
水稻粒长基因的研究进展
2020-09-06 14:05郑跃滨杨琬祺赵海燕王兰
安徽农业科学 2020年15期
郑跃滨 杨琬祺 赵海燕 王兰

摘要粒长是水稻重要的农艺性状之一,既影响水稻产量,又影响稻米品质。水稻的粒长是数量性状,遗传机理复杂。对控制水稻粒长基因的QTL定位、粒长基因的克隆与功能分析、粒长基因的相互作用及粒长基因在分子育种上的应用等方面进行综述,旨在为水稻的粒形育种与品质改良提供参考。
关键词水稻;粒长基因;QTL;克隆
中图分类号S511文献标识码A
文章编号0517-6611(2020)15-0004-05
doi:10.3969/j.issn.0517-6611.2020.15.002
开放科学(资源服务)标识码(OSID):
Research Advances of Genes on Controlling Rice Grain Length
ZHENG Yuebin1,YANG Wanqi2,ZHAO Haiyan1 et al
(1.College of Agriculture,South China Agricultural University,Key Laboratory of Molecular Breeding in Guangdong Province,Guangzhou,Guangdong 510642;2.College of Physics and Electronic Engineering,Harbin Normal University,Harbin,Heilongjiang 150042)
AbstractGrain length is one of the important agronomic traits of rice,which affects both rice yield and rice quality.Grain length of rice is a quantitative trait and its genetic mechanism is very complex.The paper summarized the research results of QTLs mapping,gene cloning and functional analysis,and genes interactions of rice grain length genes,the application of grain length genes in molecular breeding,in order to provide reference for grain shape breeding and quality improvement of rice.
Key wordsRice;Grain length genes;QTL;Cloning
基金項目广东省教育厅科技创新项目(2013KJCX0035)。
作者简介郑跃滨(1995—),男,河北抚宁人,硕士研究生,研究方向:水稻遗传育种。*通信作者,副教授,博士,从事水稻分子遗传研究。
收稿日期2020-02-27;修回日期2020-04-20
粒长(grain length,GL)是水稻粒形的三大构成因素(粒长、粒宽、粒厚)之一。粒长、粒宽和粒厚对粒重的总影响力达94.4%,直接影响水稻的单产[1],其中粒长对水稻粒重的贡献更大[2]。研究表明,粒形细长的品种米质较好[3-4]。进行品质改良时,选育的稻米要求粒形较长、宽度较小、长宽比适中。因此,粒长也是稻米品质的一个重要指标。
从20世纪30年代开始,日本就开始对粳稻品种的矮化进行研究[5],水稻矮秆基因及杂种优势的利用,实现了水稻产量的两次跨越[6]。但随着耕地面积的减少,人民生活水平的提高,现有的水稻单产与品质已不能满足人民生活的需求。改变水稻籽粒的形状,不仅能增加水稻的产量,还能提高稻米的品质。近年来,随着分子生物技术及测序技术的发展,通过改变水稻粒形来育种已越来越受到水稻育种家的重视。通过BSA重测序定位及KASP精细定位来定位水稻粒长基因,进而克隆该粒长基因,再通过基因编辑技术来改良籽粒形状,提高水稻产量,成为最新克隆基因的热点。研究表明,水稻谷粒粒形受多基因控制,属于数量遗传性状,遗传基础很复杂[7],并且粒长基因以加性效应为主,同时还存在显性作用[8]。笔者对控制水稻粒长基因的QTL定位进行总结,并对已克隆粒长基因进行功能分析,以及已克隆粒长基因的相互作用和在当代分子育种上的应用进行深入探讨,以期为水稻粒形的分子育种提供帮助。
1控制粒长相关基因的QTL定位
随着水稻功能基因组学和重测序技术的快速发展,对粒长基因的定位越来越多。据不完全统计,目前已定位到与粒长相关的 QTLs 达120 个,以 3、2、1 和 10 号染色体上最多。赵明富等[9]对BC 3F 2进行QTL定位,结果发现控制粒长QTL(暂定名GL2(t))位于第2染色体,与RM530 连锁,遗传距离为4.1 cM,是个尚未报道的粒长显性主效QTL。高虹等[10]用小粒水稻材料“日本小黑稻”和大粒水稻材料“80018-TR161-2-1”及其F 2和F 2∶3家系检测到谷粒长度受1对隐性核基因控制,命名为GL3。将该基因定位在水稻第3号染色体上SSR标记PSM379和RM16之间,它们的遗传距离分别为4.0和11.2 cM。Shao等[11]利用粳稻品种D50和籼稻品种HB27构建的重组自交系,定位到粒长QTL qGL7-2,在InDel1和RM21945这2个标记之间278 kb的物理范围内。Bai等[12]用籼稻品种南阳占和粳稻品种川7构建的F 7∶8重组自交系群体RIL,发现4个影响粒长的QTL,分别定位于第3、7、10号染色体,其中一个微效QTL,qGL7被定位到一个InDel标记RID711和RM6389之间258 kb的范围内(以日本晴基因组序列为参照),且基因与InDel标记RID710和RID76共分离。曾瑞珍等[13]利用单片段代换系定位发现粒长QTL gl-3被定位于第3号染色体的SSR标记RM6146和PSM377之间,遗传距离分别为1.5和11.0 cM[14]。笔者利用小粒矮秆野生稻品系S18和长粒栽培稻品系KJ01组配定位群体F2,也定位了一个新的主效QTL,位于3号染色体上(结果未发表)。
由于不同亲本构建的不同群体以及环境的影响,被定位的这些QTLs,有些可能是同一位点。由于粒长基因遗传机制复杂,定位的QTLs很多,但迄今为止,已克隆和进行功能研究的QTLs不多。
2粒长相关基因克隆及功能分析
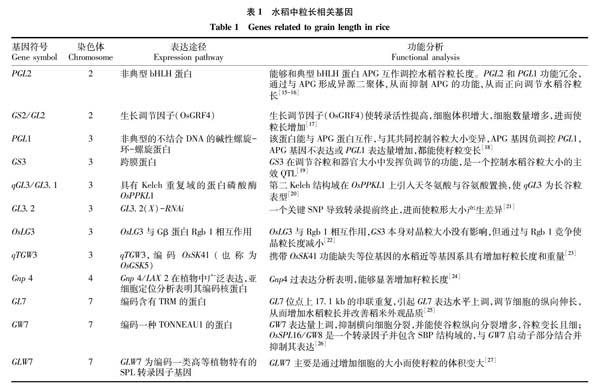
目前,已定位的这些QTLs中,其中有12个粒长基因进行了深入的功能研究,这些基因调控籽粒长度的机制不同(表1)。
尉鑫等[28]通过总结认为在内外颖过量表达PGL1,增加水稻谷粒的长度和重量,是一种结合DNA 的典型bHLH的抑制子。在内外颖过量表达PGL2 也同样可以使水稻谷粒的长度和重量增加[15],对粒长的调控是与基因的表达水平相关。
GS2在显性QTL中,被定位到2号染色体上7.4 kb区间内部,编码生长调节因子(OsGRF4)。OsGRF4为一种转录调节因子,可能作为转录激活剂,增强转录活性。GS2被定位于细胞核表达,同时GS2中的一个突变位点使microRNA、OsmiR396c发生改变,导致表达量升高,GS2表达使细胞增大增多,从而使籽粒变长[17]。
GS3编码一个跨膜蛋白[29],在长粒品种中第二个外显子处有一个碱基C突变为A,产生终止突变。为了证实GS3功能区是否在氨基端,即在OSR区,利用结构域缺失的方法,在水稻明恢63中,转入不同的缺失结构域,结果发现转化后植株粒形都变短,而没转化植株粒形无变化;同样,验证羧基端的功能,发现OSR的功能也会受到2个缺失结构域的抑制影响,转化到明恢63后粒形同样变短[30]。证实GS3对粒长负调控的功能,并且证明OSR是控制粒形的关键结构域,阐述了GS3基因不同结构域的功能与粒形自然变异之间的关联[19]。
qGL3在3号染色体上被用简单重复序列(SSR)标记定位到RM15551和RM15578之间,且95.57%的表型变异在N411×93-11 BC 2F 2群体之中,经过BC 2F 2群体的4个杂合单株筛选得到2 968个单株,在BC 2F 3的2 968个体之中,将区段定位缩小到插入缺失标记(InDel)XJ39与XJ26之间的46.6 kb之间。在该区域内有5个预测ORFs,并通过RT-PCR,发现仅表达ORF3和ORF4,而ORF1和ORF2编码反转录转座子蛋白,编码转座子的ORF5在水稻中不表达。并发现ORF4中存在的4个SNPs,其中前2个SNPs发生在第10个和第11个外显子上,从而导致从天冬氨酸变为谷氨酸和组氨酸变为酪氨酸。ORF4很可能是qGL3的候选基因,该ORF4编码的PP2A,因此被命名为OsPPKL1,属于一个小基因家族成员[20]。
GL3.2是利用大粒品种“南洋占”和小粒品种“川7”为亲本,并构建重组自交系群体,定位到一个主效QTL,发现这个主效QTL可以同时影响粒长与粒重。通过群体构建,相关的遗传分析及图位克隆,确定了候选基因GL3.2。由于GL3.2(X)-RNAi的存在,可以使候选基因GL3.2的表达得以抑制,将GL3.2(X)-RNAi转入到小粒材料中,发现转基因后的材料(6.3 mm)粒长比小粒材料(6.0 mm)长0.3 mm,并通过测序发现,致使2种粒型差异是由一个关键的SNP,转录提前终止所导致[21]。
Yu等[22]从504份栽培稻品种(Oryza ativa L.)中筛选出了99个粒长QTL,其中13个经4个连锁引物验证,92个为新的粒长位点。克隆了一个新基因OsLG3,位于3号染色体上,它可以作为水稻籽粒長度的正调节因子,在不影响稻米品质的前提下提高水稻产量。并对283份地理范围的水稻材料启动子区进行序列分析,发现4个单倍型似乎与籽粒长度有关。进一步分析表明,籼稻和粳稻中OsLG3等位基因的进化独立于不同的祖先和低的核苷酸多样性。系统发育分析表明,OsLG3在提高粳稻育种籽粒长度方面具有很大的潜在价值。OsLG3是一种很有前途的基因,可用于水稻品种改良的基因组DEP 1和GGC 2分别或与Gβ蛋白Rgb 1相互作用,使籽粒长度增加。
qTGW3控制水稻的粒长和重量。qTGW3通过编码OsSK41(也称为OsGSK5),GLYCOGEN SYNTHASE KINASE的成员,增加水稻籽粒长度和重量。OsSK41与AUXIN相互作用,使其磷酸化,从而形成OsARF4。OsSK41与OsARF4的共表达使OsARF4的积累增加,OsARF4存在水稻原生质体中,由于OsARF4的功能丧失导致籽粒增大。 分析表明,OsARF4和OsSK41还共同抑制了一组下游控制水稻籽粒发育过程的基因,包括一些生长素的响应基因,也会造成粒形的改变。因此,研究揭示了OsSK41在水稻籽粒中的重要作用,开发并提供新的候选基因,用于遗传改良水稻的产量并也可能应用在其他谷类作物中[23]。
Zhang等[24]将目的基因Gnp4定位在4号染色体上一个10.7 kb的区间内,并筛选到3个与其互作蛋白LAX1、OsIAA3和OsIAA17,发现Gnp4与OsIAA3或OsIAA17互作可以调控水稻籽粒长度;通过研究发现在过表达Gnp4植株中,提高自由生长素含量,可以激活ARF家族转录因子下游基因的调控,从而调控水稻籽粒长度过量表达,显著增加粒长。
GL7被Wang等[25]定位到CAPS1和210Q标记之间,跨度为20.4 kb,位于7号染色体上,区段内的基因座包含2个候选基因Os07g0603300(LOC_Os07g41200)和Os07g0603400(LOC_Os07g41210),Os07g0603400未具有编码蛋白质的功能表达。Os07g0603300编码含有TON1 RECRUIT MOTIF(TRM)的蛋白质,该蛋白质与来自拟南芥的LONGIFOLIA1和LONGIFOLIA2蛋白质具有20%~22%的氨基酸序列同一性,其可以调节纵向细胞伸长[31]。LONGIFOLIA2(也称为TRM1)能够结合微管,并可能通过将TON1富集到皮质微管阵来参与引导细胞的扩增[32]。
GW7被Wang等[26]从TFA和HJX74(以HJX74作为轮回亲本)的回交培育的4 500个BC 3F 2群体对qGW7进行精细定位,使得基因座的位置被限制在侧翼为标记M 1和M 10的20 kb区段。对纯合分离的子代测试,进一步将间隔缩小到侧翼标记S 5和S 6的2.6 kb区域;这段DNA包含LOC_Os07g41200基因的启动子区和第一个外显子部分。在受精过程之前,小穗颖壳NIL-gw7HJX74比NIL-GW7TFA形成的细胞更短更宽。切片显示NIL-GW7TFA中的内部薄壁细胞少于NIL-gw7HJX74。 平均宽度NIL-GW7TFA外表皮细胞略高于NIL-gw7HJX74细胞,但NIL-GW7TFA小穗壳中横向细胞增殖减少约6.4%,在NIL-GW7TFA横向细胞分裂减少导致粒宽变短,而NIL-GW7TFA小穗颖壳中纵向细胞增殖增加6.5%。 这些发现是指长粒的形成是由于纵向细胞分裂增加和横向细胞分裂减少所致。因此,GW7是通过改变细胞分裂模式来调节水稻的粒形。
GLW7在7号染色体上,编码植物特异性转录因子OsSPL13的主要数量性状基因座,GLW7正向调节籽粒的细胞大小,从而增加水稻籽粒长度和产量。并确定了串联重复序列OsSPL13的5'UTR通过影响转录和翻译来改变其表达,且OsSPL13的表达量较高,与热带粳稻中的大粒相关。进一步分析表明,热带粳稻GLW7的大粒等位基因在人工选择下,可以从籼稻品种转到粳稻品种。研究表明,新基因可以依靠全基因组关联数据有效地确定[27]。
通过以上总结,已克隆的粒长基因调控机制复杂,不同的粒长基因调控籽粒长度的机理不同,有些正调控籽粒长度,如GLW7;有些负调控籽粒长度,如GS3。有些是由于结构域缺失或移码突变,导致基因功能丧失,从而改变籽粒长度,如GS3、qTGW3;有些是由于SNP改变,引起氨基酸改变,从而改变籽粒长度,如qGL3;有些是转录提前终止,产生长短不同的2条蛋白,籽粒也相应表现出长短差异,如GL3.2;也有些是与转录因子相互作用,共同调节籽粒长度,如Gnp4。
3粒长基因间相互作用
控制水稻粒形性状的QTL之间存在相互作用。HGW可以促进水稻粒重和抽穗相关类基因的表达。 在突变体 HGW中, GW2、GW5 、GIF1和GS3 等基因会降低表达量,其中GIF1 基因表达量降低最为明显[33-34]。目前已在水稻中克隆到4个与种子大小相关的基因(GS3、GW2、qSW5/GW5和GIF1)。然而,这4个基因之间的关系尚不清楚,这将在一定程度上阻碍基因聚合育种的进程。为了揭示上述4个基因之间的关系,在转录水平上对GS3-RNAi、GW2-RNAi和qSW5的CSSL进行了基因表达分析。结果表明,qSW5和GW2对GS3的表达具有明显的正调控作用。同时,qSW5可通过抑制GW2转录而下调。此外,qSW5对GIF1的表达有正调控作用,而GW2和GS3对GIF1的表达有负调节作用。在此基础上,以180个水稻品种为材料,对qSW5和GS3的等位基因效应进行了研究。结果表明,这2个基因之间存在互作效应,其中影响种子长度的qSW5被GS3等位基因掩盖,影响种子宽度的GS3被qSW5等位基因掩盖。这些发现为深入了解水稻种子大小发育的分子机制提供了更多的信息,对提高水稻产量有一定的参考价值[35]。
Gao等[36]利用93-11(NIL-gs3/qGL3)连续回交多代以及标记辅助筛查方法获得2种近等基因系:NIL-GS3/qGL3和NIL-gs3/qgl3。再將2种近等基因系杂交,并利用分子标记辅助获得后代群体。通过比较各群体的粒长变化,利用两因素方差分析发现GS3和qGL3在粒长发育中存在加性效应。以NIL-GS3(GS3/qGL3)的粒长作为对照,GS3的突变将粒长从8.5 mm (GS3/qGL3)增加到10.2 mm (gs3/qGL3),qGL3的突变将粒长从8.5 mm(GS3/qGL3)增加到11.2 mm(GS3/qgl3),将2个基因同时突变后粒长从8.5 mm(GS3/qGL3)增加到12.2 mm(gs3/qgl3)。通过粒长性状直接可以看出当两基因同时存在时,粒长增加更为明显,比单一基因存在效果更好,对两基因的转录研究发现,存在重叠效应,并推测是qGL3和GS3共同参与BR信号转导,从而调控粒长[37]。
4粒长基因在分子育种的应用
已克隆的一批控制水稻粒长的重要基因,既有助于揭示水稻产量性状复杂的遗传机制,同时也为水稻分子标记的辅助选择提供了理论依据和技术基础。分子标记辅助选择技术是在现代作物改良育种中应用最广泛的技术之一,对现代分子育种有着十分重要的作用。该技术结合分子标记鉴定筛选,通过田间杂交和回交,可以快速有效地达到转移基因和聚合多个基因等育种目的[6]。这不仅降低了育种成本,更缩短了育种时间,育种研究者能更好更快地选育出目标品种。
Huang等[38]在其现有嘉禾系列中发现,都存在gs3、dep1等能够影响粒形的基因。黄海祥等[39]发现在其品种嘉禾218的粒形基因上,GS3产生C-A的单碱基突变,谷粒长度达7.0 mm,长宽比达3.0,以国内的籼稻一级稻米为标准,长度6.5 mm,长宽比2.8,显然嘉禾218更高于此标准。利用已克隆的基因来改良现有品种的粒形,实现分子辅助育种。张剑霞[40]通过分子标记辅助选择技术,改良了珍汕97B和Ⅱ-32B,将Xa23和GS3分别聚合到珍汕97B和Ⅱ-32B中,使得珍汕97B由8.00 mm增加到(9.81±0.31)mm,Ⅱ-32B 由8.00 mm增加到(9.71±0.26)mm,改良后效果十分明显。Fan等[41]通过测序比较GS3第二个外显子发生碱基突变,导致转录提前终止,从而在此功能位点设计标记,并用检测分析180份水稻品种,发现所有小粒形都为‘TGC基因型,大粒形都为‘TGA(终止密码)基因型,通过此标记,能更好观测出基因不同,从而导致性状的分型。Yu等[22]发现在现有粳稻品种中导入OsLG3的SLG等位基因可增加粒长、粒重。在不影响籽粒品质的前提下,提高产量。
5展望
早年来,受技术、气候等因素限制,虽然水稻的分布范围广,从热带到寒带都有种植。但不断恶化的地球环境给世界上粮食作物的生产带来了诸多不良影响,使得水稻的种植面积日益缩减。因此,水稻产量对国家粮食的安全和社会稳定的发展至关重要[42]。在有限的耕种面积里种植出稳产高产的水稻是育种工作者的重任,进一步加大水稻粒长种质创新、遗传规律探索及分子标记开发、粒长相关基因克隆及转基因等基础方面的研究显得尤为重要,对于加快水稻粒长新品种选育,培育综合性状优良、适应性广的新品种,对解决粮食问题具有重要意义。
近年来,伴随杂交水稻的广泛种植,随着人们步入小康社会,能吃饱已不能满足人们的需求。粒形较长的长粒稻在外观、口感等方面也更受人们的喜爱。虽然水稻粒形性状的遗传分析研究也已经取得了很大进展,对稻米主要品质性状的基因认识也较为明晰,并对稻米品质性状的形成也有了相应动态研究[43],但对于粒长基因克隆、调控机理、相关基因功能研究相对较少,在大面积田间推广以达到增加水稻产量的效果更是少之又少。
目前,粒形特长的长粒栽培稻品种经过不断选择,促使细胞内发生基因的重新组合优化遗传性状,来获得所需要的高产、优质品种。并在提高粒长的同时,兼顾稻米品质、营养成分,能够培育出符合人们需要的粒形。因此,从栽培稻中挖掘粒长基因,从而改变籽粒长度,提高水稻产量,是提高粮食产量和质量的一个重要途径。
参考文献
[1]XIE X B,SONG M H,JIN F X,et al.Fine mapping of a grain weight quantitative trait locus on rice chromosome 8 using nearisogenic lines derived from a cross between Oryza sativa and Oryza rufipogon[J].Theoretical and applied genetics,2006,113(5):885-894.
[2] 林荔辉,吴为人.水稻粒型和粒重的QTL定位分析[J].分子植物育种,2003,1(3):337-342.
[3] 石春海,朱军.水稻植株农艺性状与稻米碾磨品质的遗传相关性分析[J].浙江农业大学学报,1997,23(3):105-111.
[4] 陈能,罗玉坤,朱智伟,等.优质食用稻米品质的理化指标与食味的相关性研究[J].中国水稻科学,1997,11(2):70-76.
[5] 顾铭洪.矮源及其在水稻育种上的利用[J].江苏农学院学报,1980,1(1):40-44.
[6] 刘喜,牟昌铃,周春雷,等.水稻粒型基因克隆和调控机制研究进展[J].中国水稻科学,2018,32(1):1-11.
[7] 丁军.水稻外观品质在杂种F 9∶10的表现及其QTL定位[D].贵阳:贵州大学,2016.
[8] 闵超,陶慧敏,朱克明.水稻种子相关性状的研究进展[J].种子,2016,35(4):51-56.
[9] 赵明富,黄招德,吴春珠,等.水稻谷粒粒长显性主效QTL的遗传分析与定位[J].分子植物育种,2008,6(6):1057-1060.
[10] 高虹,王嘉宇,姜树坤,等.水稻粒长基因GL3的遗传分析和分子标记定位[J].植物生理学通讯,2010,46(3):236-240.
[11] SHAO G N,TANG S Q,LUO J,et al.Mapping of qGL72,a grain length QTL on chromosome 7 of rice[J].Journal of genetics and genomics,2010,37(8):523-531.
[12] BAI X F,LUO L J,YAN W H,et al.Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7[J].BMC Genetics,2010,11(1):1-11.
[13] 曾瑞珍,TALUKDAR A,劉芳,等.利用单片段代换系定位水稻粒形QTL[J].中国农业科学,2006,39(4):647-654.
[14] 齐兰.基于野生稻染色体置换系的粒型QTL分析和qGL12.2的精细定位[D].海口:海南大学,2017.
[15] HEANG D,SASSA H.Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice[J].PLoS One,2012,7(2):1-11.
[16] 刘成兵,唐明凤,张德春.水稻粒形控制基因研究进展[J].广东农业科学,2014,41(8):30-34.
[17] HU J,WANG Y X,FANG Y X,et al.A rare allele of GS2 enhances grain size and grain yield in rice[J].Molecular plant,2015,8(10):1455-1465.
[18] HEANG D,SASSA H.An atypical bHLH protein encoded by POSITIVE REGULATOR OF GRAIN LENGTH 2 is involved in controlling grain length and weight of rice through interaction with a typical bHLH protein APG[J].Breeding science,2012,62(2):133-141.
[19] MAO H L,SUN S Y,YAO J L,et al.Linking differential domain functions of the GS3 protein to natural variation of grain size in rice[J].Proceedings of the national academy of sciences,2010,107(45):19579-19584.
[20] ZHANG X J,WANG J F,HUANG J,et al.Rare allele of OsPPKL1 associated with grain length causes extralarge grain and a significant yield increase in rice[J].Proceedings of the national academy of sciences,2012,109(52):1-6.
[21] 李秋苹.OsHAP家族基因的功能研究和粒形基因GL3.2的图位克隆[D].武汉:华中农业大学,2016.
[22] YU J P,XIONG H Y,ZHU X Y,et al.OsLG3 contributing to rice grain length and yield was mined by HoLAMap[J].BMC Biology,2017,15(1):18-28.
[23] HU Z J,LU S J,WANG M J,et al.A novel QTL qTGW3 encodes the GSK3/SHAGGYlike linase OsGSK5/OsSK41 that interacts with OsARF4 to negatively regulate grain size and weight in rice[J].Molecular plant,2018,11(5):736-749.
[24] ZHANG Z Y,LI J J,TANG Z S,et al.Gnp4/LAX2,a RAWUL protein,interferes with the OsIAA3OsARF25 interaction to regulate grain length via the auxin signaling pathway in rice[J].Journal of experimental botany,2018,69(20):4723-4737.
[25] WANG Y X,XIONG G S,HU J,et al.Copy number variation at the GL7 locus contributes to grain size diversity in rice[J].Nature genetics,2015,47(8):944-948.
[26] WANG S K,LI S,LIU Q,et al.The OsSPL16GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J].Nature genetics,2015,47(8):949-954.
[27] SI L Z,CHEN J Y,HUANG X H,et al.OsSPL13 controls grain size in cultivated rice[J].Nature genetics,2016,48(4):447-456.
[28] 尉鑫,曾智鋒,杨维丰,等.水稻粒形遗传调控研究进展[J].安徽农业科学,2019,47(5):21-28.
[29] FAN C C,XING Y Z,MAO H L,et al.GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J].Theoretical and applied genetics,2006,112(6):1164-1171.
[30] 茆海亮.水稻粒形基因GS3的功能研究[D].武汉:华中农业大学,2010.
[31] LEE Y K,KIM G T,KIM I J,et al.LONGIFOLIA1 and LONGIFOLIA2,two homologous genes,regulate longitudinal cell elongation in Arabidopsis[J].Development,2006,133(21):4305-4314.
[32] DREVENSEK S,GOUSSOT M,DUROC Y,et al.The Arabidopsis TRM1-TON1 interaction reveals a recruitment network common to plant cortical microtubule arrays and eukaryotic centrosomes[J].Plant cell,2012,24(1):178-191.
[33] LI J,CHU H W,ZHANG Y H,et al.The rice HGW gene encodes a ubiquitinassociated (UBA) domain protein that regulates heading date and grain weight[J].PLoS One,2012,7(3):1-12.
[34] 陈立凯.水稻籽粒相关性状QTL鉴定与基因克隆研究[D].广州:华南农业大学,2016.
[35] YAN S,ZOU G H,LI S J,et al.Seed size is determined by the combinations of the genes controlling different seed characteristics in rice[J].Theoretical and applied genetics,2011,123(7):1173-1181.
[36] GAO X Y,ZHANG X J,LAN H X,et al.The additive effects of GS3 and qGL3 on rice grain length regulation revealed by genetic and transcriptome comparisons[J].BMC Plant Biology,2015,15:1-13.
[37] 高秀莹.OsPPKL1调控水稻粒長的分子机制研究[D].南京:南京农业大学,2016.
[38] HUANG X Z,QIAN Q,LIU Z B,et al.Natural variation at the DEP1 locus enhances grain yield in rice[J].Nature genetics,2009,41(4):494-497.
[39] 黄海祥,钱前.水稻粒形遗传与长粒型优质粳稻育种进展[J].中国水稻科学,2017,31(6):665-672.
[40] 张剑霞.利用分子标记辅助选择转移野生稻增产QTL和聚合水稻优良基因[D].武汉:华中农业大学,2009.
[41] FAN C C,YU S B,WANG C R,et al.A causal CA mutation in the second exon of GS3 highly associated with rice grain length and validated as a functional marker[J].Theoretical and applied genetics,2009,118(3):465-472.
[42] 阮班普.水稻粒型的遗传分析及qGW2的精细定位[D].金华:浙江师范大学,2014.
[43] 王丰,程方民.从籽粒灌浆过程上讨论水稻粒间品质差异形成的生理机制[J].种子,2004(1):31-35.
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
今日农业(2020年24期)2020-12-15
小学科学(学生版)(2019年5期)2019-05-21
兽医导刊(2016年12期)2016-05-17
现代检验医学杂志(2015年4期)2015-02-06
