
不同物候期妃子笑荔枝叶片光合特性及碳水化合物含量变化分析
2020-10-09 09:10周瑞云胡福初王祥和周文静吴凤芝
广东农业科学 2020年8期
周瑞云,胡福初,王祥和,周文静,吴凤芝,陈 哲
(海南省农业科学院热带果树研究所/海南省热带果树生物学重点实验室/农业农村部海口热带果树科学观测实验站,海南 海口 571100)
【研究意义】妃子笑荔枝是我国古老而著名的早熟优良荔枝品种,原产自广东,其果大肉厚、种子多不饱满、果肉肥厚多汁、风味浓甜,在我国荔枝主产区均有分布,海南目前妃子笑种植面积达90%以上,是当地农民创收增收的重要来源,也是海南重要的经济产业支柱[1-3]。妃子笑荔枝树势壮旺、枝条粗壮,树冠略疏散,树皮灰褐色,叶片较宽大,对环境有着极强的适应性,但同时存在花粉发育不良、花而不实等缺点。研究妃子笑荔枝不同物候期叶片光合特性、可溶性糖含量和淀粉含量的动态变化,可为其优质高效栽培提供理论指导。【前人研究进展】光合作用是植物生长发育的基础,影响植物的产量和品质[4]。叶片是植物进行光合作用的主要场所,也是碳氮积累的主要器官,而光合产物碳水化合物参与植物的开花结果等重要生理过程[5]。花芽分化是荔枝生长发育的重要过程,也是复杂形态形成过程。前人关于荔枝花芽分化期间光合特性以及物质含量的动态变化已有较多报道[6-13]。例如,吴志祥等[6-7]对妃子笑和鹅蛋荔在花芽分化期间枝条和花穗的激素变化以及碳氮化合物动态变化进行了研究,发现高GA3、ABA含量和高C/N有利于花芽分化;袁沛元等[8-9]研究了花芽形态分化期至开花期妃子笑叶片氮磷钾、碳水化合物和蛋白质含量的变化,发现花芽分化期间氮含量变化不大,磷、钾含量呈下降趋势;张红娜等[10-11]比较了妃子笑、糯米糍和桂味3个荔枝品种盛花期的光合特性,发现白点期是碳氮物质变化的关键点;陈炫等[12]通过喷施PP333和乙烯利研究其对妃子笑成花及碳氮含量的影响,结果表明喷施PP333和乙烯利可促进花芽分化,且碳氮含量较对照要高;张红娜等[13]研究GA3和PP333对荔枝成花及碳氮营养的影响,发现两者对淀粉含量影响不显著,喷施GA3可推迟成花且降低叶片可溶性糖含量,喷施PP333可促进提早开花并增加叶片可溶性糖含量。【本研究切入点】不同物候期荔枝叶片光合特性及碳水化合物含量都有明显差异,尤其是春季荔枝大量开花,花穗需要大量碳水化合物的积累,此时也是荔枝光合效率较高、对光合产物需求较大的季节,且荔枝属于低光效作物,改善荔枝的光合效能颇为重要。以往大部分研究均是针对荔枝1~2个物候期相关指标进行动态分析,而从秋梢老熟期到果实成熟期的光合特性和生理特征尚缺乏系统研究。【拟解决的关键问题】本研究旨在研究妃子笑荔枝不同物候期叶片光合特性、叶绿素含量和碳水化合物含量,了解不同物候期荔枝叶片光合作用能力、叶绿素含量和碳水化合物的动态变化曲线,以期为妃子笑荔枝的优质高效栽培提供理论指导。
1 材料与方法
1.1 试验地概况
试验区位于海南省澄迈县永发果树基地荔枝示范果园,地理坐标为109°45'~110°15' E、19°23'~20°01' N,属热带季风气候,雨量充沛,日照充足,年平均气温为23.7℃,极端高温38℃,极端低温为7℃。6月最热、平均气温35℃,2月最冷、平均气温15℃。全年日照时数2 060.6 h,年均降雨量1 756 mm,全年无霜期。试验地土壤为红壤土,有机质含量3.6%,pH 5.2。
1.2 试验材料
试验材料为9~10年生妃子笑荔枝品种,常规管理,随机选取3株长势一致的树木,选取无病虫害且无相互遮挡的叶片。
1.3 试验方法
1.3.1 物候期观测 2016—2019年对试验基地内正常发育的妃子笑荔枝植株进行物候期观测,在春秋季物候现象变化较快时每天观测1次,植株开花后每天观测两次。
1.3.2 光合指标测定 2018年11月至2019年5月,选择晴天,采用CIRAS-3便携式植物光合测定仪测定妃子笑荔枝树的光合生理生态特征参数。测定时间分别为秋梢老熟期(2018年11月15日、12月2日、12月21日)、白点期(2019年1月14日)、花期(2019年2月22日)、落果期(2019年3月21日)、果实发育期(2019年4月12日)、果实着色期(2019年4月24日)、果实成熟期(2019年5月13日)、采后修剪期(2019年5月27日)。选定3株长势一致的健康妃子笑荔枝植株,挑取结果母枝末次枝梢中上部且生长相对一致的成熟叶片,每株测定5张叶片,每张叶片测定1个净光合速率瞬时值。测定指标包括大气温度(Ta)、净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)。水分利用效率(WUE)计算公式为:WUE=Pn/Tr。
1.3.3 叶绿素含量测定 选定3株长势一致的健康妃子笑荔枝植株,每株取东、南、西、北、中5个方位的结果母枝末次枝梢中上部的健康叶片各1片。采样时间为秋梢老熟期(2018年11月27日)、白点期(2019年1月7日)、抽蕾期(2019年1月20日)、花期(2019年2月21日)、落果期(2019年3月25日)、果实着色期(2019年4月28日)、果实成熟期(2019年5月13日)。叶绿素的提取采用直接浸提法,称量后剪碎,添加20 mL 95%乙醇浸提,待叶片浸泡至全部发白,用95%乙醇定容至25 mL,过滤后分别于665、649 nm下测定吸光度,计算叶绿素含量(mg/g):
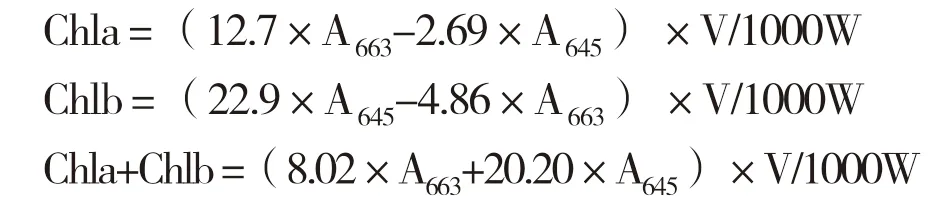
式中,V为样品提取液总体积,W为样品重。
1.3.4 可溶性糖及淀粉含量测定 选定3株长势一致的健康妃子笑荔枝植株,每株取东、南、西、北、中5个方位的结果母枝末次枝梢中上部的健康叶片各3片。采样时间为秋梢老熟期(2018年11月14日、11月27日、12月18日)、白点期(2019年1月7日)、抽蕾期(2019年1月20日)、花期(2019年2月21日)、第1次生理落果期(2019年3月15日)、第2次生理落果期(2019年3月25日)、第3次生理落果期(2019年4月8日)、果实着色期(2019年4月28日)、果实成熟期(2019年5月13日)、采后修剪期(2019年5月27日)。
可溶性糖含量采用蒽酮-硫酸法测定。取0.2 g叶片鲜样,加蒸馏水15 mL于沸水中煮沸30 min,取出冷却,定容至25 mL。取待测样品提取液0.5 mL,加入1.5 mL蒸馏水,加入5 mL蒽酮试剂,摇匀,沸水浴1 min,冷却后于630 nm处测定吸光度,计算可溶性糖含量(mg/g)。
淀粉含量采用碘量法测定。选取3片结果母枝末次枝梢中上部的新鲜成熟叶片,称取重量,剪碎,于研钵中碾碎,加入80%硝酸钙溶液,沸水浴5 min,取出冷却,定容至25 mL刻度线。取5 mL待测样品提取液,加2 mL 0.5%碘液,混匀后离心,弃上清。沉淀用含碘硝酸钙溶液冲洗,加入0.1 mol/L NaOH混匀,沸水浴5 min后转入50 mL容量瓶,加0.3 mL 0.5%碘液,蒸馏水冲洗容量瓶,加入1 mL 0.1 mol/L HCl,蒸馏水定容至刻度线并显色,于590 nm处测定吸光度,计算淀粉含量(mg/g)。
试验数据采用Excel 2007软件进行统计分析,采用Origin 2019b(OriginLab)进行图表绘制。
2 结果与分析
2.1 妃子笑荔枝的物候期
我们对2016—2019年供试妃子笑荔枝的物候期进行了观测,结果(表1)发现,树体在5月下旬至6月上旬进行采后修剪,7月上旬至9月下旬即进入秋梢生长期,期间共有3批秋梢老熟,其中第1批秋梢在7月上旬老熟,第2批秋梢在8月上旬老熟,第3批秋梢在9月中下旬老熟。9月下旬进入控梢期,翌年1月上中旬出现花序原基(白点),1月下旬开始抽蕾,2月中下旬至3月上旬则为花期。3月中旬至4月上旬开始进入生理落果期,5月上旬果实成熟。
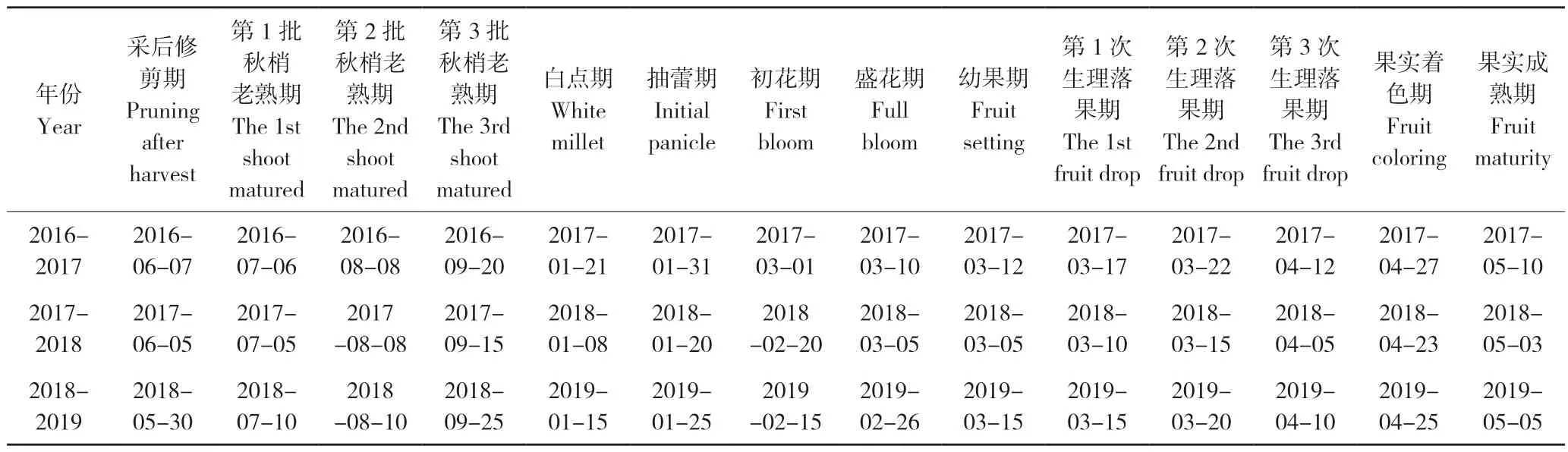
表1 2016-2019年妃子笑荔枝的物候期Table 1 Phenological periods of Feizixiao litchi from 2016 to 2019
2.2 不同物候期妃子笑荔枝叶片光合指标的变化
2.2.1 净光合速率(Pn) 由图1可知,妃子笑荔枝叶片Pn动态变化曲线为不明显的双峰型,Pn高峰值的出现时间受环境影响。叶片Pn在花芽分化期呈现下降趋势,Pn动态曲线的第一个峰值(最高值,13.04±2.38 μmol/m2·s)在此阶段出现,Pn在抽穗期小幅上升而后下降,果实发育期上升,果实着色期下降至最低值(4.73±0.60 μmol/m2·s),果实成熟期Pn再呈上升趋势。
2.2.2 胞间CO2浓度(Ci)和气孔导度(Gs) Pn最高峰值时,叶片Ci、Gs均处于峰值,后期数值下降与温度有一定关系。在果实着色期由于气温较高,叶片为防止水分流失,气孔部分关闭,导致Gs降低,CO2难以进入,促使Ci降低,从而影响净光合速率Pn。
2.2.3 蒸腾速率(Tr) 蒸腾作用是植物进行矿物质运输、叶面温度调节以及为光合作用提供水分的重要过程,与Pn有密切联系[15]。图1显示,Tr动态变化曲线为不明显双峰型,与Pn变化趋势基本保持一致。Tr的第1个峰值(也是最大值,2.84±0.28 mmol/m2·s)出现在花芽分化期,花芽分化期到抽穗期Tr呈现下降趋势,Tr在果实发育期出现第2个峰值,果实着色期下降并处于最低值(1.16±0.19 mmol/m2·s),到果实成熟期再呈现上升趋势。
2.2.4 水分利用效率(WUE) WUE可用于表明植物在进行光合作用时消耗单位水量所能固定CO2的量,一般由Pn/Tr计算而来,是植物适应能力大小的体现,田间条件下,水分不足时能反映植物生长适应程度[16]。由图2可知,WUE的变化曲线与Pn相似,均表现为波浪式减少趋势。本试验中,妃子笑荔枝叶片的WUE共有3个峰值,第1个峰值(最大值,6.48±0.66 μmol/mmol)在抽穗期出现,此时Pn第1个峰值出现,而Tr呈现下降趋势,因此WUE最大;第2个峰值在果实发育期出现,此时Pn和Tr均处于上升状态,但Pn的上升幅度大于Tr,故WUE上升;第3个峰值在果实成熟期出现,此时Pn和Tr均呈现下降趋势,但Pn的下降幅度小于Tr,故WUE呈上升趋势。成熟后期Pn的上升幅度小于Tr,故WUE下降。
2.3 不同物候期妃子笑荔枝叶片叶绿素含量的变化
植物叶片叶绿素含量的变化是体现其生理活性变化的重要指标,了解叶片叶绿素含量的变化规律是提高植物产量的基础[14]。表2反映的是妃子笑荔枝叶片从第3批秋梢老熟期到果实成熟期叶绿素a、叶绿素b以及叶绿素(a+b)含量的动态变化。从表2可以看出,妃子笑荔枝叶片的叶绿素含量在秋梢老熟期最低(1.0±0.10 mg/g),果实着色期叶最高(5.2±0.28 mg/g)。花芽分化期到果实着色期,叶片叶绿素a含量逐渐上升,果实着色期达到最大值;叶绿素b含量与叶绿素a含量的变化趋势基本一致,花芽分化期至果实着色期呈上升趋势,叶绿素b含量的最大值也出现在果实着色期;叶绿素(a+b)含量在花芽分化期间逐渐增加至果实着色期达到最高值。叶绿素a、叶绿素b含量的变化引起了叶绿素(a+b)含量的变化,从秋梢老熟期到果实成熟期叶片叶绿素a、叶绿素b和叶绿素(a+b)含量的变化基本保持一致。
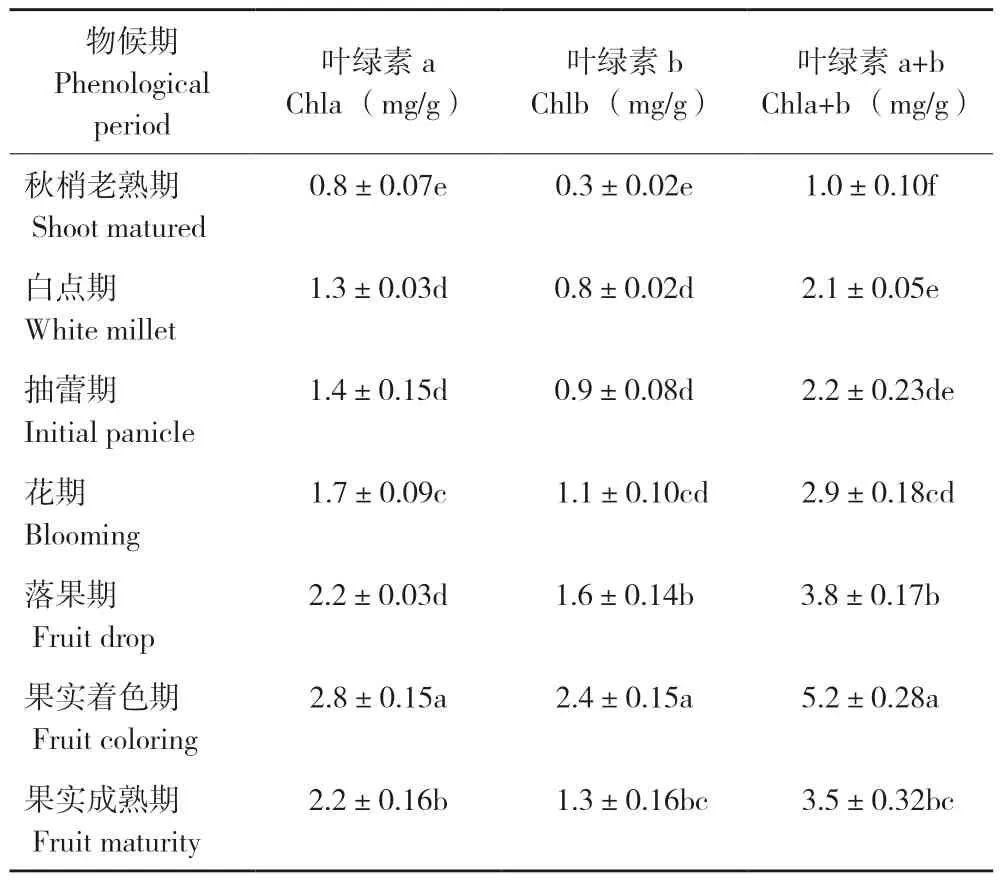
表2 不同物候期妃子笑荔枝叶片叶绿素含量的变化Table 2 Changes of chlorophyll contents of Feizixiao litchi leaves in different phenological periods
2.4 不同物候期妃子笑荔枝叶片碳水化合物含量的变化
2.4.1 可溶性糖含量 可溶性糖是植物光合作用的主要产物,也是碳水化合物代谢和临时储存的主要形式,其在植物代谢中具有重要地位。从图2A可以看出,秋梢老熟期到果实成熟期可溶性糖含量的变化趋势呈波浪形,可溶性糖含量在花芽分化期有两个低值、抽穗期有一个低值、果实发育期有一个低值、果实成熟期出现最低值(16±1.89 mg/g),果实着色期可溶性糖含量呈现最高值(42±2.81 mg/g)。
2.4.2 淀粉含量 如图2B所示,妃子笑荔枝叶片的淀粉含量在花芽分化期呈上升趋势,白点期达最高值(237±8.29 mg/g)。抽穗期淀粉含量开始下降,然后上升后又开始下降,第2次生理落果期淀粉含量最低(102±3.28 mg/g),而后一段时间上升到相对较高值,果实着色期淀粉含量有所下降,果实成熟期小幅下降后又开始上升。
3 讨论
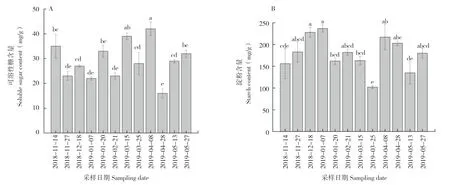
图2 不同物候期妃子笑荔枝叶片碳水化合物含量的变化Fig.2 Changes of carbohydrate content of Feizixiao litchi leaves in different phenological periods
不同物候期妃子笑荔枝叶片的净光合速率(Pn)和碳水化合物含量变化明显[17-18]。本研究发现,从花芽分化期到果实成熟期,妃子笑荔枝叶片净光合速率在花芽分化期呈下降趋势,白点期出现后回升,花蕾期下降,果实发育期有所回升,果实着色期处于最低值,进入果实成熟期后呈上升趋势;叶绿素含量在整个过程中呈现上升趋势,直至果实成熟期有小幅下降。可溶性糖含量在诱导期较高,白点期出现低值,花蕾期回升,进入花芽形态分化阶段开始下降,果实发育期有所回升,落果期达到最高值。在花芽生理分化期,叶片淀粉含量一直在积累,花芽形态分化期下降,果实发育期达到最低值,果实着色期有所回升。
花芽分化期是生长发育的关键时期,也是果树生长发育研究的重要时期。研究表明,无花果花托分化期间叶片可溶性糖含量逐渐增加,而进入花芽分化期可溶性糖含量下降,淀粉含量在整个花芽分化期间变化较为平缓[19];杨桃从“露红”到花芽分化期,枝梢可溶性糖含量和淀粉含量均呈现增加趋势[20];草莓花芽分化过程中叶片可溶性糖含量呈逐步上升趋势,淀粉含量在花芽生理分化期略有下降,花芽分化期上升到最大值后下降[21];李树花芽生理分化期间叶片可溶性糖含量不断增加,并达到最高值,而至花芽形态分化期可溶性糖含量呈明显下降趋势[22]。荔枝结果树生长发育周期分为枝梢生长发育期、花芽分化和花器官生长发育期、果实生长发育期3个阶段,在这3个生长发育时期光合作用能力以及所需碳水化合物的量也不同。从花芽分化期到果实成熟期整个过程中,妃子笑荔枝叶片的净光合速率在果实着色期处于最低值,抽穗期开始上升,花蕾期后出现下降。叶片光合作用受外界环境因素的影响,尤其是温度和光照[23-24]。荔枝成花诱导期需低温环境,导致叶片的净光合速率呈下降趋势;白点期,环境温度回升,叶片的净光合速率也随之回升。而果实着色期气温较高,为阻止叶片内水分流失,叶片气孔部分关闭,导致气孔导度(Gs)下降,进而影响CO2进入叶片细胞,所以胞间CO2浓度(Ci)也下降,故叶片的净光合速率在此时处于最低值。此时若能提高空气湿度,叶片的净光合速率可以有所提升。叶片叶绿素含量整体呈现上升趋势,在果实成熟期后小幅下降,说明妃子笑荔枝从控梢期到果实成熟期这个阶段叶片没有明显褪绿现象;果实着色期叶绿素含量达到最高值,是因为叶片在果实着色期时成熟度最高,所以此时期的叶绿素a、叶绿素b、叶绿素(a+b)呈现出最高峰值。
花芽分化主要涉及到碳水化合物库源方向的转换,在成花诱导期间,花发端要成为调运碳水化合物的库,所以果树花芽分化需要用到较多可溶性糖[22,25]。本研究结果表明,可溶性糖含量在诱导期含量较高,在白点期处于低值,之后花蕾出现,可溶性糖含量上升,这与在龙眼[26]上的研究结果基本一致。在整个花芽分化过程中,初期可溶性糖含量较高,可能是因为淀粉分解成可溶性糖;白点期后出现下降,这与花芽萌发以及抽穗需要消耗大量能量有关;花期前叶片可溶性糖含量上升,为开花积累能量物质;果实发育期,可溶性糖含量变化明显;生理落果期结束后,可溶性糖含量达到最大值;果实着色期又迅速下降到最小值,说明果实发育前期可溶性糖的积累是为后期果实生长发育提供所需能量。淀粉含量高低对植物开花结果有很大影响。花芽分化期初期,淀粉含量与可溶性糖含量变化趋势相反,说明两者之间相互转化;采后修剪后,经过3批枝梢的生长,花芽生理分化期淀粉含量呈现上升趋势,说明淀粉在进行大量积累,为后期叶片花芽分化作足营养储备;抽穗期,淀粉含量下降,是因为淀粉分解转化成可溶性糖,以满足花芽分化期叶片较为旺盛的新陈代谢;果实发育期,淀粉含量处于最低值,是因为果实的生长发育需要消耗大量能量,故淀粉分解转化以提供果实生长所需能量;果实着色期,淀粉含量回升到相对较高水平,再次进行积累,为后续果实的成熟发育提供营养。
4 结论
通过分析妃子笑荔枝不同物候期叶片光合特性和碳水化合物含量的变化情况发现,从花芽分化期到果实成熟期整个过程中,叶片净光合速率存在两个峰值,分别在花芽分化期和果实发育期出现;可溶性糖含量和淀粉含量有两次较大的积累,可溶性糖含量在果实发育期和果实着色期达到最大值,淀粉含量在花芽分化期和果实着色期达到最大值;两次积累均是为后续生长发育储备充足营养。在整个过程中,叶片无明显退绿现象。
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
——妃子笑
宝藏(2021年10期)2021-11-22
——以长白山区为例
东北林业大学学报(2020年9期)2020-10-21
落叶果树(2020年3期)2020-06-17
农业灾害研究(2020年8期)2020-05-11
农业与技术(2019年5期)2019-05-29
农民致富之友(2018年21期)2018-12-27
现代妇女(2018年8期)2018-08-23
