
布氏田鼠种群密度对不同放牧方式及植被变化的响应
2020-10-17 13:23郭乾伟杨素文付和平武晓东张卓然马崇勇
草原与草业 2020年3期
郭乾伟,袁 帅,杨素文,纪 羽,付和平,武晓东,*,张卓然,马崇勇,金 国
(1.内蒙古农业大学草原与资源环境学院/内蒙古农业大学啮齿动物研究中心,呼和浩特 010018;2.内蒙古农业大学生命科学学院,呼和浩特 010019;3.内蒙古农业大学草地资源教育部重点实验室,内蒙古 呼和浩特 010011;4.内蒙古自治区草原工作站,内蒙古 呼和浩特 010020;5.内蒙古自治区锡林郭勒盟东乌珠穆沁旗草原工作站,乌里雅斯太 026300)
放牧作为一种最普遍、最经济的草地管理利用方式,不仅对草地生态系统有重要影响[1~2],同时对草原啮齿动物种群也产生了一定的影响。放牧改变了土壤和植被的结构和组成[3],一方面影响一些啮齿动物的栖息地环境,导致啮齿动物的迁徙[4];另一方面植物的结构和组成变化后,对啮齿动物取食行为也有所影响[5~7]。啮齿类动物作为草地生态系统中很重要的成员,不仅充当着消费者的角色,而且还是物质与能量的传递者[8~9]。布氏田鼠(Lasiopodomysbrandtii)作为内蒙古典型草原主要害鼠之一,繁殖能力强,繁殖期长达6~7个月,且主要集中在春、夏两季[10~11]。布氏田鼠种群数量增加会对草地产生严重的危害[12~13]。国内外对布氏田鼠种群的研究相对时间较长,内容也多样化,在生态学层面主要集中在对其种群密度[14]、雌雄比例[15]以及繁殖率[16]的研究方面;在分子生物学层面集中在能量代谢[17~18]、遗传多样性[19~20]等方面。本文通过对2016~2018年每年7月份内蒙古锡林郭勒大针茅(Stipagrandis)草原不同放牧方式下布氏田鼠种群密度与植物种群特征变量的分析,明确不同放牧方式下布氏田鼠种群密度与植物种群的关系,旨在探索大针茅草原中布氏田鼠种群数量变化的主导生物因子,为揭示典型草原野生动植物互作机制提供参考。
1 研究区概况
研究区位于内蒙古自治区锡林郭勒盟东乌珠穆沁旗,地理坐标为E115°30′~116°30′、N44°42′~45°15′,草地类型为典型草原,草层高10~20cm,主要植被种类为大针茅、羊草(Leymuschinensis)、糙隐子草(Cleistogenessquarrosa)、多根葱(Alliumpolyrhizum)、栉叶蒿(Neopallasiapectinata)等。气候属于典型的北温带大陆性气候,其特征为冬季寒冷风大、夏季水热同期;年均气温1.6℃,1月平均气温-18.9℃,7月平均气温21℃,极端最高气温39.7℃,最低气温-40.7℃,年日平气温0.7℃。年生长期(日均5℃以上)95d,无霜期为120d。年降水量300mm左右,主要集中在6~8月份,占年降水量的70%;年蒸发量在3000mm以上[21]。
2 研究方法
2.1 试验设计及啮齿动物取样方法
2016~2018年,在锡林郭勒大针茅草原区,采用完全随机区组试验设计,将9个小区划分为3个区组,即三次重复,每个区组中设有3个处理,即三种不同放牧方式,分别为连续放牧(CG,小区面积4hm2,放牧绵羊6只)、按月轮牧(MG,小区面积4hm2,放牧绵羊6只)和禁牧(GE,小区面积4hm2)。每个区组内三种放牧方式完全随机排列。连续放牧为自由放牧,按月轮牧将样地分为2个等面积小样区,在2个小样区间进行周期为1个月的轮流放牧,禁牧作为对照不进行放牧。每个样区设置3个啮齿动物调查样方,分别进行同心圆式取样,同心圆半径分别为7m、14m、21m,每个同心圆布放25只活捕笼(27.5cm×12.5cm×12.5cm,图1),以新鲜花生米为诱饵,采用标志重捕法捕获啮齿动物,每年7月份进行一次试验,每次连续调查4d,每天6时、12时和18时各查笼1次,并记录捕获布氏田鼠的位置、性别、体重(g)及繁殖状况,使用PIT芯片对捕获的布氏田鼠进行皮下注射标记。
2.2 植被调查
2016~2018年每年的7月,在各处理的动物取样区分别随机选取3个样方(面积为0.25m2),记录每个样方的植被盖度、植物种名、高度(cm)和密度,并称量每个物种的地上生物量鲜重(g)及风干重(g)。
2.3 数据分析
用百笼捕获率表示布氏田鼠种群密度,以100笼所捕获的不重复啮齿动物计算,公式为:
式中:P为捕获率;N为所捕获鼠的数量;H为总的布笼数;n为连续布笼天数。
采用混合线性模型将年份作为随机效应,不同放牧方式作为固定效应,分析不同放牧方式下布氏田鼠种群密度的差异性,以及不同放牧方式下植物高度、密度和地上生物量的差异性。选择存在显著差异的变量以及样方的植被盖度与布氏田鼠种群密度进行典型相关分析,所有数据分析显著水平为0.05。统计分析采用SAS 9.0完成,运用Sigmaplot 12.5软件完成作图。
3 结果与分析
3.1 不同放牧方式下布氏田鼠的种群密度
本研究期间在禁牧(GE)、连续放牧(CG)和按月轮牧(MG)三种处理样地共布放活捕笼2700笼日,捕获布氏田鼠97只,总的百笼捕获率为3.59%。其中,捕获雌性布氏田鼠41只,占总数的42.27%;雄性56只,占总数的57.73%。对三种不同放牧方式下布氏田鼠种群密度(百笼捕获率)分析可知,布氏田鼠种群密度在连续放牧区显著低于禁牧区(F=5.59,P<0.05,图2)。
3.2 不同放牧方式下植物种群特征分析
本研究期间在三种放牧处理植物种群中,主要植物有羊草、糙隐子草、大针茅、栉叶蒿、苔草(Carexbrevicuspis)、多根葱、野韭(Alliumramosum)、二裂委陵菜(Potentillabifurca)、小叶锦鸡儿(Caraganamicrophylla)、银灰旋花(Convolvulusammannii)和黄芪(Astragalusmembranaceus)。通过对不同放牧方式下植物的高度、密度、地上生物量以及样方植被盖度进行差异性分析可知(表1),大针茅密度在连续放牧区和按月轮牧区均高于禁牧区,而在连续放牧与禁牧区之间的差异达到了极显著水平(F=8.08,P=0.0021);栉叶蒿密度在禁牧区和按月轮牧区显著高于连续放牧区(F=10,P=0.0008);多根葱密度在连续放牧区和按月轮牧区显著高于禁牧区(F=11.15,P=0.0005);多根葱地上生物量在连续放牧区和按月轮牧区显著高于禁牧区(F=6.69,P=0.0045);野韭地上生物量在禁牧区和连续放牧区均高于按月轮牧区,而在连续放牧区和按月轮牧区之间出现显著差异(F=4.08,P=0.0326);小叶锦鸡儿高度在禁牧区和按月轮牧区均高于连续放牧区,而在连续放牧区和按月轮牧区之间出现显著差异(F=3.46,P=0.0494);其他植物指标在三种处理间差异均未达到显著水平。样方植被盖度在3个样区中也未达到差异显著水平(F=0.30,P=0.7466),但禁牧区植被盖度大于连续放牧区和按月轮牧区。
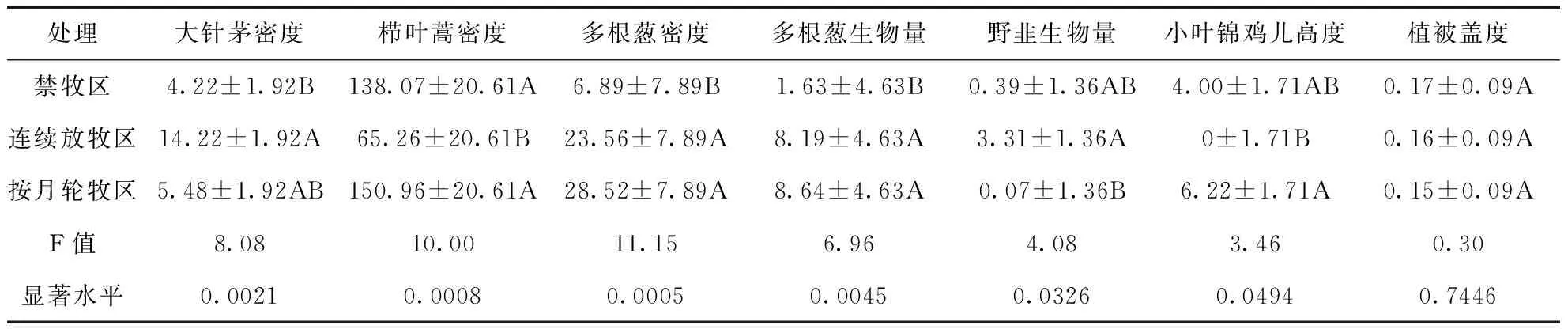
表1 不同放牧方式下植物特征指标差异性分析
3.3 布氏田鼠种群密度与植物种群特征变量的相关分析
为了明确布氏田鼠种群密度与植物种群因子变量之间的关系,应用上述三种处理样地间植物种群特征变量差异性分析结果,选取6个存在显著差异的植物种群指标变量(大针茅密度、栉叶蒿密度、多根葱密度、多根葱地上生物量、野韭地上生物量和小叶锦鸡儿高度)以及样地植被盖度与布氏田鼠种群密度进行典型相关分析,将布氏田鼠种群密度设为Y1、大针茅密度设为X1、栉叶蒿密度设为X2、多根葱密度设为X3、多根葱地上生物量设为X4、野韭地上生物量设为X5、小叶锦鸡儿高度设为X6和植被盖度设为X7共8个变量。通过对这8个变量进行典型相关分析可知,动物指标与植物指标的第一典型相关变量的累积方差贡献率为100%,其显著性检验达到极显著水平(P<0.001),因此得到动物指标和植物指标标准化表达式为:
DW(动物指标)=-Y1
ZW(植物指标)=0.2295X1+0.2730X2-0.2037X3-0.0660X4+0.5501X5-0.0649X6+0.5845X7
由以上表达式可知,动物指标第一典型变量由Y1(布氏田鼠种群密度)决定,植物指标第一典型变量主要由X5(野韭地上生物量)和X7(植被盖度)决定。Y1与X5和X7符号相反,即布氏田鼠种群密度与野韭地上生物量以及植被盖度呈负相关关系。
3.4 不同放牧方式下布氏田鼠种群密度与植物特征变量的相关分析
为了探究在不同放牧方式下布氏田鼠种群密度与植物特征变量的关系,对上述8个指标在不同放牧方式下的典型相关进行了分析。分析结果表明,禁牧区布氏田鼠种群密度与上述七项植物指标的典型相关达到极显著水平(F=4.30,P=0.0052),连续放牧区和按月轮牧区布氏田鼠种群密度与上述七项植物指标的典型相关均未达到显著水平(F=1.65,P=0.1834;F=0.73,P=0.6503)。在禁牧区动物指标与植物指标的第一典型相关变量的累积方差贡献率为100%,其显著性检验达到极显著水平(P<0.01),因此得到的动物指标和植物指标标准化的表达式如下:
DW(动物指标)=-Y1
ZW(植物指标)=-0.0603X1+0.2906X2+0.5388X3-1.4212X4+0.0627X5+0.6142X6+0.5473X7
由以上表达式可知,在禁牧区动物指标第一典型变量由Y1(布氏田鼠种群密度)决定,植物指标第一典型变量主要由X3(多根葱密度)、X4(多根葱地上生物量)、X6(小叶锦鸡儿高度)和X7(植被盖度)决定。Y1和X4符号相同,即布氏田鼠种群密度与多根葱地上生物量呈正相关关系;Y1和X6、X7、X3符号相反,即布氏田鼠种群密度与小叶锦鸡儿高度、植被盖度和多根葱密度呈负相关关系。
4 讨论
布氏田鼠在我国仅分布在内蒙古呼伦贝尔和锡林郭勒的典型草原区[22~23],由于受气候变化和人为干扰的影响,其种群数量极不稳定[24~25],历史上曾经多次出现的种群爆发,对草地造成严重危害[26]。多年来,关于布氏田鼠种群数量与草地植物之间的关系引起许多学者的关注[27~28]。Augustine和Springer的研究结果表明,家畜放牧的地区会降低小型啮齿动物的种群数量[14]。Schmidt等研究认为,放牧地区小型哺乳动物的种群密度明显低于未放牧的地区[29]。李国梁等在锡林郭勒草原毛登牧场研究显示,进行长期的轻度放牧和适度放牧会对布氏田鼠的种群密度产生负面影响,主要由于绵羊与布氏田鼠喜食植物有很大的重叠,而长期的放牧会导致布氏田鼠食物的短缺,并建议进行适当的轮牧制度来控制布氏田鼠种群密度[30]。在有蹄类家畜放牧时,家畜进行采食时会减少田鼠栖息地的食物资源,家畜在活动时通过践踏会对田鼠的洞穴产生影响,从而降低了田鼠的种群密度[3,31~32]。本研究也表明,在连续放牧和按月轮牧样区布氏田鼠种群密度均低于禁牧样区,而且在连续放牧区与禁牧区之间出现显著差异,说明放牧对布氏田鼠种群数量具有一定的抑制作用,这与上述研究结果相似。但是也有研究认为,放牧对布氏田鼠种群密度产生正面影响。施大钊研究提出,布氏田鼠生境的选择与放牧强度存在正相关[33]。任修涛等研究显示,放牧草场中的布氏田鼠种群密度显著高于禁牧草场[24]。刘书润在东乌珠穆沁旗典型草原的研究提出,过度放牧导致的退化草场中植被盖度的降低,为布氏田鼠繁殖创造了良好条件[23]。李鹏年等研究认为,围栏封育草场会使草地植被恢复、植被盖度升高,从而导致布氏田鼠栖息地环境的改变,并且会对布氏田鼠的种群数量起到控制作用[34],这就意味草场封育可以抑制布氏田鼠种群数量。本研究中,布氏田鼠种群密度与植被盖度是负相关关系,也就是说植被盖度的增加就意味着布氏田鼠种群数量的降低,但是放牧区和禁牧区中植被盖度的差异性分析中未见显著,所以禁牧区布氏田鼠种群数量也未见降低。因此,本研究结果与上述研究结果也是一致的。总结前人的上述研究结果结合本研究结果来看,放牧家畜与布氏田鼠会出现食物资源的竞争,降低了布氏田鼠的食物可用性导致布氏田鼠种群数量的降低,而禁牧由于植被的恢复使植被盖度增加,布氏田鼠的捕食风险也大大增加导致了布氏田鼠种群数量也会降低。这样看来,就典型草原而言,本研究认为当食物可用性对布氏田鼠种群数量占主导作用时,放牧会抑制布氏田鼠种群密度;当捕食风险对布氏田鼠种群数量占主导时,禁牧会抑制布氏田鼠种群密度。草地利用(放牧)与不利用(禁牧)都可以抑制布氏田鼠种群数量,这是食物可用性和捕食作用相互权衡的结果。
有研究表明,啮齿动物对不同气味会表现出不同的行为特征[35~36]。从本研究对布氏田鼠种群密度与植物特征指标的典型相关分析可知,随着野韭地上生物量的升高布氏田鼠种群数量会有降低的趋势(显著负相关),而关于布氏田鼠对野韭的采食描述的文献甚少,并且本研究中野韭地上生物量在样方植被地上生物量中占据较少的数量,因此本文推测野韭所散发的气味对布氏田鼠有趋避作用。从本研究对三种草地利用方式样区的布氏田鼠种群密度与植物特征指标的典型相关分析中可知,只有在禁牧区中布氏田鼠种群密度与植物特征指标出现显著关系:随着多根葱地上生物量的增加,布氏田鼠种群数量有增加的趋势;随着多根葱密度和小叶锦鸡儿高度的增加,布氏田鼠种群数量则有减少的趋势。本研究在对各样地植物特征指标进行差异性分析时,发现禁牧区中多根葱地上生物量、多根葱密度和小叶锦鸡儿高度均低于放牧样区,而布氏田鼠种群密度却高于放牧样区,这可能是由于布氏田鼠对草丛高大、繁茂的环境有回避行为[33],因此在禁牧区中多根葱密度和小叶锦鸡儿高度对布氏田鼠种群密度的负影响消减了多根葱地上生物量对布氏田鼠的正影响,即多根葱密度和小叶锦鸡儿高度对布氏田鼠种群密度的负影响要大于多根葱地上生物量对布氏田鼠的正影响作用。有关其影响机制还有待于进一步探讨。
5 结论
综合上述研究结果,可以认为草地利用方式通过对布氏田鼠食物可用性以及捕食风险的影响来影响布氏田鼠种群密度,是二者权衡的结果,因此草地利用方式并不是直接影响布氏田鼠种群消长的唯一因子,一定存在更为关键的限制因子会对布氏田鼠种群数量波动产生重要的影响。这一问题也是典型草原野生动植物互作关系研究的重要问题,值得我们继续深入探讨。
猜你喜欢
作文大王·低年级(2021年11期)2021-12-22
安徽农业科学(2021年16期)2021-08-30
西藏艺术研究(2021年3期)2021-06-02
小天使·一年级语数英综合(2021年2期)2021-05-08
南方农业·中旬(2020年9期)2020-11-30
甘肃教育(2020年22期)2020-04-13
甘肃教育(2020年20期)2020-04-13
小学生(看图说画)(2020年4期)2020-04-01
实践·党的教育版(2019年9期)2019-11-06
绿色科技(2017年15期)2017-09-01
