
转基因大豆向野生大豆基因漂移研究进展
2020-07-27 11:54刘来盘
生态与农村环境学报 2020年7期
刘 标,薛 堃,刘来盘,周 延,韩 娟
(1.生态环境部南京环境科学研究所,江苏 南京 210042;2.中央民族大学生命与环境科学学院,北京 100081;3.南京林业大学生物与环境学院南方现代林业协同创新中心,江苏 南京 210037;4.农业农村部食物与营养发展研究所,北京 100081)
1996年,转基因作物的全球种植面积为170万hm2,2017年扩大到1.898亿hm2,其中转基因大豆的种植面积最大,为9 410万hm2[1]。转基因作物的商业化种植不仅产生了巨大的经济效益,而且通过降低化学杀虫剂和除草剂用量保护了环境和人体健康[2]。但是,转基因作物也可能产生环境和人体健康等方面的风险,其中,外源基因漂移或基因流(gene flow)是最主要的环境风险之一,尤其是如果外源基因通过花粉介导的基因漂移从转基因作物逃逸到作物的野生近缘物种及其带来的潜在环境风险,更是引起了广泛关注[3-7]。
基因漂移原本是群体遗传学中的概念,指一个或多个基因从一个孟德尔遗传群体转移到另一个孟德尔遗传群体的现象或者过程[8]。根据这个定义,基因漂移是可以发生在一个物种的不同群体之间和具有不同亲缘关系的物种之间。事实上,同一作物的不同品种之间、作物与其野生近缘种和杂草类型之间进行基因漂移和遗传物质的交换是植物(如农作物)进化过程中存在了数千年的自然现象[9]231-232,[10]。世界上的主要农作物中有12种可以与其野生近缘种杂交,并对野生近缘种的进化产生重要影响:从7种作物到其野生近缘种的基因漂移导致其野生近缘种的杂草性增强(more aggressive weeds),或者在特定情况下导致某些稀有野生近缘种灭绝(extinction)[9]217,[11-14]。目前,认为转基因植物通过花粉逃逸是最普遍的基因漂移方式,即转基因作物通过花粉传播与其性亲和的野生近缘种进行杂交,从而逃逸到自然环境中。在花粉传播过程中,外源基因通过亲代遗传给子代,所以此类发生在同种或相近种之间的基因漂移也叫作垂直的基因漂移[15]。 转基因作物向其野生近缘种基因漂移的风险评价和研究主要包括基因漂移频率(异交率)、外源基因在杂交后代中的表达以及杂交后代的适合度评价[10,16-17]3个步骤。
已经商业化释放的转基因生物需要持续监测,处于中间阶段的转基因生物则需要更加广泛和深入的研究。世界范围种植面积最大的转基因作物是转基因大豆,即以大豆属Soya亚属中的栽培大豆〔Glycinemax(L.)Merr.〕为受体的转基因品种,而Soya亚属有2个种,另一个种是栽培大豆〔G.max(L.)Merr.〕的野生近缘种野生大豆(GlycinesojaSeib. et Zucc.)[18]。两者有相同的基因组(染色体均为2n=40),容易杂交,结实性良好,相互之间易于发生基因漂移,而且杂交后代性状的遗传方式与栽培大豆品种间杂交后代的遗传方式相似。野生大豆是一年生自花授粉草本植物,广泛分布于中国大陆、台湾岛、日本、朝鲜和俄罗斯西伯利亚等地,是我国第一批重点(二级)保护野生植物。我国是世界野生大豆最主要的分布和分化中心,主要分布于山野以及河流沿岸、湿草地、湖边、沼泽附近或灌丛等自然环境中,在农田及其周边环境中也可正常生长[19-20]。随着我国大量进口转基因大豆以及自主研发的多个转基因大豆品种具备商业化种植的技术条件,转基因大豆基因漂移风险,尤其是外源基因向我国野生大豆发生基因漂移及其对野生大豆种质资源可能产生的影响,成为转基因生物安全研究中的重要课题。
1 转基因作物向其野生近缘种的基因漂移
转基因通过基因漂移转移到野生近缘种的群体,不仅可能影响野生近缘种的遗传完整性(genetic integrity),还可能通过改变后者的生存和繁殖能力而使野生近缘种迅速扩展分布空间,甚至形成难以控制的杂草,产生难以预测的环境风险[3,10,17,21-22]。随着转基因作物商业化种植和贸易规模的逐渐扩大,多种转基因作物中外源基因漂移的现象已经被证实。多种转基因抗除草剂油菜(BrassicanapusL.)在加拿大进行商业化种植之后,抗不同除草剂的转基因油菜之间发生的基因漂移不仅导致加拿大转基因油菜田及其周围环境中出现了同时含有抗草甘膦、抗草丁膦和抗咪唑啉类3种除草剂的油菜自生苗[23-24],而且转基因油菜与油菜的杂草型野生种芜菁(BrassicarapaL.)之间发生基因漂移,使得外源抗除草剂基因进入并长期存在于芜菁自然群体中[25-26]。阿根廷也出现了转EPSPS基因油菜(B.napusL.)向芜菁(B.rapaL.)发生基因漂移并导致表达EPSPS基因的抗草甘膦芜菁在农田存在和蔓延的报道[27]。墨西哥是世界玉米的生物多样性起源中心,随着转基因玉米在美国(特别是靠近墨西哥的边界地区)的大规模种植以及出口到墨西哥,墨西哥的地方玉米品种中已经被发现含有转基因玉米的外源基因成分[28-30]。转Bt基因棉花(Gossypiumhirsutum)于1996年被批准在墨西哥进行商业化种植,2011年发表的论文中确认了墨西哥的陆地棉野生群体中检测到Bt基因[31]。在中国,Bt和EPSPS等外源基因可以通过基因漂移从转基因水稻转移到栽培稻(OryzasativaL.)、杂草稻(O.sativaf.spontanea)和普通野生稻(O.rufipogonGriff.)中并在杂交后代中正常表达[8,32]。
总之,大量研究和调查结果表明,只要能够杂交而且在时间和空间上具备杂交条件,转基因作物中的外源基因可以通过基因漂移转移到其野生近缘种并在杂交后代中正常表达[8,33-34]。
2 转基因大豆与野生大豆之间的基因漂移
2.1 栽培大豆与野生大豆之间的基因漂移
不同品种栽培大豆之间发生杂交的频率通常小于3%,隔行种植情况下的自然异交率为0.65%~6.32%,平均自然异交率为1.8%,超过10 m的自然异交率低于0.01%;栽培大豆之间主要通过昆虫(如蜜蜂)传粉进行杂交,风媒传粉几乎可以忽略不计[35-39]。
栽培大豆与野生大豆之间的异交率一般低于栽培大豆之间的异交率。日本栽培大豆与野生大豆在相隔50 cm、约30 d花期重叠条件下平均自然异交率为0.73%[40],KURODA等[41]发现6.8%的受检日本野生大豆植株中含有来自栽培大豆的基因。栽培大豆与野生大豆的杂交后代可以在自然条件下长期生存[42-44]。分子生物学和表型证据都表明,栽培大豆自从诞生以来就同野生大豆在重叠分布区域共同生存至少有5 000 a的历史并发生了持续性的遗传渗透。栽培大豆可以通过花粉而将其基因传递给野生大豆,即栽培大豆的基因已经渗透到野生近缘种,导致现代野生大豆含有栽培大豆的某些基因(野生大豆已经渗入0.73%的栽培基因),野生大豆的某些稀有性状(如白花、灰毛以及黄、绿、棕色种皮)可能来源于栽培大豆[41,45-49]。刘亚男[50]对26个省份的96份野生大豆种质材料的研究结果表明,有些野生大豆个体含有来自栽培大豆的基因或遗传成分。切叶蜂(Megachiletsurugensis)等访花昆虫是栽培大豆向野生大豆基因漂移的主要媒介[40,43-44,51-52]。
不同野生大豆个体和群体之间也存在基因漂移。日本学者研究结果表明,由于意大利蜂(Apismellifera)、木蜂(Zylocopaappendiculata)等传粉昆虫经常性地访问野生大豆的花,日本4个野生大豆群体间的多位点异交率(multilocus outcrossing rate)可达9.3%~19%,而且野生大豆群体内部的基因漂移频率高于栽培大豆与野生大豆之间的基因漂移频率[53-54]。中国野生大豆群体内以及相邻的不同种群之间也存在着基因漂移,野生大豆自然群体间的异交率为0~3.5%,而且在50 km范围内,不同野生大豆群体间的距离越近,其基因漂移频率越高[55-58]。
可见,栽培大豆与野生大豆之间、野生大豆不同群体之间都会发生低频率的基因漂移,昆虫是基因漂移的主要媒介。如果转基因大豆在我国进行商业化种植,外源基因可以通过基因漂移进入野生大豆,并可能在不同野生大豆群体间进一步扩散。
2.2 转基因大豆向非转基因大豆和野生大豆的基因漂移
为了研究和评价转基因大豆的基因漂移风险,国内外学者从转基因大豆向非转基因大豆的基因漂移以及转基因大豆向野生大豆的基因漂移2个方面开展了一些大田试验(表1[20,40-41,44,53-54,59-66])。
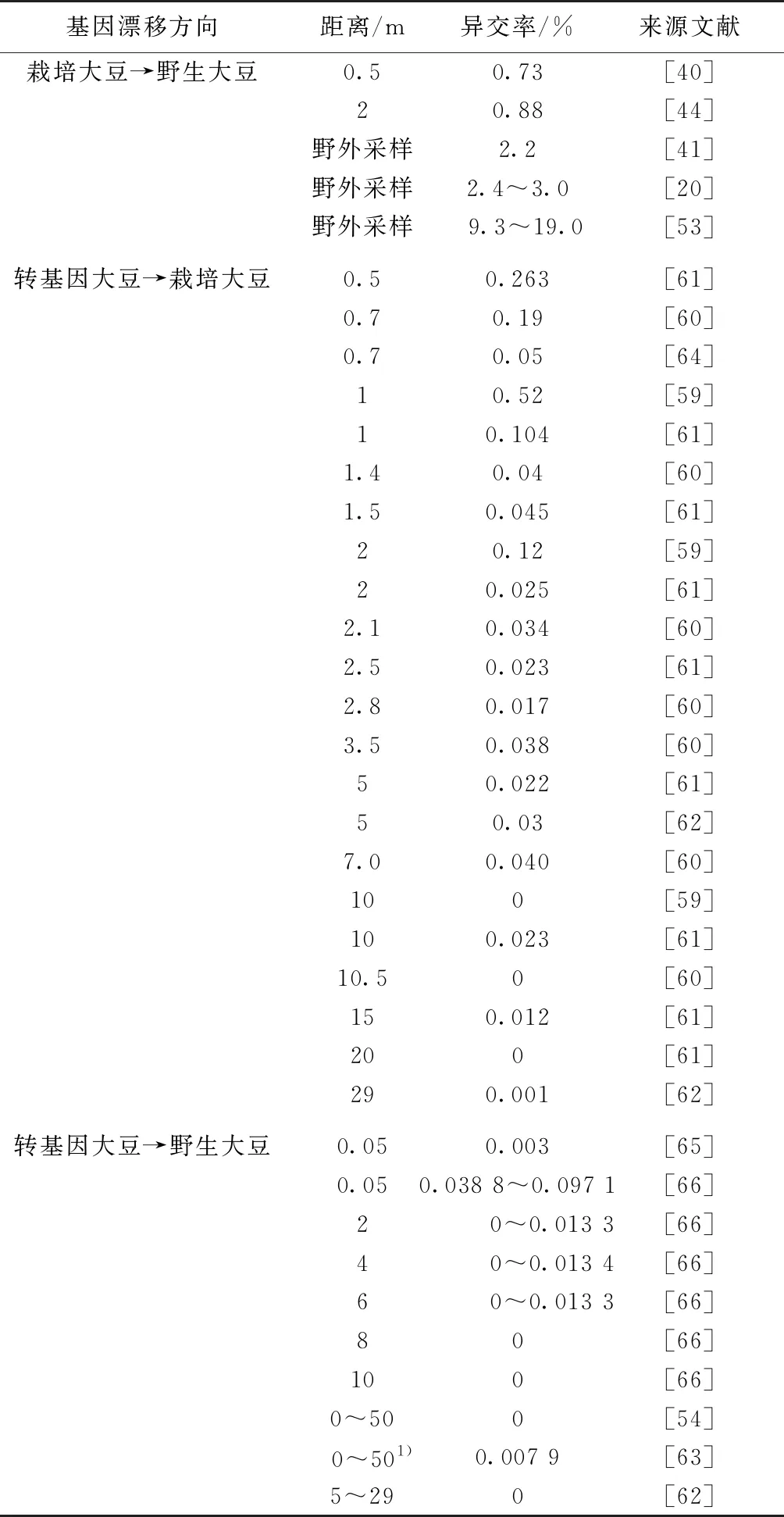
表1 不同大豆材料间基因漂移的异交率
在巴西开展的一个转基因大豆与栽培大豆基因漂移的大田试验中,当非转基因大豆距离转EPSPS基因大豆花粉供体1 m时,平均基因漂移频率为0.52%,2 m处的平均基因漂移频率为0.12%,当距离增至10 m时未检测到基因漂移[59]。日本学者开展的一个为期4 a的试验结果表明,当转EPSPS基因大豆AG3701RR(Event 40-3-2)与非转基因大豆相距0.7 m时,两者之间的基因漂移频率最高(0.19%),相距10.5 m处检测不到基因漂移[60]。在中国的隔行种植以及同心圆种植试验中,转EPSPS基因抗除草剂大豆AG5601向36个常规非转基因大豆品种的自然异交率为0.039%~0.934%,最远漂移距离15 m处的异交率为0.012%[61]。刘杰等[62]的研究结果表明,转EPSPS基因大豆和不同品种非转基因大豆之间相距5 m处基因漂移频率为0.03%,而在29 m处降至0.001%。中国和日本的试验均发现,转基因大豆与常规大豆之间主要通过传粉昆虫发生基因漂移,通过风力传播花粉而发生基因漂移的概率极低[60-61]。
已经发表的转基因大豆与野生大豆之间的基因漂移研究主要由中国和日本学者完成。陈新等[67]发现转EPSPS基因大豆ARG04与野生大豆之间的基因漂移频率约为0.09%,昆虫是基因漂移的主要媒介;陈新等[63]的另外一个试验结果显示:转EPSPS基因大豆ARG04与野生大豆之间最远漂移距离为10 m,与野生大豆之间的漂移频率低于万分之一。也有一些大田试验则未检测到转EPSPS基因大豆向野生大豆的基因漂移[62,64]。MIZUGUTI等[65]在日本开展的基因漂移试验中,转基因大豆AG3701RR与野生大豆的花期重合期为18~24 d,虽然32 502个F1大豆苗中只有1株是转基因阳性,但是这株杂交转基因大豆可以产生947粒种子,其中824粒种子可以发育为正常植株。MIZUGUTI等[66]的另外一个研究结果表明,转基因大豆与野生大豆间杂交频率(hybridization frequencies)为0~0.097%,且转基因大豆与野生大豆的花期重合度越高,两者之间的杂交频率也越高。
由上述文献可知,虽然基因漂移频率较低,但转基因大豆是可以向栽培大豆和野生大豆发生基因漂移的,而且杂交后代可育。如果转基因大豆中的外源基因进入野生大豆中,分析和评价转基因为野生大豆带来的适合度效应,对于转基因大豆基因漂移风险评价以及野生大豆种质资源保护至关重要[8,17]。
3 转基因大豆与野生大豆杂交后代的适合度
转基因作物与野生近缘种杂交后代的适合度分析,是评价转基因漂移引起的生态风险的重要步骤[17,68-69]。虽然KURODA等[70]认为栽培大豆与野生大豆杂交后代中带有来自栽培大豆的一部分人工驯化基因,可能不利于杂交后代在自然环境中生存,使得杂交后代的适合度可能低于野生大豆,但是,到目前为止仅有2个公开发表的相关研究。
GUAN等[71]通过人工授粉获得转EPSPS基因大豆与野生大豆的杂交F1代种子,然后在温室和大田条件下研究F1和F2代的适合度,发现虽然F1代结荚率很低,但是含有EPSPS基因的F1大豆比不含EPSPS基因的F1大豆有更长的营养生长期、更高的生物量和更低的百粒重,F1和F2的百粒重显著高于野生大豆,表明EPSPS基因没有对含有EPSPS基因杂交后代的生长产生不利影响,由此推测逃逸的EPSPS基因可以在没有草甘膦除草剂的自然条件下存在于野生大豆群体中。阚贵珍等[72]在网室条件下开展的研究结果表明,4个野生大豆材料和转EPSPS基因大豆RR能够杂交结实并产生有抗草甘膦性状的杂交后代群体F1和F2,在没有草甘膦选择压力条件下,杂交后代在一些性状上的相对适合度高于母本野生大豆材料(尤其是结荚率、百粒重、单株种子质量等繁殖指标),其他适合度指标没有发生显著差异。上述2个实验结果显示,在没有草甘膦的条件下,与母本野生大豆相比,表达EPSPS基因的“转基因大豆-野生大豆”杂交后代没有显示出适合度代价,某些适合度参数(尤其是繁殖指标)甚至显著高于野生大豆,表明表达EPSPS基因的“转基因大豆-野生大豆”杂交后代可能具有更强的环境入侵性,这与KURODA等[70]的预测结果并不相符,需要对此进行更加系统而深入的研究。
4 转基因大豆向野生大豆基因漂移研究中存在的问题
综上所述,虽然目前国内外已经有了一些转基因大豆向野生大豆基因漂移的研究报道,但是该领域的研究还存在着一些不足和问题,主要包括以下3个方面。
首先,没有考虑我国不同地域野生大豆群体之间存在的巨大遗传差异。除青海、新疆、海南3省(区)以外,野生大豆在我国其余各省(区、市)均有分布。巨大的生态、地理和气候条件差异使得我国不同地域的野生大豆形成了适应当地环境的各种生态型和种群。不同生态型野生大豆在叶型、花色、绒毛色、种子大小、种皮色、茎形态、生育期等表型以及遗传多样性方面存在很大差异,尤其是不同生态型野生大豆在生育期(特别是花期)、花色和种子等性状方面的差异将直接影响其与转基因大豆间基因漂移的频率以及杂交后代在不同环境条件下的生存和适合度[49,73]。现有的转基因大豆向野生大豆基因漂移研究仅仅涉及少数几个地域的野生大豆,没有考虑我国不同地域野生大豆之间存在的巨大遗传差异可能导致转基因大豆在不同地域向野生大豆发生基因漂移的风险等级存在很大不同,如转基因大豆向不同地域野生大豆基因漂移的频率可能有很大差异,杂交后代在不同地域的不同土壤、温度等环境条件下的适合度表现也可能有很大差别,从而导致不同的环境风险。
其次,已经研究的转基因大豆所涉及的外源基因集中于EPSPS基因,只有少量涉及其他外源基因(如Bt基因)以及基因叠加性状。除了EPSPS基因外,我国已经大量进口的转基因大豆以及自主研发的转基因大豆所表达的外源基因还包括能够降解2,4-D类除草剂的芳氧基链烷酸酯双加氧酶-12基因(aad-12)、使草铵膦除草剂失活的膦丝菌素乙酰转移酶基因(pat)、对鳞翅目害虫具有毒杀作用的Bt基因、抗麦草畏除草剂的麦草畏单加氧酶基因(dmo)以及复合性状转基因大豆所含有的多个外源基因(如陶氏益农公司研发的转基因大豆DAS-44406-6含有 2mEPSPS、aad-12 和pat3个外源基因)等。不同的外源基因以及外源基因组合如果通过基因漂移进入野生大豆基因组,所产生的适合度效应和环境风险是不同的,应根据个案原则(case-by-case principle)分别对其风险进行评价和研究。
第三,转基因大豆向野生大豆发生基因漂移所产生杂交回交后代的适合度研究不足。转基因大豆向野生大豆发生基因漂移所产生的危害性取决于外源基因在杂交后代中的表达以及杂交后代的适合度[17,74]。但是,目前公开报道的相关研究案例只有2个,而且研究内容中没有涉及杂交后代在不同环境条件下的外源基因表达、生育期、休眠性和落粒性变化等重要内容,更没有考虑外源基因还可能通过与野生大豆回交在不同野生大豆群体间扩散所带来的风险。另外,我国转基因大豆环境安全评价技术标准(农业部2031号公告-3-2013 转基因植物及其产品环境安全检测 耐除草剂大豆 第3部分:外源基因漂移)中只规定需要评价转基因大豆向栽培大豆和野生大豆基因漂移的距离和频率,没有涉及转基因大豆向野生大豆发生基因漂移所产生杂交后代的适合度评价。可见,目前不仅我国政府发布实施的转基因大豆环境安全评价技术标准中缺少转基因大豆向野生大豆发生基因漂移所产生杂交后代适合度评价的要求,而且相关的科学研究在广度和深度上也存在很多不足。
5 转基因大豆向野生大豆基因漂移研究的未来发展趋势
为了解决我国对于大豆的巨大刚性需求,除了继续大规模进口转基因大豆以外,我国大豆产业界对自主研发转基因大豆商业化种植的呼声也越来越高。作物野生种的保护已经引起国际组织和各国政府的高度重视,而野生大豆是我国和世界宝贵的生物遗传资源[75-76]。转基因大豆大规模商业化种植会产生其向野生大豆发生基因漂移的风险,为了在保护野生大豆的前提下进行转基因大豆的产业化,未来需要从以下几个方面就转基因大豆向野生大豆基因漂移可能对野生大豆生物物种资源保护所产生的风险问题进行深入而系统的评估和研究。
(1)转基因大豆与我国不同区域野生大豆发生基因漂移的风险程度。我国既是栽培大豆重要产区,又有野生大豆大规模分布的区域,调查该区域栽培大豆与野生大豆的地理分布、花期重叠程度、传粉昆虫的种类和数量等,研究转基因大豆与这些区域野生大豆之间的地理重叠情况、杂交亲和性、基因漂移的距离和频率,明确转基因大豆与不同地区野生大豆间基因漂移的风险程度,为确定转基因大豆产业化可能的重点区域以及开展安全管理提供科学数据。
(2)“转基因大豆-野生大豆”杂交回交后代适合度。首先,要研究除了表达EPSPS基因以外的表达其他外源基因的大豆以及同时表达多个外源基因的复合性状转基因大豆与野生大豆杂交后代适合度;其次,外源基因如果通过基因漂移从转基因大豆进入野生大豆,不仅可能通过杂交后代的自交而传递下去,还可能通过杂交后代与野生大豆的回交而在野生大豆群体内和群体间扩散,因此,还需要研究和评价转基因大豆与野生大豆杂交后代与野生大豆回交所产生的回交后代的适合度;第三,应考虑野生大豆生长环境的多样性,尽可能地在模拟的野生大豆各种典型自然生长环境条件下研究和评价“转基因大豆-野生大豆”杂交后代适合度;第四,测定的指标不仅包括营养生长和生殖生长指标,还应包括外源基因表达、休眠性和落粒性等指标。
(3)EPSPS基因对野生大豆适合度的影响。从世界范围来看,已经投入商业化应用的耐除草剂基因种类很多,其中EPSPS基因是应用范围最大的耐除草剂基因[77]。我国进口的绝大部分转基因大豆品种中含有EPSPS基因,目前具备产业化价值的自主研发转基因大豆所含有的外源基因也以EPSPS基因为主。在施用靶标除草剂草甘膦的条件下,表达EPSPS基因的“转基因作物-野生近缘种”杂交回交后代个体比不表达EPSPS基因的个体有极其显著的选择优势。最新的一些研究结果表明,在没有靶标除草剂的选择压力下,EPSPS基因不会对野生近缘种带来适合度代价(costs),而是带来适合度利益(benefits)[78-81]。GUAN等[71]和阚贵珍等[72]的研究结果也表明,在没有草甘膦除草剂的情况下,表达EPSPS基因的“转基因大豆-野生大豆”杂交后代在某些指标上表现出适合度利益。这些研究结果显示,作物的野生近缘种如果通过基因漂移被转入和表达EPSPS基因,将可能在没有草甘膦的自然生态系统显著增加适合度,从而产生难以预料的风险。产生这种现象的可能原因是:通过莽草酸途径所产生的芳香族氨基酸对于维持植物本身的新陈代谢与生长繁殖有着重要作用,而EPSPS蛋白是几乎所有植物和微生物莽草酸途径中所依赖的关键酶,不同来源的EPSPS蛋白虽然结构不同,但是生物学功能相似[82-83];即使在没有草甘膦存在的情况下,外源EPSPS基因表达的EPSPS蛋白依然可以对植物的生长发育产生正面价值。如果表达EPSPS基因的“转基因大豆-野生大豆”杂交后代的适合度高于野生大豆,将产生一些不可预期的风险:一方面可能产生适合度和竞争能力更强的含转基因杂交后代群体,通过遗传同化作用取代局部分布的野生大豆小群体,改变野生大豆群体的遗传多样性和遗传结构;另一方面,由于EPSPS转基因的强自然选择优势,扩散到野生大豆群体后也可能产生选择性剔除效应,使转基因及其位点周边区域位点的基因多态性降低,导致遗传多样性下降。因此,与转Bt基因作物在没有靶标害虫的情况下表现出适合度代价的结果不同[8],通过过量表达EPSPS蛋白而达到草甘膦抗性的转EPSPS基因大豆向野生大豆基因漂移所产生的风险需要重点加以评估和研究。
(4)进口转基因大豆向野生大豆的基因漂移风险。近10 a来,我国一直是世界上最大的转基因大豆进口国,其中,2017年进口量达9 553万t。巨量的进口转基因大豆在国内的转运、储存和加工过程中可能会有少部分种子意外散失到农田或者自然生态系统,引发向栽培大豆和野生大豆的基因漂移。在日本和韩国就曾经出现进口的转基因大豆、玉米洒落在运输线路和储存地点附近并生长的情况[84-85]。鉴于我国已经而且将继续大量进口转基因大豆,应开展进口转基因大豆在国内的运输线路、储存和加工地点及其周围环境中残留状况的调查,为评估和管理进口转基因大豆可能产生的基因漂移风险提供更多的数据支持。
(5)转基因大豆向野生大豆基因漂移的安全控制和管理措施。根据转基因玉米、油菜、棉花在加拿大、阿根廷和墨西哥大规模种植或者进口后已经向其野生近缘种发生基因漂移的案例,结合栽培大豆与野生大豆之间基因漂移的已有研究结果,再考虑到野生大豆在我国的地理分布极其广泛和以小农户为主的栽培大豆分散种植模式,如果转基因大豆在我国进行大规模、长期的商业化种植,其向野生大豆发生基因漂移几乎是不可避免的。因此,在充分调查和研究的基础上,制定适合我国国情的控制转基因大豆向野生大豆基因漂移可能产生风险的安全管理措施是极其必要的。
猜你喜欢
学与玩(2022年10期)2022-11-23
舰船科学技术(2022年11期)2022-07-15
今日农业(2022年3期)2022-06-05
今日农业(2021年19期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
今日农业(2021年10期)2021-11-27
疯狂英语·初中天地(2021年6期)2021-08-06
创新科技(2015年1期)2015-12-24
小说月刊(2014年11期)2014-04-18
食品工业科技(2014年21期)2014-03-11
