
聚果榕传粉榕小蜂传粉与产卵之间的权衡
2020-10-12 00:56汪雪敏苗白鸽彭艳琼
昆虫学报 2020年7期
汪雪敏,苗白鸽,彭艳琼,*
(1.云南大学生态学与环境学院,昆明 650091;2.中国科学院西双版纳热带植物园,云南勐腊 666303)
有花植物的有性生殖依赖一定的媒介传授花粉(黄双全,2008),除少数为风媒、水媒外,87.5%的被子植物由动物传粉(Ollerton,2017)。全球传粉动物约有35万种,其中昆虫占80%~85%,是种类最多的传粉动物(张立微和张红玉,2015)。昆虫在为植物传授花粉的同时,植物也为昆虫提供食物和生长场所(钦俊德和王琛柱,2001)。绝大多数昆虫都是在取食花粉、花蜜的过程中,体表、口器粘附花粉,被动地为植物完成授粉(黄双全和郭友好,2000);仅有约2%的传粉昆虫具有主动为植物传粉的行为,其为植物主动传粉并且依赖植物繁衍后代,与植物建立高度专性的互惠共生关系,榕树与传粉榕小蜂就是其中经典的案例(Cooketal.,2004;Herreetal.,2008)。
榕属Ficus在植物界具有独特的隐头花序(syconium)结构(通称榕果),只能依赖体长2~3 mm的专性传粉榕小蜂从顶生苞片处钻入瓮型的果腔内传粉,同时榕小蜂把卵产于部分雌花子房中(Janzen,1979;Weiblen,2002)。这种繁殖上的互惠共生关系,至今已协同演化了7 500万年(Cruaudetal.,2012)。已知全世界有约800种榕树、500多种传粉榕小蜂,其中2/3的榕-蜂种对是主动传粉类型,仅1/3的种对为被动传粉类型(Kjellbergetal.,2001;Jousselinetal.,2003;Wangetal.,2014)。主动传粉类型中,榕树及榕小蜂均演化出相互适应的传粉结构与行为,表现为榕果内雄花数量少,花药胚珠比小于0.16,成熟花粉囊不会自动破裂散粉;传粉榕小蜂前足基节上有花粉刷,中胸腹板两侧有一对发达的花粉筐,并且配合有主动传粉行为。而被动传粉的类型,榕果内雄花较多,花药胚珠比大于0.21,成熟花粉囊自动破裂散粉,其传粉榕小蜂的传粉结构和行为消失或退化,依靠体壁粘附花粉进行被动传粉(Kjellbergetal.,2001;Wangetal.,2014)。
传粉榕小蜂成虫寿命通常为1~2 d(Dunnetal.,2008;Zhangetal.,2014),主动传粉的榕小蜂进入雌花期榕果内既要传粉、也要产卵。榕小蜂在有限的存活时间内如何在传粉和产卵之间进行合理分配,是长期被关注但悬而未解的问题。Galil和Eisikowitch(1974)报道,榕小蜂的传粉行为发生在每一次产卵行为末期至小蜂从雌花花柱拔出产卵器之前。也有学者认为,榕小蜂进入雌花期果腔内首先传粉,同时标记适宜产卵的柱头,传粉结束后在标记过的雌花上产卵(杨大荣等,2001)。这些研究都基于显微镜下直接的跟踪观察,研究条件有限。随着技术的发展,现在显微视频技术既能够显微观察、又可以连续记录传粉榕小蜂在果腔内的行为。本研究选择雌雄同株榕树聚果榕,采用显微视频设备,清晰观察、连续记录传粉榕小蜂在雌花期果内搜索、产卵和传粉的行为,并进行相关特征测量及控制性引蜂试验,拟回答如下问题:(1)传粉榕小蜂产卵器长度与雌花花柱长度的匹配程度;(2)雌花期榕果内传粉榕小蜂如何在搜索、产卵及传粉行为之间分配时间;(3)传粉榕小蜂个体大小与孕卵量及携粉量之间的关系;(4)传粉榕小蜂不同状态和工作时间下的产卵和传粉效率及最终繁殖榕小蜂的后代和榕树种子数量的差异。
1 材料与方法
1.1 研究地点
研究地点位于云南省西双版纳傣族自治州勐腊县勐仑镇,中国科学院西双版纳热带植物园(21°54′N,101°46′E,海拔580 m)。该地处东南亚热带北缘,属于热带季风型气候。年平均气温21.8℃,年平均降水量1 493 mm。干湿季分明,大部分降水(84%)发生在5-10月,极少降水(16%)发生在11月至翌年4月(Caoetal.,2006)。
1.2 观察的榕树和榕小蜂
聚果榕Ficusracemosa隶属于荨麻目(Urticales)桑科(Moraceae)榕属Ficus聚果榕亚属Sycomorus,多年生乔木,雌雄同株,榕果多聚生于老茎瘤状短枝上。聚果榕广泛分布于喜马拉雅南麓至澳大利亚北部热带地区,近期分子证据显示,其整个分布区共有4种传粉榕小蜂,其中分布于西双版纳及周边中南半岛区域的是一种尚未描述的传粉榕小蜂——一种栉颚榕小蜂Ceratosolensp.(Bainetal.,2016)。该蜂隶属于膜翅目(Hymenoptera)榕小蜂科(Agaonidae)栉颚榕小蜂属Ceratosolen,雌雄二型,雌蜂有翅,前足基节上具花粉刷,中胸腹板两侧有一对发达的花粉筐,主动为聚果榕传粉;雄蜂无翅,眼及触角退化(Weiblen,2002)。
榕果内雌雄花异熟。雌花先熟,此时称为雌花期,传粉榕小蜂经苞片口进入果腔传粉和产卵。经过一段时间的发育后,雄花成熟到达雄花期,榕小蜂后代同步发育到成虫期,采集花粉后离开雄花期果,寻找下一株处于雌花期的榕树,开始新的生活史循环(Gaili and Eisikowitch,1968)。
1.3 花柱长度及榕小蜂产卵器长度测量
选取2株处于雌花期的聚果榕,每株树采集雌花期果15果,每果随机取4朵雌花,共120朵,借助安装有测微尺的体视显微镜(NTX-3C)测量花柱长度。此外,再选取2株处于雄花期的聚果榕,每株树采集雄花期果30果,用120筛目的纱网袋单果分装,待榕小蜂自然羽化出蜂后,每果随机取1头小蜂,2株树共取60头,在体视镜下解剖榕小蜂,拉出并测量其产卵器长度。
1.4 榕小蜂搜索、产卵及传粉行为观察
选取2株刚结果的聚果榕,用纱网袋对榕果进行套袋隔离,排除非传粉小蜂的干扰。当榕果发育至雌花期,打开纱网袋,为避免蜂夹死于苞片到达不了果腔,每个果引入2头榕小蜂,进果30 min后,及时采摘进蜂果,带回实验室,借助可连接电脑录像的Digital Microscope(AM4115T-CFVW),观察榕小蜂在果腔内的搜索、产卵和传粉行为,同时录制视频,记录并定量每头小蜂的任意3次各行为所持续时间。每株树观察10个进蜂果,2株树共观察20个果,共计20头蜂。
1.5 榕小蜂个体大小、孕卵量及携粉量测定
当榕果发育至雄花期时,每株树采集30果,用纱网袋单果分装,待榕小蜂自然羽化后,每果随机取1头小蜂,2株树共取60头蜂,在体视镜下解剖,测量其头宽,用于代表小蜂个体大小(Liuetal.,2011);解剖腹部卵巢,计数双侧卵巢中的孕卵量(Dunnetal.,2011;徐睿等,2016);用解剖针移除胸部的双翅及3对足,保留花粉筐位置,用超声波仪振破花粉筐,散落出花粉,然后用细胞计数仪(Multisizer 3)计数花粉量(刘明新等,2017)。
1.6 榕小蜂不同状态及工作时间下卵巢中剩余卵量及花粉筐中剩余花粉量统计
当榕果发育到雌花期,每果控制引入1头榕小蜂,待榕小蜂完全进入苞片后套袋隔离以防其他小蜂进入,并挂上记录进蜂时间的标签,共计引入蜂120果。榕小蜂进入榕果后6 h,先采集30个进蜂果,带回实验室,取出果腔中的榕小蜂,借助体视显微镜解剖、计数其卵巢中剩余卵量,并参照1.5节的方法同步计数花粉筐中剩余花粉量。此外,榕小蜂进果后24 h,再分别采集30个进蜂果,同样的方法计数榕小蜂卵巢中剩余卵量和花粉筐中剩余花粉量。
1.7 雄花期传粉榕小蜂后代及种子数量统计
1.6节中单果控制引蜂试验剩余的60个进蜂果,其中30果在传粉榕小蜂进入榕果6 h后,用注射器从苞片口往果腔中注入0.1 mL浓度为0.05%的噻虫·高氯氟杀虫剂,预试验该浓度的杀虫剂能及时杀死榕小蜂但不影响榕果正常发育(Rajaetal.,2008);另外30果作为榕小蜂进果产卵、传粉24 h的样本,榕小蜂进入榕果后24 h基本奄奄一息快死亡,榕小蜂进果后套上纱网袋,让其自然发育。当试验榕果进入雄花期时,及时采摘,单果分装于纱网袋中。待榕小蜂自然羽化出蜂后,统计每个榕果内榕小蜂后代数量和榕果种子数量。6 h及24 h的试验果最终分别获得7个和18个果的样本量。
1.8 数据分析
运用Excel计算聚果榕雌花花柱长度,其传粉榕小蜂产卵器长度、头宽、孕卵量、携粉量,以及单果内榕小蜂后代及种子数量(mean±SE)。由于数据不呈正态分布,采用Mann-WhitneyU检验比较聚果榕雌花花柱长度与榕小蜂产卵器长度的差异。采用Kruskal-Wallis检验比较榕小蜂搜索、产卵及传粉各行为之间的差异,并比较榕小蜂在不同状态及工作时间后剩余卵量和花粉量,如存在显著差异则用Wilcoxon符号秩检验进行多重比较。采用Pearson相关分析比较榕小蜂个体大小与孕卵量、携粉量的相关性。采用Mann-WhitneyU检验比较榕小蜂进果后6 h与24 h最终繁殖的榕小蜂子代数量差异,采用独立样本t检验(正态分布)比较树间雌花花柱长度差异,以及相同条件下榕果最终繁殖的种子数量差异。所有的数据分析采用R 3.6.1统计软件完成。
2 结果
2.1 聚果榕传粉榕小蜂产卵器长度与花柱长度的匹配程度
聚果榕的雌花花柱长度范围为0.70~2.45 mm,呈单峰分布,树间存在显著差异(t检验:t=-9.50,df=118,P<0.01)。相比较,其传粉榕小蜂的产卵器长度范围在1.56~2.02 mm之间,结果显示花柱长度与榕小蜂产卵器长度有显著差异(Mann-WhitneyU:P<0.01)(图1)。从两者的匹配程度看,榕小蜂产卵器长1.83±0.014 mm(N=60),长于雌花花柱长度(1.47±0.036 mm)(N=120),榕小蜂平均可以利用90.83%雌花产卵。此外,雌花花柱长度的变异(CV=0.27)比榕小蜂产卵器长度的变异(CV=0.06)大,表明部分长花柱的雌花不能被榕小蜂产卵。
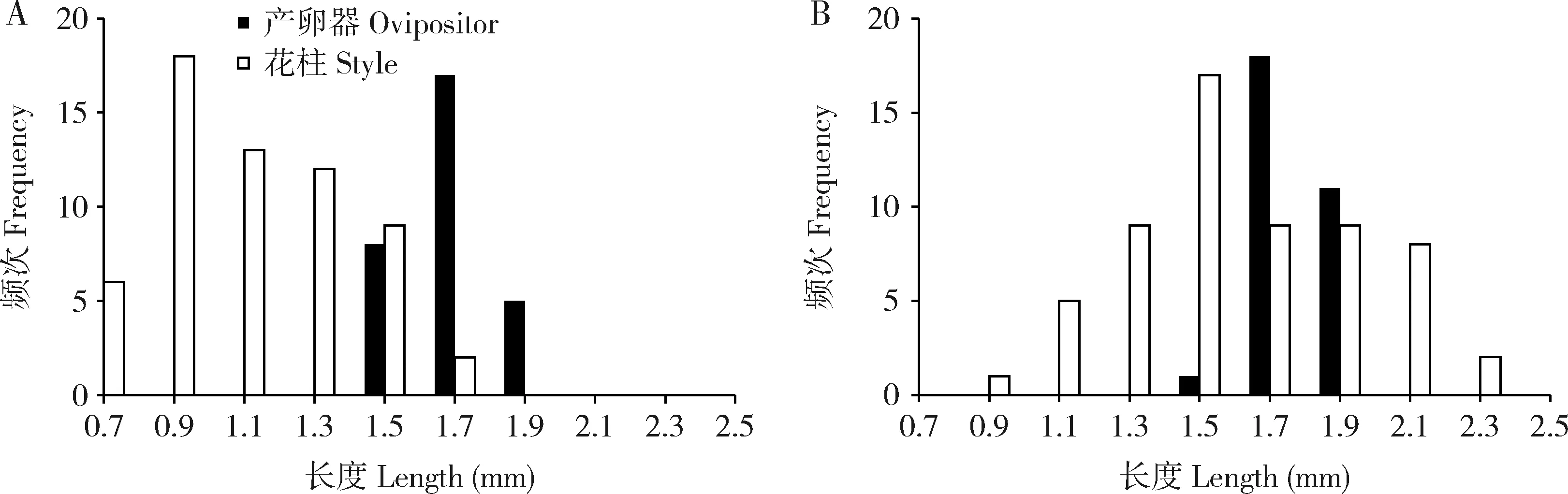
图1 聚果榕雌花花柱长度和榕小蜂(Ceratosolen sp.)产卵器长度的频次分布Fig.1 Frequency distribution of style length of female flowers and ovipositor length of fig wasps (Ceratosolen sp.) in Ficus racemosaA:样树1 Tree 1;B:样树2 Tree 2.
2.2 聚果榕雌花期榕小蜂搜索、产卵及传粉行为
在聚果榕的雌花期,榕小蜂进入榕果后,其产卵器从雌花柱头凹陷处沿花柱插入子房,搜索产卵位点。找到合适的产卵位点后,榕小蜂腹部下沉,产卵于雌花子房的珠被与珠心之间,产卵后拔出产卵器,再搜索下一个产卵位点。当卵巢中孕育的卵基本产完,榕小蜂才开始为聚果榕传粉。传粉行为表现为:前足积极运动,基节上的花粉刷从中胸腹板两侧的花粉筐中刷出花粉。传粉行为与搜索行为交替进行,或连续多次传粉,直至榕小蜂死亡。
榕小蜂在聚果榕雌花期果内的搜索时间平均为26.57±1.74 s(N=120),产卵用时46.07±1.35 s(N=120),传粉用时1.98±0.09 s(N=120);三者之间时间分配存在极显著差异(H=270.51,P<0.01)(图2)。虽然两株树的观察结果存在一定差异,但总体规律一致,说明榕小蜂在雌花期榕果内的搜索、产卵及传粉行为有较稳定的模式。
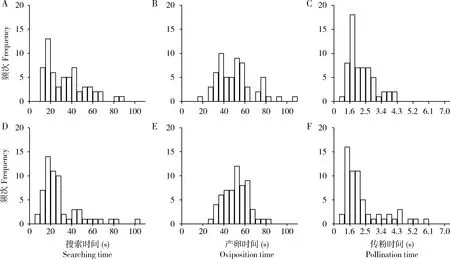
图2 聚果榕雌花期榕小蜂(Ceratosolen sp.)在隐头花序内搜索(A,D)、产卵(B,E)及传粉(C,F)行为的时间分配Fig.2 Time allocation of searching (A,D),oviposition (B,E) and pollination (C,F) behaviors of fig wasps (Ceratosolen sp.) in the receptive female figs of Ficus racemosaA,B,C:样树1 Tree 1;D,E,F:样树2 Tree 2.
2.3 聚果榕传粉榕小蜂个体大小与孕卵量、携粉量的关系
聚果榕传粉榕小蜂个体大小(头宽)平均为0.44±0.003 mm(N=56),刚离开雄花期果的雌蜂卵巢中孕卵量为289±4.74粒(N=56),花粉筐中携带的花粉量为1 887±122.72粒(N=56)。分析结果显示,榕小蜂个体大小与孕卵量之间呈显著正相关关系(r=0.60,P<0.01)(图3:A),即个体大的榕小蜂孕卵量更多。相比之下,榕小蜂个体大小与携粉量相关性不显著(r=0.33,P=0.06)(图3:B)。同时,榕小蜂孕卵量与携粉量之间也没有显著相关性(r=0.32,P=0.06),两株树间的测量结果支持一致性的结论。
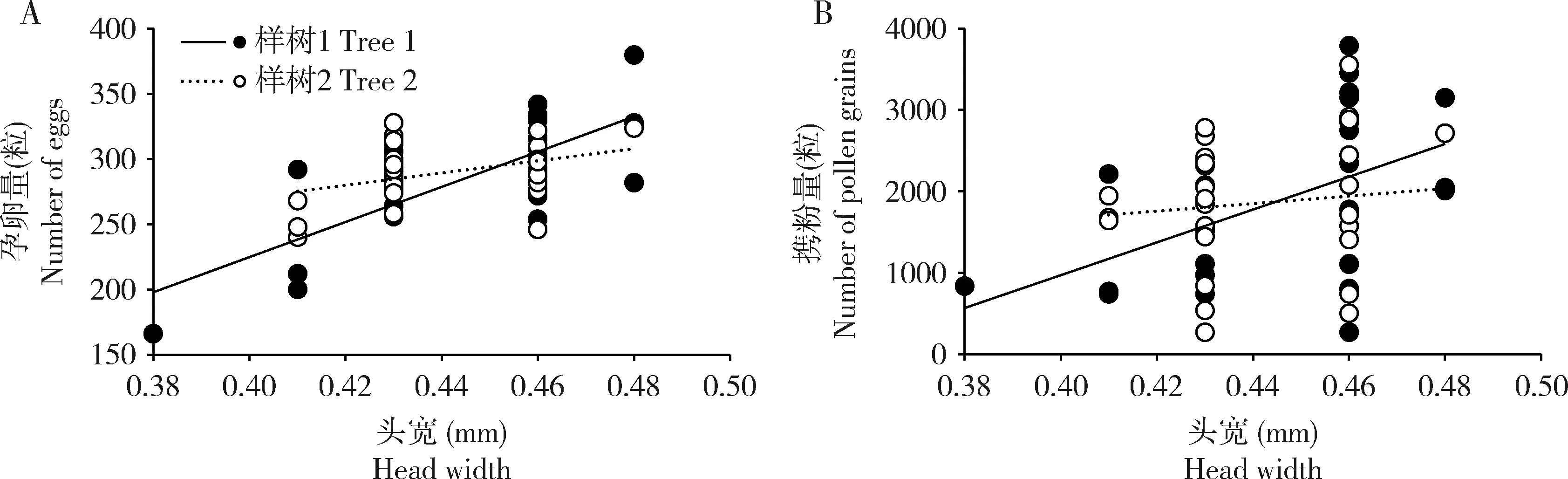
图3 聚果榕传粉榕小蜂(Ceratosolen sp.)个体大小与其孕卵量(A)及携粉量(B)的关系Fig.3 Relationship between the body size of fig wasps (Ceratosolen sp.) and the number of eggs (A) and the number of pollen grains (B) in Ficus racemosa
2.4 聚果榕传粉榕小蜂在不同状态及雌花期不同工作时间下产卵和传粉差异
传粉榕小蜂雌蜂刚离开雄花期果时,卵巢中孕卵量为230±6.62粒(N=23)(图4:A),花粉筐中携粉量为2 190±185.69粒(N=23)(图4:B)。当进入雌花期榕果后6 h,其卵巢中剩余卵量为11±2.53粒(N=22),与刚离开雄花期果的雌蜂相比,卵量显著减少(P<0.01)(图4:A);其花粉筐中花粉量为1 783±158.04粒(N=22),变化不显著(P=0.14)(图4:B)。结果表明聚果榕传粉榕小蜂在进果6 h内主要执行产卵,可产下孕卵量的95.22%。至进果后24 h,榕小蜂卵巢中的卵仅剩8±5.18粒(N=15)(图4:A),与进果6 h的雌蜂相比,剩余卵量无显著变化(P=0.56);花粉筐中花粉量则显著减少至422±60.18粒(N=15)(P<0.01)(图4:B),说明聚果榕传粉榕小蜂在进果后的第6-24小时里主要执行传粉,在此期间产卵较少。与刚羽化的榕小蜂相比,在雌花期果内工作24 h后,榕小蜂卵巢中剩余卵量和携粉量均显著减少(卵量:P<0.01;携粉量:P<0.01),最终可产完孕卵量的96.52%,传完携粉量的80.73%,总体显示聚果榕传粉榕小蜂在雌花期榕果内的产卵及传粉行为主动、完成高效。
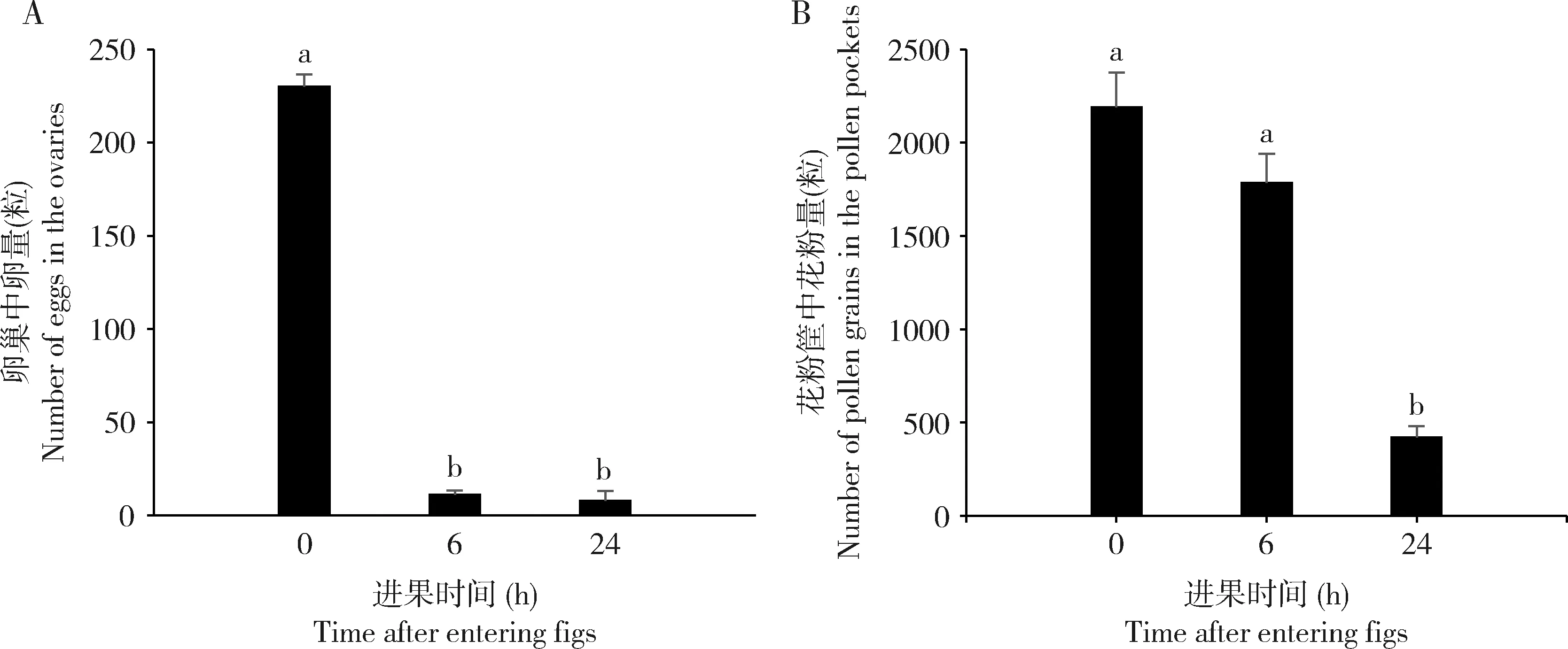
图4 聚果榕传粉榕小蜂进果不同时间后卵巢中卵量(A)及花粉筐中花粉量(B)Fig.4 Numbers of eggs in the ovaries (A) and pollen grains in the pollen pockets (B) of fig wasps (Ceratosolen sp.) entered the receptive figs of Ficus racemosa for different time图中数据为平均值±标准误。柱上不同小写字母示差异显著(P<0.05,Kruskal-Wallis检验)。Data in the figure are mean±SE.Different small letters above bars indicate significant difference (P<0.05,Kruskal-Wallis test).
2.5 聚果榕雄花期传粉榕小蜂子代数量及种子繁殖差异
传粉榕小蜂进入雌花期榕果内工作6 h后,雄花期最终繁殖小蜂后代221±11.56头(N=7),榕树种子23±6.40粒(N=7),榕小蜂后代数量显著多于榕树种子数量。相比之下,榕小蜂在雌花期榕果内工作24 h后,雄花期最终繁殖小蜂后代195±17.34头(N=18),榕树种子983±118.87粒(N=18),繁殖的榕树种子数量显著多于榕小蜂后代数量(图5)。榕小蜂产卵6 h与24 h相比,最终繁殖的后代数量之间没有显著差异(Mann-WhitneyU:P=0.88);但榕小蜂传粉24 h的榕果所繁殖的种子数量比传粉6 h的榕果所繁殖的种子数量显著多(t检验:t=-8.07,df=17.10,P<0.01)。引蜂试验结果证实聚果榕传粉榕小蜂在进入榕果的前6 h基本完成产卵工作,之后的6-24 h则主动为榕树传粉以繁殖种子。
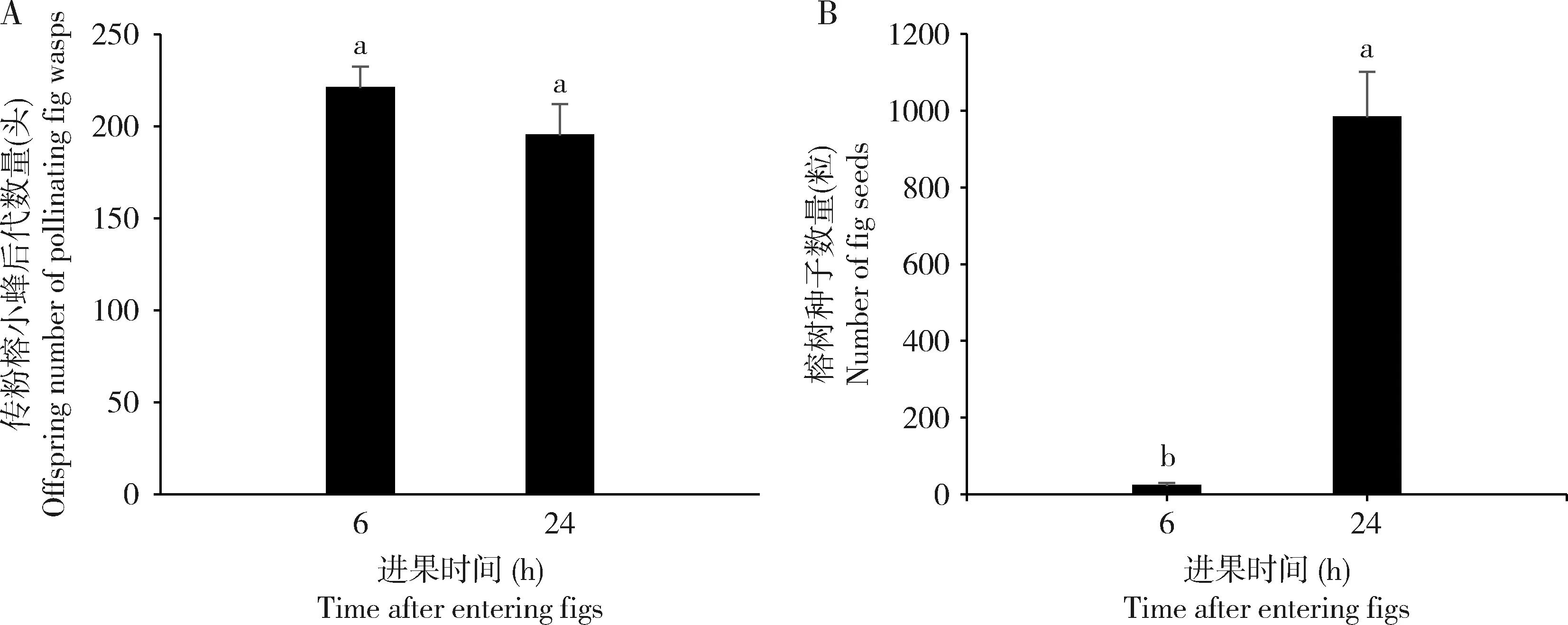
图5 聚果榕传粉榕小蜂进果后不同时间段繁殖的榕小蜂子代数量(A)及榕树种子数量(B)Fig.5 Numbers of wasp offspring (A) and fig seeds (B) produced by fig wasps (Ceratosolen sp.) entered the receptive figs of Ficus racemosa for different time图中数据为平均值±标准误。柱上不同小写字母示差异显著(P<0.05)(榕小蜂子代数量:Mann-Whitney U检验;榕树种子数量:独立样本t检验)。Data in the figure are mean±SE.Different small letters above bars indicate significant difference in the number of wasp offspring by Mann-Whitney U test and the number of fig seeds by independent samples t-test (P<0.05).
3 讨论
榕树和榕小蜂都必须利用榕果内的雌花资源来完成繁殖,从这个角度来说,它们二者之间存在繁殖利益冲突,榕树和榕小蜂之间权衡繁殖利益是维持该互惠共生体系稳定的关键。短产卵器假说(short-ovipositor-length hypothesis)认为,榕果内的雌花有短花柱和长花柱之分,由于短花柱雌花的花柱长度与传粉榕小蜂的产卵器长度相匹配,所以传粉榕小蜂将卵产于短花柱雌花的子房中以繁殖自身后代,而长花柱雌花则被传粉受精发育成为种子(Ganeshaiahetal.,1995)。但是相关的研究显示,不同的榕树及其传粉榕小蜂系统存在差异,部分榕-蜂系统中榕小蜂产卵器长度足以到达大多数雌花子房产卵(Nefdt and Compton,1996)。本研究中聚果榕传粉榕小蜂的产卵器比90%的雌花花柱长,与雌雄同株钝叶榕、高榕及它们的传粉榕小蜂共生体系相似,传粉榕小蜂能产卵于绝大多数雌花子房中(张媛等,2010;Chenetal.,2013)。此外,聚果榕雌花期,雌花柱头间紧密粘连,具有联合柱头平台(synstigma),这有利于花粉管侧向生长,提高传粉和受精效率(Jousselin and Kjellberg,2001;Jousselinetal.,2003;Teixeiraetal.,2018)。本研究发现传粉榕小蜂进入榕果后,搜索雌花,优先产卵,每次平均产卵用时46 s,前6 h小蜂主要执行产卵工作,之后集中传粉,其行为主动,连贯高效,单次传粉仅用时2 s。这一结果主要得益于先进显微视频设备的运用,第一次展示了雌雄同株榕树雌花期果内榕小蜂传粉、产卵的行为及时间分配,为我们理解榕-蜂如何分配繁殖雌花资源,稳定繁殖共存提供了直接证据。
先前的研究由于难于清楚、长时间观察传粉榕小蜂在雌花期榕果内的活动行为,往往只统计榕小蜂剩余卵量和花粉量,以及最终成功繁殖的小蜂后代及种子数量来倒推榕小蜂可能的传粉、产卵行为(Herreetal.,2008;徐睿等,2016)。本研究直接对传粉榕小蜂行为进行观察,并统计榕小蜂在不同传粉状态及工作不同时间段的产卵及传粉情况(图2和4),结果一致性地支持:进入榕果内前6 h,榕小蜂主要执行产卵行为,能够产完孕卵量的95%,优先保证自身的繁殖;之后的6-24 h,小蜂主要执行传粉行为,可传完携粉量的80%。单果控制性引蜂试验也证实了雌花期传粉榕小蜂产卵及传粉效率,以及雄花期最终成功繁殖的小蜂后代及种子数量之间的对应关系(图5)。结果很好地支持:执行产卵的前6 h,繁殖显著多的榕小蜂后代;而后的6-24 h,则繁殖显著多的榕树种子。雌雄同株不同属传粉榕小蜂以及雌雄异株雌雄果内榕小蜂的繁殖行为值得进一步对比研究。
在自然界中,主动的传粉模式很少见,多发生于互惠共生系统中,通常花及传粉昆虫之间有特化的传粉结构和行为(Kjellbergetal.,2001;Cooketal.,2004;Wangetal.,2014)。聚果榕传粉榕小蜂除了具有主动传粉的行为外,还有专门用于传粉的花粉刷与花粉筐,其通常将花粉装于胸部腹面两侧的囊状花粉筐中进行携带。本研究表明聚果榕传粉榕小蜂的个体大小与孕卵量呈显著正相关关系(图3),这与高榕传粉榕小蜂的研究结果(徐睿等,2016)一致。然而,本研究还发现聚果榕传粉榕小蜂个体大小与携粉量相关性不显著,孕卵量和携粉量之间没有相关性(图3),探明了传粉榕小蜂个体大小与其孕卵量及携粉量之间的关系。
在对传粉者有利的情况下,主动传粉行为才可能被保留并得到进化(Jousselinetal.,2003)。有研究报道榕小蜂幼虫以传粉后形成的胚乳组织为食,因此,传粉受精可确保其幼虫发育过程中有充足的食物(Verkerke,1989;Jousselinetal.,2003;Tarachaietal.,2008)。对分布在西双版纳的垂叶榕及分布于中美洲的几种榕树上,控制性引蜂试验发现榕树存在寄主惩罚(host sanctions),没有授粉的榕果会提前脱落,推测榕小蜂进入榕果后首先进行传粉,以保证榕果的正常发育(管俊明等,2007;Jander and Herre,2010)。Jansen-González等(2012)对F.citrifolia的传粉榕小蜂Pegoscapussp.发育过程进行了一系列研究,认为其幼虫的取食策略随生长发育而改变,首先取食植物珠心组织,进入3龄幼虫期后取食植物胚乳;当榕果缺乏授粉时,其3龄幼虫期将取食榕果内其他资源,但死亡率显著升高。总之,主动传粉有利于榕小蜂后代的正常发育。本研究揭示了聚果榕传粉榕小蜂进入雌花期榕果后优先产卵、之后传粉的生物学现象,这一策略保证了榕-蜂双方的成功繁殖。本研究的结果为深入探究榕-蜂共生体系的机制奠定了基础。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
学苑创造·A版(2022年2期)2022-01-29
科学之友(2021年12期)2021-12-23
环境昆虫学报(2021年4期)2021-08-13
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
大科技·百科新说(2018年3期)2018-04-11
小学生作文选刊·中高年级版(2017年4期)2017-04-24
创新作文(3-4年级)(2016年9期)2016-12-22
热带农业科学(2015年1期)2015-02-15
