
灰林鸮和北领角鸮在人工巢箱中的繁殖记录
2020-06-21 06:51李雨霖张建志彪杨
生物学通报 2020年8期
王 宁 李雨霖 孙 霄 张建志 刘 彪杨 军 陈 晨
(1 北京师范大学生命科学学院 北京 100875 2 北京市门头沟区西峰寺林场 北京 102308 3 北京百花山国家级自然保护区管理处 北京 102311)
灰林鸮(Strixaluco)和北领角鸮(Otussemitorques)是在我国分布比较广泛、栖息在森林中的2种的鸮形目(Strigiformes)鸱鸮科(Strigidae)鸟类[1-2]。国内对于这2种鸟类的繁殖研究很少,只有零星记录[3-5]。为进一步了解这2种猫头鹰的繁殖情况,为生物多样性监测和生态保护提供依据,于2015—2018年在北京百花山国家级自然保护区,开展了利用人工巢箱招引灰林鸮和北领角鸮繁殖的实验。
1 研究地区和研究方法
1.1 研究地区 实验区位于北京百花山自然保护区境内,面积848.8 ha,海拔1 100~1 600 m,G109国道从中间穿过。地貌为山沟、山脊相间,沟底有季节性溪流或水坑。植被类型为针阔混交林,主要树种有油松(Pinustabulaeformis)、华北落叶松(Larixprincipis)、山杨(Populusdavidiana)、青杨(P.cathayana)、绢柳(Salixviminalis)、白桦(Betula platyphylla)、棘皮桦(B.dahurica)、蒙古栎(Quercusmongolica)、核 桃 楸(Juglansmandshurica)、元宝槭(Acertruncatum)、春榆(Ulmuspropinqua)等。
1.2 研究方法
1.2.1 安放巢箱 参考欧洲鸮形目鸟类巢箱的尺寸[6-7]制作了2种规格的巢箱(图1),大型巢箱用于招引灰林鸮,小型巢箱用于招引北领角鸮。
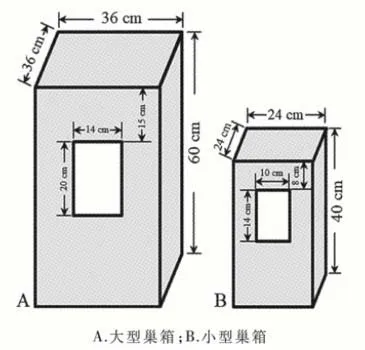
图1 用于招引鸮类繁殖的巢箱
选择每条山沟沟底较为开阔的区域,用铁丝将巢箱固定在树干上,距地面约5 m,箱口朝向沟底背风的方向,箱口周围无树枝阻挡鸮类飞行、进出巢箱。巢箱底部铺垫约5 cm厚的锯末和干枯树叶作为巢材。每条山沟安放2~4个巢箱,大、小巢箱各占一半,巢箱间距离大于100 m。
2015年2月底安放10个大巢箱,10个小巢箱;2015年12月初又安放20个大巢箱,20个小巢箱。2016—2018年未安放新巢箱,巢箱总数不变。
1.2.2 繁殖监测 2015年4月初检查巢箱时,发现灰林鸮已产卵,未能记录到产卵日期。2015年12月,在部分巢箱的箱口附近安装了红外线触发相机,监测鸮类进出巢箱的情况。2016—2018年,每年3月初开始检查巢箱。发现灰林鸮在产卵前1~2 d的白天已在巢箱中停留,产卵后白天几乎都在巢箱中孵卵,当人检查巢箱时会飞离。北领角鸮产卵后,人检查巢箱时并不从巢箱中飞离,甚至触碰其羽毛也不飞离,无法观察到被其身体覆盖的卵及雏鸟的情况。为了避免因检查巢箱造成的干扰导致繁殖失败,每隔约1周检查一次巢箱。因此,只有少数繁殖的巢箱记录到了准确的产卵日期和雏鸟孵化日期,通过红外相机记录了部分巢箱内幼鸟离巢的日期。
1.2.3 数据测量与分析 利用GPS定位仪结合Google Earth软件测量同一年中被利用的巢箱之间的距离、被利用的巢箱与公路间的距离。将产卵日期换算成距离3月1日的天数,用于比较2种鸮形目鸟类之间产卵日期的早晚。利用有准确记录的产卵日期、雏鸟孵化日期和幼鸟离巢日期,计算孵卵期和育雏期,同一窝中,以第1枚产出的卵孵出雏鸟的日期计算孵卵期,以第1只雏鸟离巢的日期计算育雏期。每巢的孵化率等于巢中孵出的雏鸟数除以窝卵数。卵的大小用长径和短径表示。用统计软件SPSS for Windows 20.0比较2种鸮的各项数据,由于样本量较小,使用非参数检验进行比较。每项数据的结果用平均值±标准误(样本量,最小值~最大值)表示。
2 结果
2.1 利用巢箱繁殖的鸮形目鸟类数量 2015年2月底悬挂巢箱后,当年4月初即发现有灰林鸮在巢箱中产卵,4月底发现有北领角鸮在巢箱中产卵。从2015年至2017年,灰林鸮的繁殖巢数逐渐上升(图2),繁殖密度达到8.2对/10 km2,2018年则与2017年持平。北领角鸮各年的繁殖巢数不稳定(图2),繁殖密度在1.2~5.9对/10 km2。2015—2018年,未发现其他鸮形目鸟类在巢箱中繁殖。

图2 2015—2018年灰林鸮和北领角鸮在巢箱中繁殖的数量
2.2 对巢箱的利用
2.2.1 对不同型号巢箱的选择 灰林鸮从12月开始进出巢箱,直至第2年产卵,并非所有进出过的巢箱都在其中繁殖。2016年用红外相机监测的19个巢箱中,灰林鸮进出过17个,最终利用繁殖的3个,选择率为17.6%。北领角鸮从当年3月开始有进出巢箱的现象。灰林鸮和北领角鸮都既利用大巢箱,也利用小巢箱,对大巢箱的利用多于小巢箱(表1)。
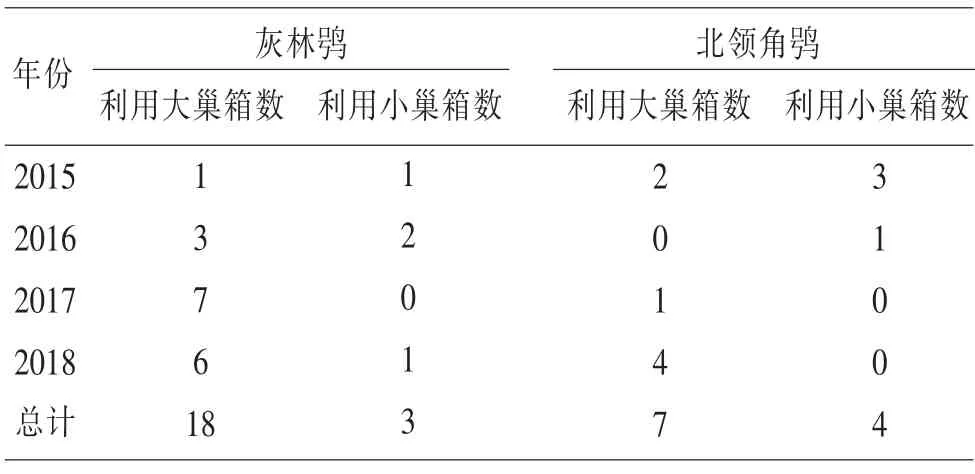
表1 2015—2018年灰林鸮和北领角鸮对巢箱大小的选择数量(单位:个)
2.2.2 对不同区域巢箱的利用 同一年中,灰林鸮利用的巢箱分别位于不同的沟里(图3),灰林鸮利用的巢箱间距为(2 012±130)m(n=53,618~3 995 m)。2017年和2018年,各有一条沟里的2个巢箱分别被北领角鸮和灰林鸮利用,其余各年北领角鸮利用的巢箱与灰林鸮利用的巢箱都不在同一条沟里。2015年有3个在同一条沟里的巢箱被北领角鸮利用,其余各年北领角鸮利用的巢箱都不在同一条沟里,北领角鸮利用的巢箱间距为(1 064±140)m(n=16 119~1 976 m)。北领角鸮利用的巢箱间距显著小于灰林鸮利用的巢箱间距(Mann-Whitney U检验,Z=-3.392,P=0.001)。

图3 2015—2018年被灰林鸮和北领角鸮利用的巢箱位置
被灰林鸮利用的巢箱至公路的距离为(333±59)m(n=21,71~911 m)。被北领角鸮利用的巢箱至公路的距离为(338±78)m(n=11,71~747 m)。2种鸮利用的巢箱到公路的距离没有显著差异(Mann-Whitney U检验,Z=-0.000,P=1.000)。
2.2.3 巢箱重复利用的情况 有6个巢箱分别被灰林鸮重复利用2~3次;1个巢箱被北领角鸮重复利用2次;1个巢箱被北领角鸮利用后,在之后的2年被灰林鸮利用;2个巢箱被灰林鸮利用后,在之后的1年被北领角鸮利用。
2.3 繁殖参数
2.3.1 产卵日期 灰林鸮3月产卵,产卵日期距3月1日的天数为(8.1±1.1)d(n=12,4~16 d)。多数北领角鸮在4月产卵,产卵日期距3月1日的天数为(53.9±4.1)d(n=8,45~82 d)。灰林鸮的产卵日期显著早于北领角鸮(Mann-Whitney U检验,Z=-3.317,P<0.001)。
2.3.2 窝卵数 灰林鸮的窝卵数为(2.9±0.2)个(n=21,1~5个),北领角鸮的窝卵数为(4.8±0.5)个(n=9,3~8个)。灰林鸮的窝卵数显著少于北领角鸮的窝卵数(Mann-Whitney U检验,Z=-3.280,P=0.001)。
2.3.3 卵大小 灰林鸮和北领角鸮的卵均为白色短椭球型。灰林鸮的卵大小为:长径(46.3±0.2)mm(n=34,43.9~48.4 mm),短径(38.5±0.1)mm(n=34,30.6~40.0 mm)。北领角鸮的卵大小为:长径(37.2±0.2)mm(n=27,35.5~38.7mm),短径(31.0±0.2)mm(n=27,29.7~32.9 mm)。灰林鸮的卵显著大于领角鸮的卵(Mann-Whitney U检验:卵长径,Z=-6.669,P<0.001;卵短径,Z=-6.668,P<0.001)。
2.3.4 孵卵期 灰林鸮和北领角鸮都是在产第1枚卵后即开始孵卵,后面的卵隔日或次日产出,同一窝的卵不在同一天孵出雏鸟,最先产出的卵最先孵出雏鸟,为异步孵化。灰林鸮的孵卵期为(32.7±0.7)d(n=3,32~34 d),北领角鸮的孵卵期为(30.0±2.0)d(n=4,27~36 d)。
2.3.5 孵化率 灰林鸮卵的孵化率为(81.4±7.1)%(n=17,0~100个),北领角鸮卵的孵化率为(60.2±12.0)%(n=7,0~100个),种间差异不显著(Mann-Whitney U检验,Z=-1.742,P=0.082)。刚孵出的雏鸟体表有白色绒毛,头不能抬起,眼睛也不能睁开。
2.3.6 育雏期 灰林鸮的育雏期为(29.5±0.8)d(n=6,27~32 d)。只有2窝北领角鸮记录了准确的育雏期,分别为23 d、29 d。幼鸟离巢时,飞羽和尾羽还未完全长好,身体的羽毛大部分是灰色的绒羽,还不能飞行。幼鸟从巢箱中跳至地面后,用双足从树干爬上树冠,隐藏在枝叶中,仍需亲鸟喂食。
2.3.7 繁殖失败的原因 松鼠(Sciurusvulgaris)会在2种鸮产卵前占据巢箱,在其中筑巢进行冬眠和繁殖。松鼠的巢用树枝构筑,树枝伸出巢箱口,将巢箱口遮挡,鸮类无法进入巢箱繁殖。未发现灰林鸮的卵、雏被天敌捕食的现象。红外相机记录到灰林鸮攻击巢箱中的北领角鸮,还记录到了赤峰锦蛇(Elapheanomala)捕食巢箱中的北领角鸮雏鸟。
3 讨论
利用人工巢箱招引灰林鸮和北领角鸮繁殖获得了成功,但每年利用巢箱繁殖的灰林鸮和北领角鸮都不多,可能是作为位于食物链的上层或顶端的夜行性猛禽,灰林鸮和北领角鸮在自然界的密度本身较低,而试验区的面积较小,鸮类种群数量有限。
2015—2017年,在实验区内人工巢箱中繁殖的灰林鸮数量逐渐上升,而2018年则与2017年持平,可能是灰林鸮的繁殖密度已经达到了饱和。灰林鸮在欧亚地区分布较广,在欧洲的森林中,灰林鸮的繁殖密度为2~20对/10 km2[8]。在本研究中,灰林鸮的繁殖密度达到8.2对/10 km2,繁殖密度处于已知的范围内。灰林鸮体长近40 cm,需要较大的树洞繁殖,但灰林鸮对巢址的选择比较宽泛,树洞缺乏时,灰林鸮会选择山崖陡坡的石洞、其他鸟类废弃的用树枝搭成的巢、甚至地面的凹坑繁殖[9]。灰林鸮适应在巢箱中繁殖,特别是在缺少大树的中、幼龄森林中,安放人工巢箱后,灰林鸮几乎都会在人工巢箱中繁殖[10]。灰林鸮在我国的分布较为广泛,以北京为例,根据观鸟记录,从近郊的香山、八大处、百望山等低山丘陵地带,到远郊的百花山、东灵山、海坨山等中山区,均发现过灰林鸮繁殖。在北京地区,山区森林大多是中、幼林,缺乏适于灰林鸮繁殖的较大的树洞。本研究区安放人工巢箱前,灰林鸮是很少见的。2014—2015年,连续发现灰林鸮在同一个陡坡的石洞里繁殖,推测可能缺乏适合繁殖的树洞[5]。安放人工巢箱后,灰林鸮逐渐成为比较常见的鸟类,经常能听到叫声。因此,在北京山区森林中安放一定数量适合灰林鸮繁殖的巢箱,会增加灰林鸮的密度,对于控制森林鼠害有积极的作用。本研究中的灰林鸮繁殖参数,与欧洲灰林鸮的繁殖参数大致 相 当[9]。
在本研究中,利用巢箱繁殖的北领角鸮和灰林鸮数量呈现出一定的消长变化(图2)。北领角鸮选择繁殖的巢箱多数位于当年没有灰林鸮繁殖的沟里,极少数与当年繁殖的灰林鸮巢箱在同一条沟里时,北领角鸮选择的巢箱位于沟的更深处。结合红外相机记录到的灰林鸮攻击北领角鸮的现象,可推测北领角鸮受到了灰林鸮的竞争排斥。北领角鸮体长约24 cm,体型比灰林鸮小得多,在种间竞争中处于劣势。在欧洲,灰林鸮对于体型较小的其他鸮类有竞争排斥作用[11-13],甚至捕杀小型鸮类巢中的亲鸟和雏鸟[14]。本实验中,北领角鸮利用巢箱繁殖的特点,可能反映了对来自灰林鸮的竞争排斥作用的规避。根据红外相机的记录,3月中旬后,北领角鸮才在研究区出现,北领角鸮在低山平原地带过冬,可能是季节性垂直迁移至山区繁殖。北领角鸮抵达繁殖区时,灰林鸮已产卵,领域相对固定,推测北领角鸮选择灰林鸮活动较少的区域进行繁殖。本研究发现,北领角鸮利用的巢箱间距显著小于灰林鸮利用的巢箱间距。在没有灰林鸮繁殖的沟里,北领角鸮繁殖的巢箱间距只有约100 m。在同一条沟中繁殖的北领角鸮和灰林鸮的巢箱间距只有约400 m。推测北领角鸮的领域是较小的,可利用灰林鸮领域之间的空隙作为自己的领域,完成繁殖。在同一年中,北领角鸮的繁殖期比灰林鸮的繁殖期晚,随着季节推后,昆虫等猎物增加,可能减少了北领角鸮和灰林鸮在猎物选择上的竞争,这需要今后对二者的猎物组成进行分析,方能确证。
致谢
感谢北京师范大学生命科学学院研究生张敬刚、周冰、史洁、李扬、陈功、赵凯、刘逸侬等协助野外工作。
猜你喜欢
学与玩(2022年9期)2022-10-31
疯狂英语·新悦读(2021年5期)2021-11-28
疯狂英语·新阅版(2021年5期)2021-06-21
文苑(2020年12期)2020-04-13
中国交通信息化(2019年4期)2019-07-13
小太阳画报(2019年1期)2019-06-11
发明与创新·中学生(2018年11期)2018-11-30
小学生导刊(低年级)(2016年5期)2016-05-27
海峡姐妹(2016年3期)2016-02-27
校园足球(2010年8期)2010-01-24
