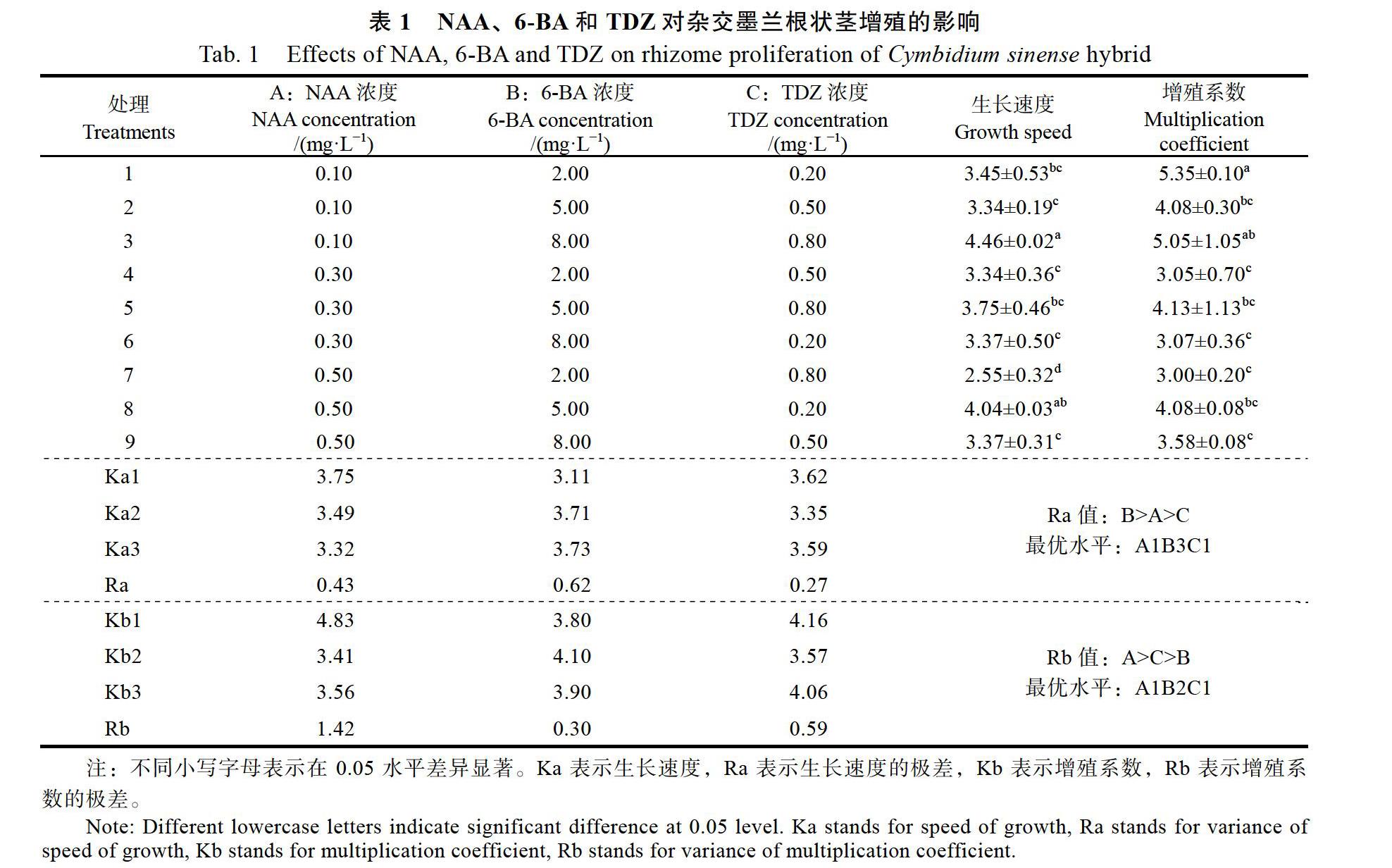
由表1可知,激素组合为8 mg/L 6-BA+ 0.1 mg/L NAA+0.8 mg/L TDZ(处理3)对根状茎生长速度的影响高于其他组合;当6-BA为2 mg/L,添加0.1 mg/L NAA+0.2 mg/L TDZ(处理1)时根状茎增殖系数最高,但添加0.5 mg/L NAA+0.8 mg/L TDZ(处理7)时增殖系数明显降低至3.00。
通过极差分析得出:在根状茎生长速度指标中,3种影响因素的主次关系为:6-BA>NAA> TDZ,6-BA浓度之间的关系5 mg/L 6-BA>8 mg/L 6-BA>2 mg/L 6-BA。在增殖系数的指标中,3种因素的主次关系为NAA>TDZ>6-BA,NAA浓度之间的关系是0.1 mg/L NAA>0.3 mg/L NAA> 0.5 mg/L NAA。最有利于根状茎生长的培养基为:8 mg/L 6-BA+0.1 mg/L NAA+0.2 mg/L TDZ;有利于提高根狀茎增殖系数的培养基为:5 mg/L 6-BA+0.1 mg/L NAA+0.2 mg/L TDZ。
由图1可知,根状茎接种在培养基上培养180 d后根状茎数量明显增加,并且新生根状茎有分支出现(图1)。处理1(图1A)根状茎粗壮、增殖数量较多。处理3(图1B)根状茎生长速度较高。处理7(图1C)根状茎生长速度和增殖的数量较低。
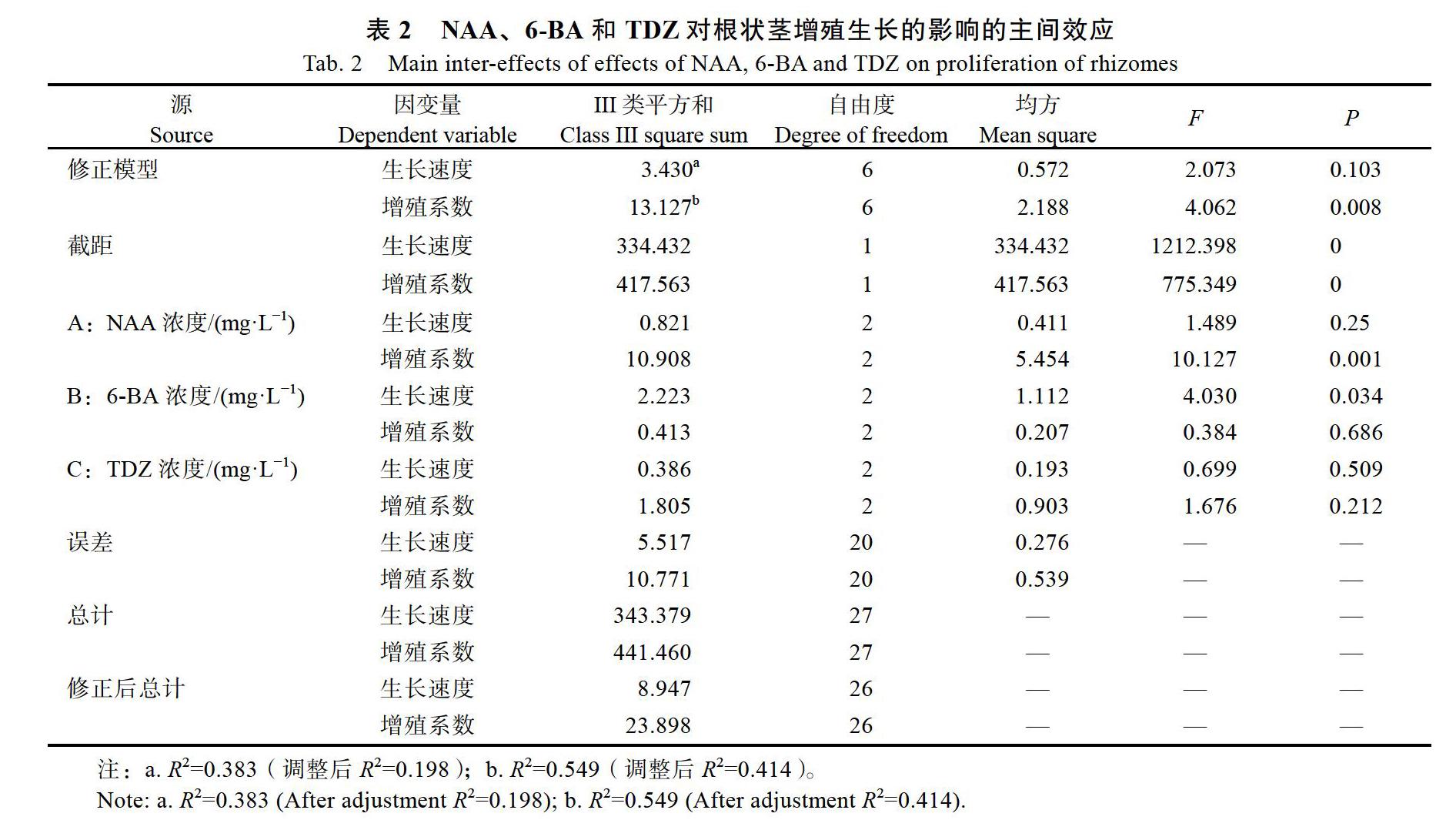
通过方差分析可知(表2),NAA和TDZ对根状茎生长速度的影响不显著。6-BA对根状茎生长速度具有显著性影响(P<0.05),LSD多重比较分析可知,2 mg/L 6-BA与5 mg/L 6-BA(P=0.026< 0.05)、2 mg/L 6-BA与8 mg/L 6-BA(P=0.021< 0.05)之间具有显著性影响,5 mg/L 6-BA与8 mg/L 6-BA(P=0.925)不具有显著性影响。NAA对根状茎增殖系数的影响具有显著性(P<0.01),6-BA和TDZ对增殖系数的影响不显著(P>0.05)。进一步进行LSD多重比较分析可知,0.1 mg/L NAA与0.3 mg/L NAA(P=0.001<0.01)、0.1 mg/L NAA与0.5 mg/L NAA(P=0.001<0.01)具有显著性差异,0.3 mg/L NAA与0.5 mg/L NAA(P= 0.688)无显著性差异。
2.2 6-BA、NAA和TDZ对根状茎叶芽分化的影响
根状茎接种在培养基中,暗培养30 d有明显侧枝生长(图2A),培养120 d后,根状茎进行转绿生长并有叶芽分化,并形成植株(图2B),培养180 d后芽数明显增加,并生长状态良好(图2C、图2D),到240 d后,植株高明显,芽诱导数明显提高(图2E)。
由表3可知,当6-BA为10 mg/L,添加0.2 mg/L TDZ和0.7 mg/L NAA时叶芽分化率最高,为52.50%,并且高于其他组合,即处理8;但添加0.8 mg/L TDZ和0.5 mg/L NAA时芽分化率仅次最高值,叶芽分化率为40%,即处理5。当6-BA降至8.0 mg/L时,组合为8 mg/L 6-BA+ 0.3 mg/L NAA+0.2 mg/L TDZ略低于处理5,处中间值叶芽分化率达37.50%。
由极差分析得出,适宜诱导根状茎叶芽分化及株高的组合为10 mg/L 6-BA+0.7 mg/L NAA+ 0.2 mg/L TDZ,叶芽分化影响因素的主次关系为:6-BA>TDZ>NAA,6-BA浓度之间的关系10 mg/L 6-BA>12 mg/L 6-BA>8 mg/L 6-BA。因此,当6-BA含量由小到大逐渐增大时,K值先增大后减小,说明墨兰6-BA对墨兰叶芽分化的影响先增大后减小。
通过NAA、6-BA和TDZ对叶芽分化率的主间效应检验可知(表4),NAA、TDZ对墨兰叶芽分化的影响不显著,6-BA对叶芽分化的影响具有显著性(P<0.05)。进行LSD多重比较分析可知,8 mg/L 6-BA和10 mg/L 6-BA(P=0.016<0.05)之间差异显著,8 mg/L 6-BA与12 mg/L 6-BA(P= 0.474)、10 mg/L 6-BA与12 mg/L 6-BA(P=0.059)之间差异不显著。
2.3 蛋白胨、椰子汁和活性炭对根状茎叶芽分化的影响
由表5可知,处理8的添加物组合为7 g/L蛋白胨+100 mL/L CW+1.0 g/L AC,叶芽分化率最高,达173.30%,并显著高于其他组合。处理2的添加物组合为1 g/L蛋白胨+100 mL/L CW+ 1.5 g/L AC,叶芽分化率最低,为26.70%。通过极差分析可知,适宜诱导芽分化的有机物配比为7 g/L蛋白胨+50 mL/L CW+1.0 g/L AC,3种因素之间的主次关系为AC>蛋白胨>CW,AC浓度之间的关系为1.0 g/L AC>2.0 g/L AC>0.5 g/L AC。因此,当AC含量由小到大逐渐增加时,K值先减小后增大,说明活性炭浓度对墨兰根状茎的影响先减小后增大。
杂交墨兰根状茎接种在培养基上,暗培养30 d后,根状茎有明显萌动,并且向下生长(图3A);接种培养90 d后,墨兰根状茎有明显芽分化,形成一定高度的植株(图3B);180 d后有大量植株形成,生长状况良好(图3C);并且在不同培养基中,处理8的叶芽分化率最高(图3F),处理2中的叶芽分化率最低(图3E),处理1的叶芽分化率处于中间值(图3D)。
通过蛋白胨、CW和AC对叶芽分化率的主间效应检验可知(表6),蛋白胨和活性炭对叶芽分化的影响具有显著性(P<0.05),CW对叶芽分化的影响不显著(P>0.05)。进行LSD多重比较分析可知,1 g/L蛋白胨与4 g/L蛋白胨(P=0.01< 0.05)、1 g/L蛋白胨与7 g/L蛋白胨(P=0.000< 0.001)和7 g/L蛋白胨与4 g/L蛋白胨(P=0.04< 0.05)之间存在显著性差异。1.0 g/L AC与2.0 g/L AC(P=0.000<0.05)、1.0 g/L AC与1.5 g/L AC(P=0.000<0.001)之间具有显著性差异,1.5 g/L AC与2.0 g/L AC(P=0.326<0.05)之间差异不显著。
3 讨论
组织培养是进行大量种苗繁殖的重要手段,一般包括诱导、增殖、分化、生根炼苗4个阶段。目前,关于墨兰组织培养的研究已经有许多相关报道,并主要以墨兰根状茎为试验材料。但墨兰不同品种、不同外植体部位之间差异很大,并且对培养基的组合、培养环境的要求都不尽相同。目前,在部分墨兰的组培繁殖研究中,以自交品种的根状茎为试验材料[2, 4],本研究以‘吴字翠× 墨兰‘红花杂交种子无菌播种诱导出的根状茎为试验材料,相对自交品种繁殖周期长,生长速度较慢,增殖系数较低,芽诱导率较低,但选用杂交品种的根状茎为材料有利于选育优良品种,促进墨兰产业化生产。因此,本研究通过对墨兰根状茎增殖分化的研究,探讨不同培养基对墨兰根状茎增殖分化的影响。
3.1 激素在墨兰根状茎增殖阶段的作用
组织培养增殖阶段对细胞分裂素的需求低于生长素,同时要保持生长素和细胞分裂素之间的平衡;大多数植物激素在调控植物生长发育过程中的作用比较复杂,同一种激素可调控影响多个发育过程,一个生长发育过程需要多种激素共同作用于植物[14]。相关研究表明,一定浓度范围6-BA(1~2 mg/L)、NAA(0.1~1.0 mg/L)组合有利于增殖,增殖系数达5.9[15]、7.4[2]、8.3[4]等,并且TDZ对根状茎增殖也有促进作用。刘昳雯等[16]提到激素配比为2.0 mg/L TDZ+0.3 mg/L NAA最适宜莲瓣兰‘滇梅(C. tortisepalum ‘Dian Mei)增殖。
本研究中6-BA、NAA和TDZ的激素配比有利于墨兰根状茎增殖,6-BA使用浓度与前人的研究有所不同,适宜浓度范围为2~8 mg/L,在后期生长起主要主导作用,其最适浓度为5.0 mg/L;NAA对根状茎增殖起主导作用,其浓度0.1 mg/L与以上结果接近;TDZ的作用不显著,并未取得较好的进展。与刘昳雯的结果不同,对此还需进一步实验确定。当2.0 mg/L 6-BA、0.1 mg/L NAA和0.2 mg/L TDZ组合配比时,增殖系数达5.35,这一结果进一步确定一定浓度6-BA、NAA和TDZ有利于根状茎增殖,虽然激素配比与丁雪珍[4]的结果接近,与其他品种研究结果相比较低,但高于左利娟等[3]的研究结果。这可能与不同品种之间存在差异有关,本研究杂交品种相较其他品种生长周期长、增殖系数低。
3.2 激素在墨兰叶芽分化阶段的作用
在葉芽分化的相关研究中,墨兰根状茎存在芽分化较低的问题,6-BA对叶芽的影响高于NAA,同时TDZ有利于芽分化,但其添加组合浓度各不相同,6-BA使用浓度范围较广为(1~ 10 mg/L),NAA为(0.5~1.0 mg/L)[5-12],叶芽分化率达0.8[11]、1.8[7]、2.9[10]、6.43[9]。
本研究中6-BA对墨兰叶芽分化起主要主导作用,6-BA浓度范围(8~10 mg/L)、NAA(0.3~ 0.7 mg/L)与以上研究接近,但与前人研究不同,6-BA浓度更广。当激素组合为10 mg/L 6-BA+ 0.7 mg/L NAA时叶芽分化率达52.50%,当激素组合为8 mg/L 6-BA+0.3 mg/L NAA时叶芽分化率达37.50%,研究结果与朱根发[8]的接近,这一结果表明,高浓度(8~10 mg/L)6-BA有利于叶芽分化,但可能与不同品种之间存在差异有关,并且对培养基的要求也不同,其叶芽分化率较低。
3.3 其他添加物对墨兰叶芽分化的作用
在相关研究中表明,100 mL/L CW能改变培养基中的渗透压提高蔗糖含量[9];于1/4 MS培养基中添加椰奶、马铃薯粉、蛋白胨和AC有利于花芽分化[17-18];加入活性炭会降低细胞分裂素的有效活性[19];陈云喜等[20]发现100 mL/L CW对芽分化有促进作用,活性炭对分化有抑制作用;陈兰芳[5]等表明培养基为MS+0.1 mg/L KT+1.0 mg/L NAA+100 ml/L CM+0.3 g/L活性炭,叶芽分化率达56%;Zeng等[21]提到添加1.0 ml/L NAA,1.0 g/L蛋白胨,10% CW和1.0 g/L AC有利于兜兰(Pap hiopedilum wardii Sumerh.)的芽培养;罗虹[22]指出活性炭明显抑制了墨兰分化,对增殖和生根壮苗阶段起促进作用。翁锦周等[23]研究发现MS+ 3.0 mg/L BA+0.5 mg/L NAA+4.0 g/L活性炭适合于不定芽分化。以上研究表明1.0 g/L AC和100 ml/L CW及1.0 g/L蛋白胨的添加有利于叶芽分化。
本研究结果表明1.0 g/L活性炭对叶芽分化起主导作用并具有显著性影响,蛋白胨对叶芽分化具有显著性影响,最适宜叶芽诱导的添加物组合7 g/L蛋白胨+100 mg/L CW+1.0 g/L AC,与Zeng等[21]结论接近,但蛋白胨浓度有所不同为7 g/L;本研究中CW对叶芽分化不具有显著性影响与陈云喜等[20]和陈兰芬等[5]研究结果不同,叶芽分化率高于陈兰芳的研究结果。过高含量的活性炭会抑制叶芽分化与罗虹[22]研究结果活性炭明显抑制叶芽分化类似。因此,一定浓度6-BA、NAA、活性炭和蛋白胨能够提高叶芽分化率,但过高含量的活性炭会抑制叶芽分化,会导致芽诱导数降低,活性炭的适宜浓度为1.0 g/L。
4 结论
本研究通过添加外源激素和其他添加物对杂交墨兰根状茎进行诱导繁殖,结果表明NAA对根状茎增殖影响具有显著性,且较高浓度6-BA(2.0~8.0 mg/L)有利于根状茎后期生长。最适宜根状茎增殖的培养基组合为MS+2 mg/L 6- BA+0.1 mg/L NAA+0.2 mg/L TDZ+1.5 g/L AC+ 35.0 g/L Su+6.0 g/L Ag,增殖系数达5.35。在杂交墨兰根状茎叶芽分化阶段,6-BA起主要影响作用,在一定范围浓度内6-BA(8.0~10.0 mg/L)有利于叶芽分化,最适宜叶芽分化的培养基为MS+10.0 mg/L 6-BA+0.7 mg/L NAA+0.2 mg/L TDZ+1.5 g/L AC+35.0 g/L Su+6.0 g/L Ag,叶芽分化率为52.5%。其他添加物对叶芽分化有影响作用,1.0 g/L活性炭起主导因素。杂交墨兰根状茎叶芽分化最适宜的培养基为MS+8.0 mg/L 6-BA+0.1 mg/L NAA+7.0 g/L蛋白胨+100.0 mg/L CW+1.0 g/L AC,叶芽分化率为173.30%,株高达3.58 cm。
参考文献
陈 丽, 潘瑞炽, 陈汝民. 墨兰原球茎生长的研究[J]. 热带亚热带植物学报, 1999(1): 59-64.
施福军, 莫昭展, 韦江萍, 等. 墨兰的无菌播种及根状茎的增殖研究[J]. 安徽农业科学, 2008, 36(32): 13968-13969, 13992.
左利娟, 汤久杨, 李志强. 墨兰杂交及F1代离体培养研究[J]. 北京农业职业学院学报, 2017, 31(3): 22-25.
丁雪珍, 韩 磊, 张文静. 墨兰增殖培养基的筛选研究[J]. 北方园艺, 2009(8): 208-209.
陈兰芬, 王 晶, 田亦平, 等. 墨兰组织培养根状茎分化技术研究[J]. 河北林果研究, 2011, 26(1): 22-24.
鲁雪华, 郭文杰, 林 勇. 墨兰的无菌播种和植株再生[J]. 亚热带植物科学, 1999(1): 34-37.
傅雪琳, 张志胜, 何 平, 等. 墨兰根状茎绿芽分化的研究[J]. 华南农业大学学报, 2000(3): 53-55.
朱根发, 叶秀斌, 陈明莉, 等. 培养基不同成分对墨兰根状茎分化成苗的影响[J]. 中南林学院学报, 2003(5): 42-44, 58.
许申平, 袁秀云, 王默霏, 等. 墨兰(Cymbiduim sinense)组培快繁技术体系研究[J]. 热带作物学报, 2018, 39(5): 926-930.
Yan X, Feng-an Y U, Zhen-hua P. Tissue culture of Cymbidium sinensis[J]. Forest Research, 2003, 16(4): 434-438.
Chen Y Y, Zhang Y, Zhang C, et al. A study on aseptic seed germination of interspecific hybrid between Cymbidium hybrida × C. sinense and C. faberi[J]. Acta Horticulturae Sinica, 2009, 36(3): 441-446.
Gao R, Wu S Q, Piao X C, et al. Micropropagation of Cymbidium sinense using continuous and temporary airlift bioreactor systems[J]. Acta Physiologiae Plantarum, 2014, 36(1): 117-124.
Chang C, Chang W. Effect of thidiazuron on bud development of Cymbidium sinense Willd in vitro[J]. Plant Growth Regulation, 2000, 30(2): 171-175.
徐 程, 詹忠根, 張 铭. 中国兰的组织培养[J]. 植物生理学通讯, 2002(2): 171-174.
郑艳艳. 墨兰组培体系的优化及根状茎在生物反应器培养的研究[D]. 延吉: 延边大学, 2011.
刘昳雯, 许春梅, 王 丹, 等. 莲瓣兰“滇梅”根状茎的增殖与分化技术研究[J]. 安徽农业科学, 2014, 42(10): 2890-2892.
Duan J X, Yazawa S. Floral induction and development in Phalaenopsis in vitro[J]. Plant Cell Tissue and Organ Culture, 1995, 43(1): 71-74.
Duan J X, Yazawa S. In vitro floral development in × Doriella Tiny (Doritis pulcherrima × Kingiella philippinensis)[J]. Scientia Horticulturae, 1994, 59(3-4): 253-264.
Duan J X, Yazawa S. In vitro flowering of Doriella, Phalaenopsis and Dendrobium[J]. Proceedings of NIOC, 1994: 87-96.
陈云喜, 何丹丹, 廖浩如, 等. 影响墨兰×兔耳兰根状茎芽分化的因素[J]. 中国农学通报, 2010, 26(9): 65-69.
Zeng S, Wu K, da Silva J A T, et al. Asymbiotic seed germination, seedling development and reintroduction of Paphiopedilum wardii Sumerh., an endangered terrestrial orchid[J]. Scientia Horticulturae, 2012, 138: 198-209.
罗 虹. 活性炭对墨兰根状茎生长的影响[J]. 广东农业科学, 1998(1): 30-31.
翁锦周, 林加耕, 林江波. 不同浓度活性炭对墨兰离体培养的影响[J]. 亚热带植物科学, 2006(3): 37-38, 36.
猜你喜欢
清华金融评论(2022年4期)2022-04-13
现代农业科技(2018年13期)2018-10-20
上海师范大学学报·自然科学版(2018年3期)2018-05-14
山东农业科学(2016年12期)2017-01-21
中国现代医生(2016年27期)2016-12-21
科技视界(2016年5期)2016-02-22
中国信息化周报(2014年4期)2014-02-17
文学教育·中旬版(2012年4期)2013-02-01
