
四川白水河自然保护区大熊猫主食竹种群特征及其影响因素
2020-11-04 11:10彭科陈旭张宣房超杨彪
四川林业科技 2020年5期
彭科, 陈旭, 张宣, 房超, 杨彪
1. 彭州市规划和自然资源局,四川 彭州 611930;
2. 成都畅新环保科技有限公司,四川 成都 610047;
3. 西华师范大学,西南野生动植物资源保护教育部重点实验室,四川 南充 637002
大熊猫(Ailuropodamelanoleuca)是我国珍稀濒危特有种,被称为国宝,同时也是全球生物多样性保护的旗舰物种和伞护物种,对大熊猫的保护研究一直受到国内外学者的广泛关注。大熊猫原本为大型的食肉动物,随着自然环境的改变,大熊猫在数千年的演化与适应下逐渐转变为以亚高山小径竹类为主食的植食性动物[1-2]。
据全国第四次大熊猫调查:我国野生大熊猫主要分布于秦岭、岷山、邛崃山、大相岭、小相岭和凉山等六大山系,野生大熊猫取食竹种共有8属38种,然而不同区域的优势竹种不同,大熊猫主要取食竹种亦有差异[1]。大熊猫主食竹是大熊猫繁衍和发展的基础和保证,开展大熊猫主食竹的调查研究对于大熊猫的保护具有重要的意义[3]。以往研究已对大熊猫主食竹的多样性[4]、分类与分布[5-6]、天然更新[7]、种群结构[8]、种群动态及其影响因素[9]、生物量分配与模拟[10]、生理生化特征[11]、营养成分[12-14]、粗蛋白、粗脂肪和氨基酸[15]及其季节动态[16]、适口性[14]、自然灾害的影响[17]、对全球变化的响应(如CO2浓度升高和N沉降[3])、全球变化情景下竹子分布区的可能变化[18]、病虫害[19]、人为管理与干扰影响[13,20]等方面做了相关报道,但这些研究仅局限于六大山系的部分保护区,如王朗保护区[7]、卧龙保护区[8,9,12]、秦岭保护区[11,16]、大风顶保护区[20]、栗子坪保护区[10]、鞍子河保护区[14]等,研究区域不平衡。已有研究指出大熊猫取食竹的种类存在明显的区域差异[1,21]。加之20世纪80年代岷山山系大熊猫主食竹曾出现大量开花致死现象,2005年以来又陆续发现岷山山系大熊猫主食竹有新的开花现象。因此,有必要加紧积累不同大熊猫分布区的大熊猫主食竹研究数据从而为不同区域大熊猫的科学保护提供重要的数据支撑。
四川白水河国家级自然保护区位于四川盆地西北边缘的彭州市龙门山和小鱼洞镇境内,是离成都市最近(仅50 km)的大熊猫自然保护区,多年来保护区虽开展了不少针对大熊猫的保护工作,但针对大熊猫的研究报道极其缺乏,仅见于李华东[22]对保护区内大熊猫生存环境评价及研究的报道,目前保护区内关于大熊猫主食竹的相关研究仍较为缺乏。本研究通过对该保护区大熊猫主食竹开展调查,旨在查清该区域大熊猫主食竹的种类、种群特征及其受环境因子(海拔和郁闭度)的影响,以期为不同大熊猫分布区的大熊猫主食竹研究积累基础数据,同时也为该保护区大熊猫的保护提供数据支撑和理论依据。
1 研究区域与方法
1.1 研究区域概况
四川白水河国家级自然保护区(下文简称“保护区”)位于龙门山褶皱带的中南段、横断山东部,地理位置北纬31°10’—31°29’、东经103°41’—103°57’。该区域年平均降水量1 500 mm左右,主要集中在7、8、9这3个月;年平均气温为15.6 ℃,年平均无霜期277 d,日照仅711 h。保护区最低海拔1 481 m(锅框岩)、最高海拔4 814 m(太子城),海拔高差跨度极大、地形地貌复杂,温暖湿润,降水丰富,云雾多日照少,且富垂直变化的气候条件,形成了水热条件各异的多种生态环境。土壤具有明显的垂直地带性,自下而上是山地黄棕壤、暗棕壤、棕色针叶林土、亚高山草甸土和高山草甸土。植被类型完整丰富,有亚热带山地常绿、落叶阔叶混交林、落叶阔叶林、针阔混交林、高山山地硬叶常绿阔叶林、暗针叶林、高山灌丛和高山草甸等。
1.2 野外调查
野外调查采用样线法和样方法进行。在收集保护区本底调查等历史资料的基础上共设置25条调查样线(见图1),每条样线两侧50 m内布设不少于4个20 m×20 m的样方进行调查。2018年5—6月,在白水河保护区及周边共设置植物样方70个,竹子样方197个。由于第一次调查未能发现所有竹种的竹笋,同年9—10月在同一区域设置植物样方36个,竹子样方123个以补充调查。另外,调查期间在保护区边界外围发现有与保护区内同种的“小径竹”分布,故做样方调查以增加低海拔区域的样方,所有调查样方最低海拔1 240 m,最高海拔3 200 m。对样方中的竹子实测其克隆分株数(笋、1年生克隆植株和超过1年生克隆植株)、基径、高度,采用GPS和罗盘仪测量海拔、坡度和坡向等环境因子,采用“多点人工目测”方式测定郁闭度,同时记录竹子的病虫、开花、大熊猫和其他动物取食等情况。
1.3 数据统计与分析
以本研究获得的所有“小径竹种类”来组织研究区域的全部竹种名录。在做环境因子相关分析时,由于刺黑竹(Chimonobambusa neopurpurea)、箬 叶 竹(Indocalamus longiauritus)和 柺 棍 竹(Fargasia robusta)的分布海拔范围较窄,其调查得到的样方数达不到统计的最小样本数(各仅有6个小样方),故仅对调查样本量较多的白夹竹(Phyllostachys bissetii)、短锥玉山竹(Yushania brevipaniculata)、冷箭竹(Bashania fangiana)和细枝箭竹(Fargesia stenoclada)4种竹种做环境要素(海拔和郁闭度)与其种群特征之间的关系分析,以初步评估研究区域种群分布较广的竹种受环境因子的变化情况。
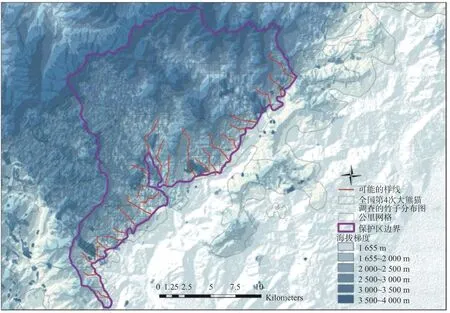
图1 调查样线分布图Fig.1Distribution of survey sample lines in Baishuihe National Nature Reserve, Sichuan
所有数据的统计检验分析在Origin Pro 8.0软件上完成,数据作图在 Excel 2016软件上完成。
2 结果和分析
2.1 种类与分布
本研究共调查到竹类6属7种,分别为刚竹属(Phyllostachys)的白夹竹、方竹属(Chimonobambusa)的刺黑竹、玉山竹属(Yushania)的短锥玉山竹、箬竹属(Indocalamus)的箬叶竹、巴山木竹属(Bashania)的冷箭竹以及箭竹属(Fargesia)的拐棍竹和细枝箭竹。7种竹子海拔分布范围主要在3 000 m以下,郁闭度在0.3~0.9之间。本研究中仅发现短锥玉山竹和细枝箭竹2种竹子的茎叶被大熊猫取食后留下食痕,7种竹子均未发现被其他动物取食的痕迹,未发现有开花和病虫害现象(见表1)。
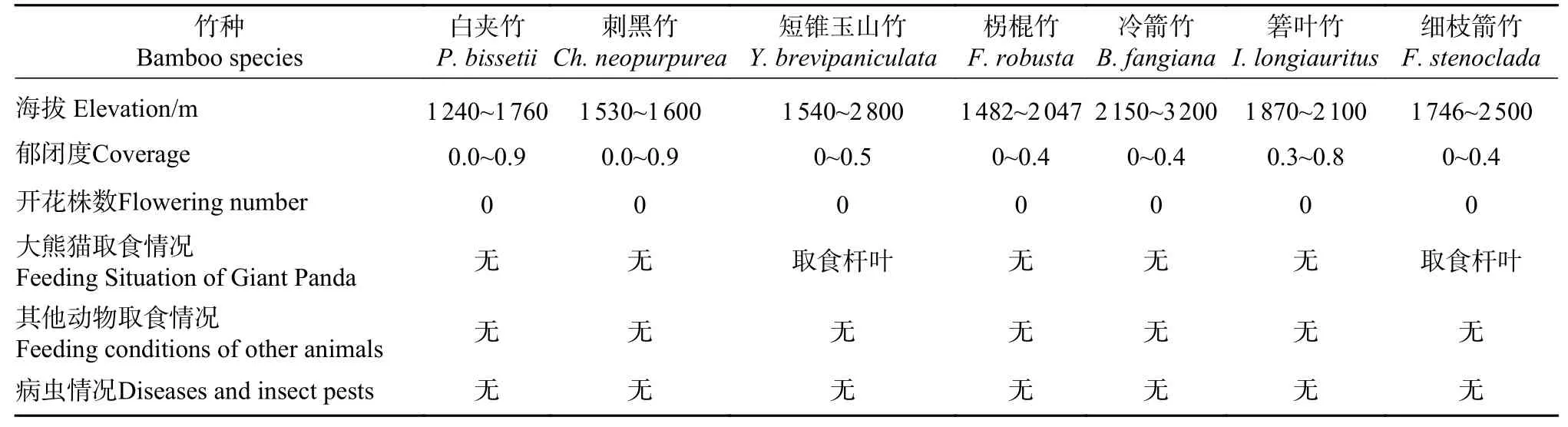
表1 四川白水河国家级自然保护区内的竹子种类及概况Tab.1Bamboo species and general information in Baishuihe National Nature Reserve, Sichuan
2.2 种群特征
2.2.1 竹笋与幼竹
竹笋是评价竹子种群更新的重要指标之一。本研究期间调查样方中未发现刺黑竹和箬叶竹的竹笋。其原因一方面可能与刺黑竹种群较少导致发笋情况较少有关,据保护区以往调查资料得知虽该区域刺黑竹的发笋期在农历8月中旬之后,但本次调查获得的刺黑竹的样方数较少,刺黑竹种群数量不多从而导致发笋情况较少可能是本次调查难以见到其发笋情况的重要原因;另一方面可能与箬叶竹新笋识别难度较大有关,箬叶竹的新克隆分株是一边生长,一边发侧枝,野外调查时极易忽略是“新笋”。而白夹竹、短锥玉山竹、柺棍竹、冷箭竹和细枝箭竹均有竹笋出现,但其竹笋样方的百分比均小于50%(见表2)。
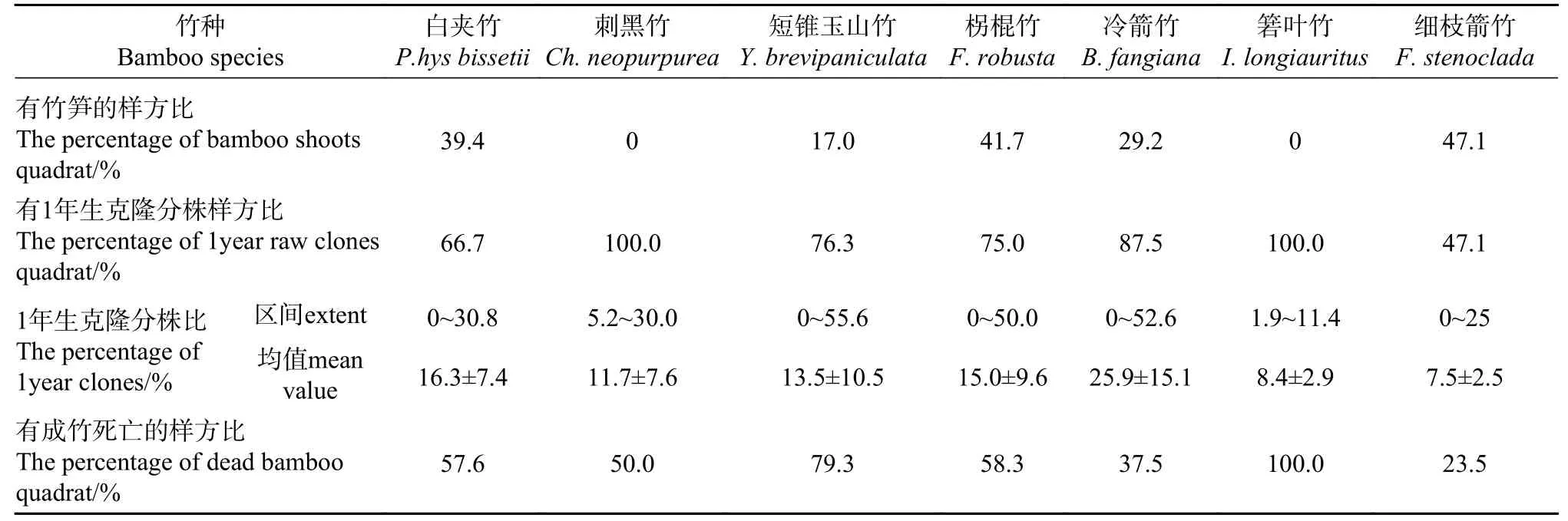
表27 种竹子的竹笋、1年生克隆分株、死亡成竹Tab.2Bamboo shoots, 1-year-old clonal ramets and dead bamboo of 7 bamboo species
1年生克隆分株是竹子种群更新扩展的新生力量,也是经过自然淘汰后保留的最具生命力的克隆分株。虽本研究在调查样方中7种竹子均有出现“自然死亡的成熟克隆分株”,其中出现箬叶竹死亡的样方占比最高(100%),然而除细枝箭竹(47.1%)外,其余6种竹子出现1年生克隆分株的样方比均超过65%,而刺黑竹和箬叶竹出现1年生克隆分株的样方比达到100%(见表2),表明调查区内竹子更新良好。
西南山地竹子是以克隆繁殖为主的多年生植物,1年生克隆分株比是衡量竹子种群更新和种群稳定性的重要指标。研究期间发现1年生克隆分株比最高的是冷箭竹,其均值达25.9%;其次是白夹竹,达16.3%;再次是柺棍竹、短锥玉山竹和刺黑竹,分别为15.0%、13.5和11.7%;而箬叶竹和细枝箭竹的1年生克隆分株比的均值最低,在7%~9%之间(见表2)。
2.2.2 克隆分株密度
本研究发现,样方内7种竹子的1年生克隆分株数量为0~12株。1年生克隆分株数量最多的是冷箭竹,最多达12株;其次是短锥玉山竹,最多达11株;而其余竹种的1年生克隆分株数量较低。1年生克隆分株密度最大的是冷箭竹,均值达6株·m−2;其次是箬叶竹,均值达4株·m−2;密度最小的是细致箭竹,为1.3株·m−2。
样方内7种竹子的1年生以上克隆分株数量为2—103株。1年生以上克隆分株数量最多的是短锥玉山竹,最多达103株。1年生以上克隆分株密度最大的是箬叶竹,为44.8株·m−2;其次是刺黑竹,为33.4株·m−2;密度最小的是细枝箭竹,为11.1株·m−2。
样方内7种竹子的数量为3~103株。其中,密度最大的是箬叶竹,为48.8株·m−2;其次是刺黑竹,为36.6株·m−2;密度最低的是细枝箭竹,为14.5株·m−2(见表3)。
2.2.3 基径与秆高
本研究发现7种竹子中,克隆分株基径最大的是刺黑竹,均值为8.4 mm;其次是白夹竹,为7.5 mm;最小的是冷箭竹,为4.6 mm。调查到最粗壮的克隆分株是细枝箭竹,基径达15.6 mm;其次是短锥玉山竹,基径达11.9 mm。另外,克隆分株最高大的是白夹竹,均值为189.0 cm;其次是刺黑竹,为180.7 cm;最矮小的是冷箭竹,为110.9 cm。调查到最高的克隆分株是白夹竹,达297.0 cm;其次是短锥玉山竹,达288.0 cm(见表3)。
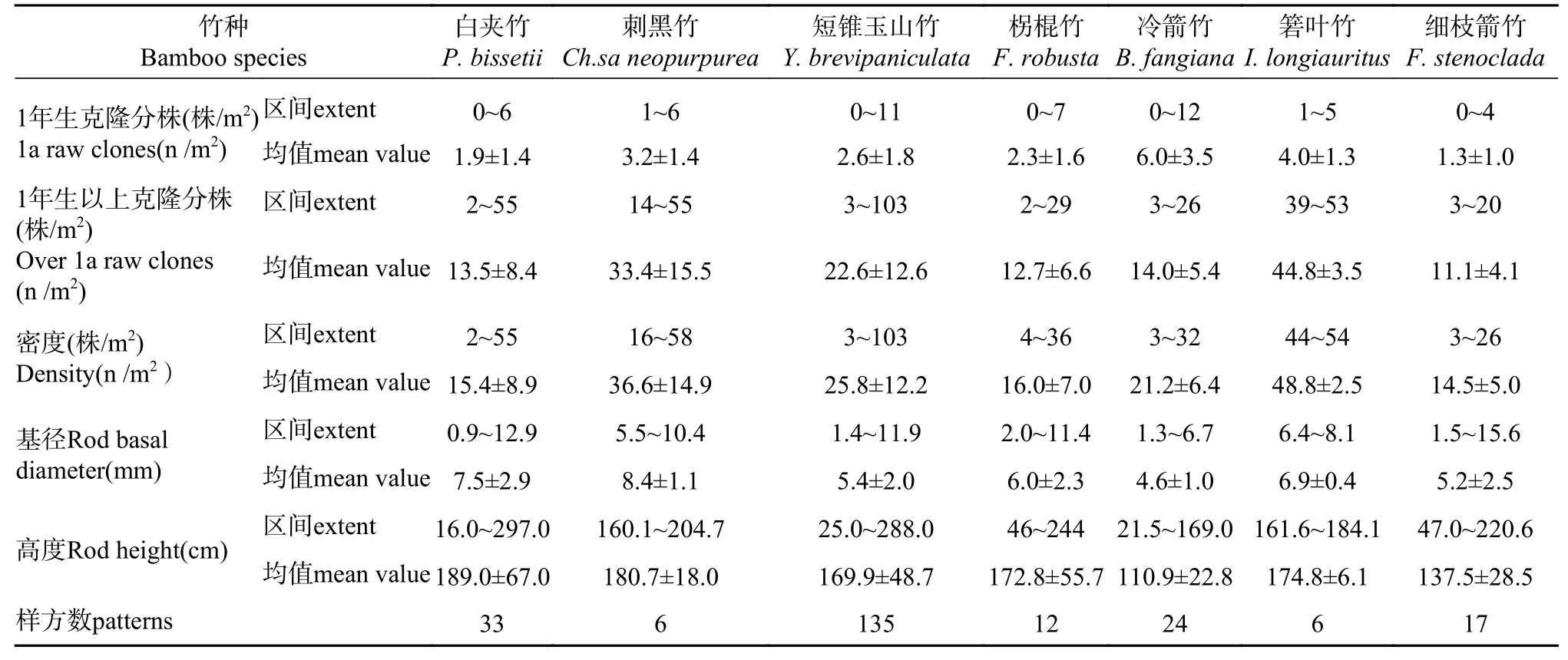
表3 七种竹子的密度、基径和高度Tab.3Density, basal diameter and height of 7 bamboo species
2.3 竹子与环境要素的关系
2.3.1 海拔的影响
统计分析发现,海拔变化仅对冷箭竹的密度有显著影响(P<0.05),而对白夹竹、短锥玉山竹以及细枝箭竹的密度均无显著影响(P>0.05),同时海拔变化对以上4种竹种的1年生克隆分株比、基径以及高度均无显著影响(冷箭竹:P>0.05,见图2;白夹竹:P>0.05,见图3;短锥玉山竹:R2<0.1,见图4;细枝箭竹:P>0.05,见图5)。
2.3.2 郁闭度的影响
统计分析发现,郁闭度变化对细枝箭竹的密度、基径和高度均有极显著影响(P<0.01),而对1年生克隆分株比无显著影响(P>0.05,见图6);郁闭度变化对冷箭竹和白夹竹的密度有显著影响(P<0.05),但对其1年生克隆分株比、基径和高度无显著影响(P>0.05;见图7,图8);而郁闭度变化对短锥玉山竹的密度、1年生克隆分株比、基径和高度均无显著影响(R2<0.1,见图9)。
3 讨论
竹类是大熊猫的食物资源,是大熊猫生存和繁衍的基础,不同区域竹子资源及种类的多样性直接影响该分布区大熊猫的食谱情况。与其他大熊猫保护区相比,如四川王朗自然保护区(5种)[23]、卧龙保护区(6种)[8,24]等,四川白水河保护区的竹子种类相对较多,高达7种,且7种竹类的不同克隆分株年龄阶段的无性系分株都有分布,各类竹种的年龄结构相对合理,除刺黑竹、箬叶竹和柺棍竹的种群数量相对较少、分布区域相对集中外,其余4种竹类资源相对丰富,能够为该区域大熊猫提供较为充足的食物资源。调查区域的7种竹类海拔分布多在3 000 m以下,竹类分布海拔略低于其他保护区。
已有研究指出,在大熊猫分布区不同大熊猫主食竹的种群数量、生长情况以及分布面积往往存在差异[4,24]。本研究取得了类似的研究结果,如研究发现该区域短锥玉山竹、白夹竹、刺黑竹增长率较高,而冷箭竹增长率相对较低;短锥玉山竹在数量和分布面积上占有绝对优势等。本研究发现,虽白水河保护区竹子种类较多,但大熊猫取食的种类较少(仅发现短锥玉山竹和细枝箭竹有取食现象),这与全国第四次大熊猫调查的结果一致[1],即同一区域,大熊猫一般仅取食少数几种竹子,这可能与大熊猫的取食偏好有关。已有研究发现不同区域大熊猫取食竹种存在差异[1,24,25],并且相同区域的大熊猫食性也会存在季节性变化[26]。
环境因子诸如温度、湿度、光照强度等对竹林密度、高度、基径、竹笋的高生长、出笋量、无性系根茎等具有显著影响[27]。而海拔因子包含了诸多环境因子,因此竹林各项指标往往会随着海拔变化而变化[27,28]。本研究表明,海拔变化对冷箭竹的密度影响显著,海拔2 400~3 200 m的冷箭竹明显高于低海拔区域情况,说明2 400~3 200 m为该区域冷箭竹生长更新的最适海拔,天然更新良好。然而本研究发现海拔对白夹竹、短锥玉山竹以及细枝箭竹的密度无显著影响,同时海拔对以上4种竹种的1年生克隆分株比、基径以及高度也无显著影响,说明以上4种竹种在海拔分布上相对均匀(冷箭竹:2 150~3 200 m;白夹竹:1 240~1 760 m;短锥玉山竹:1 540~2 800 m;细枝箭竹:1 750~2 500 m)。乔木作为森林群落的最高层,其郁闭度的大小会对林下环境(如光照强度、温度、湿度等)产生影响,从而影响竹子的生长和天然更新。本研究发现,郁闭度变化对细枝箭竹的密度、基径和高度以及对冷箭竹和白夹竹的密度影响显著,说明不同乔木郁闭度下竹子生长和更新往往存在差异。这与李波等对岷山北部箭竹的研究结果[29]以及贾炅和武吉华[7]对王朗自然保护区缺苞箭竹的研究结果类似。已有研究指出,郁闭度过高的过分荫蔽环境会导致竹子缺乏光照而影响根系的发育,使竹子幼苗的根系生长脆弱,根系竞争力差,进而影响幼苗对水分和养分的吸收,造成幼苗死亡[30]。而在郁闭度很低的开阔地段,因为林下地面的光照充足,这些区域林下往往会灌木杂草丛生,从而不利于竹子的生长和天然更新[7,31]。
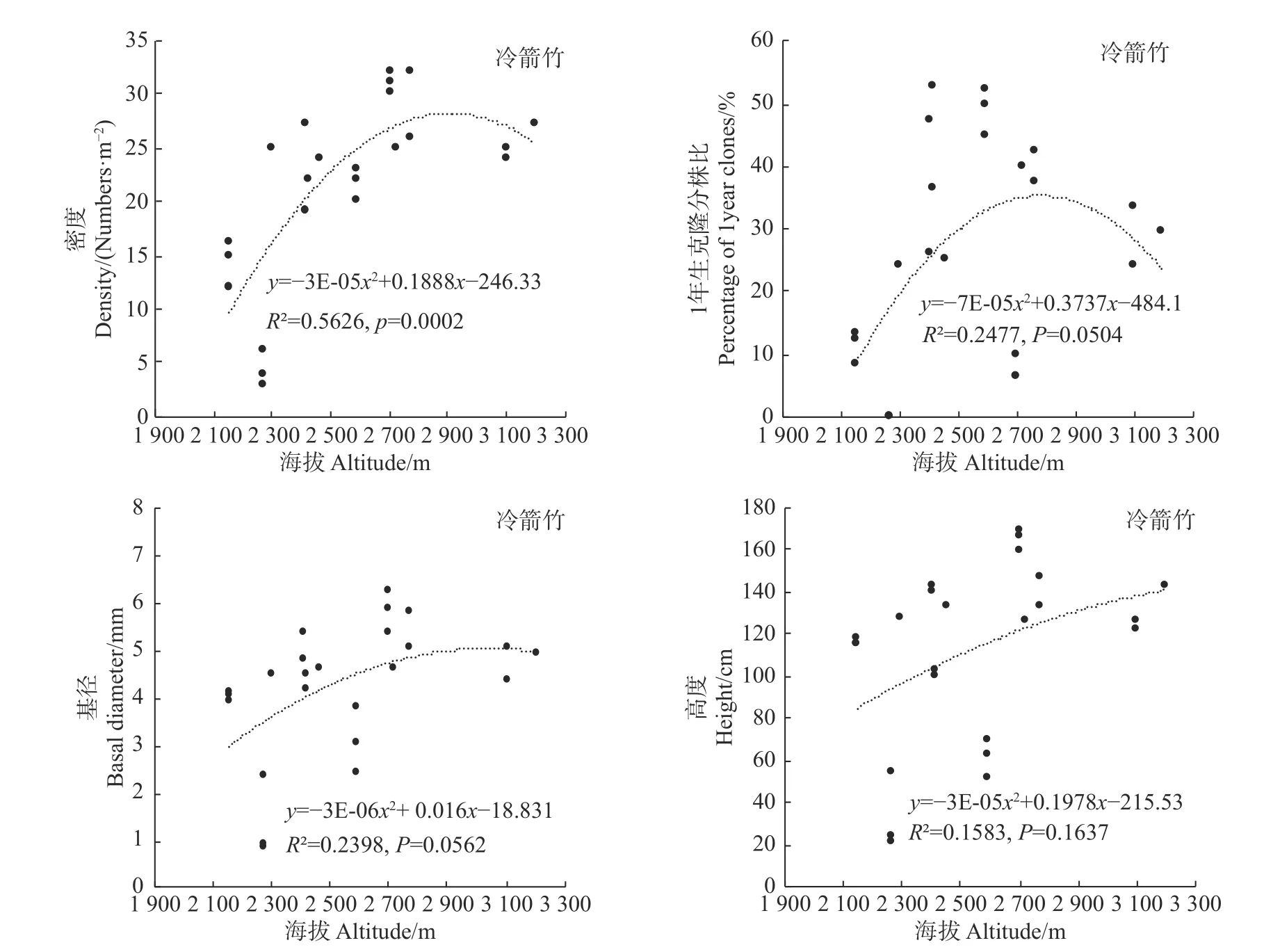
图2 海拔对冷箭竹的影响Fig.2Effects of altitude on Bashania fangiana
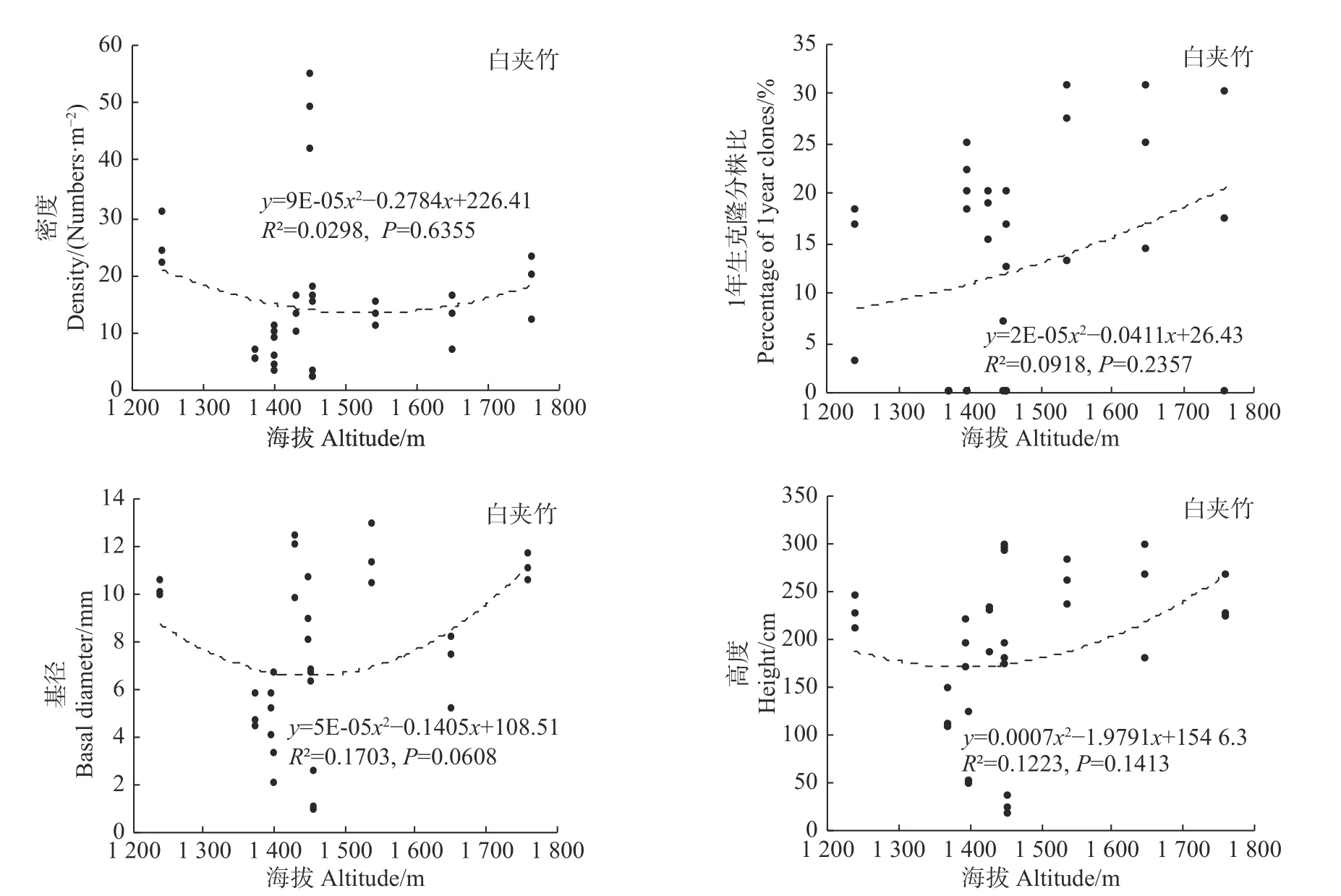
图3 海拔对白夹竹的影响Fig.3Effects of altitude on Phyllostachys bissetii
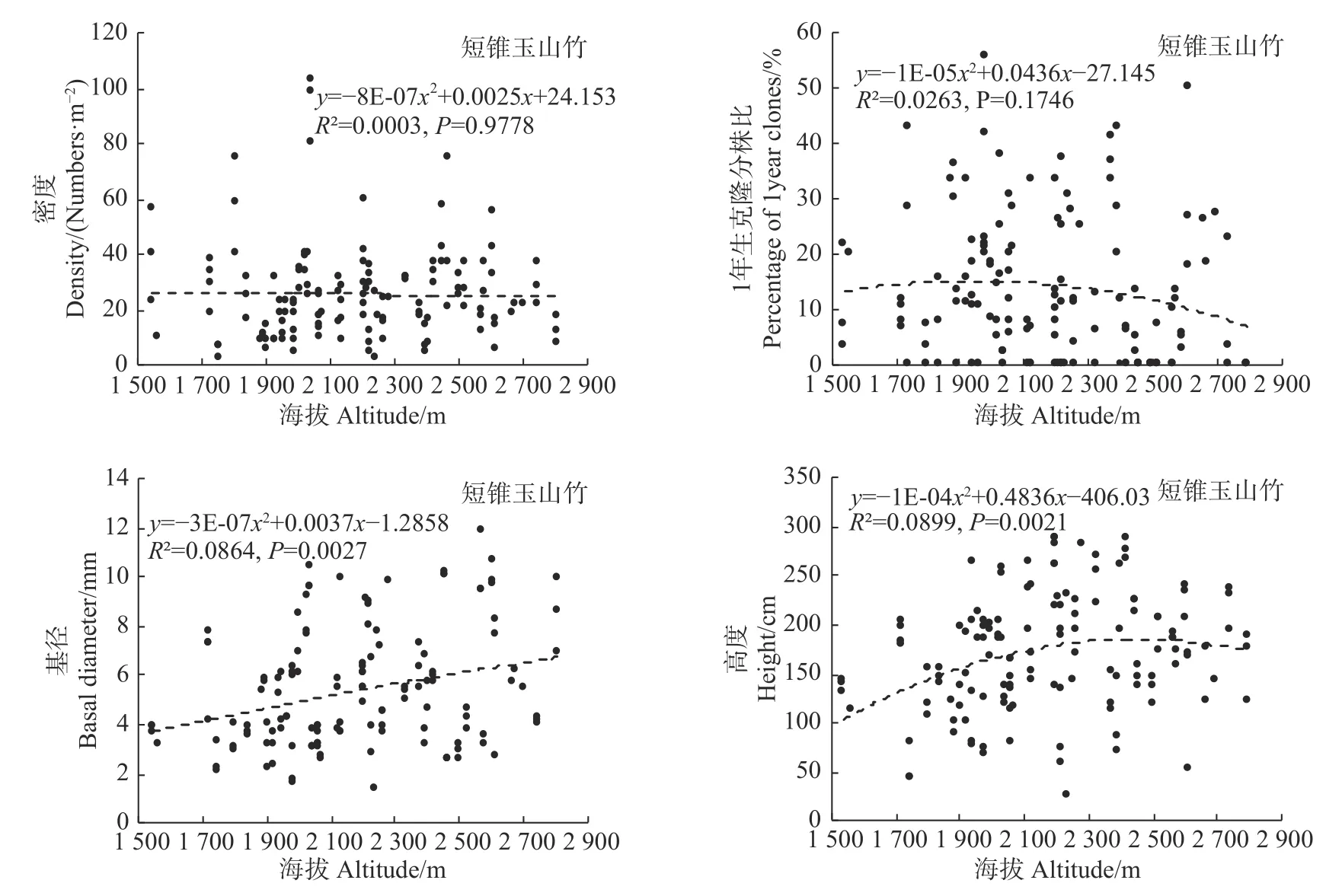
图4 海拔对短锥玉山竹的影响Fig.4Effects of altitude on Yushania brevipaniculata
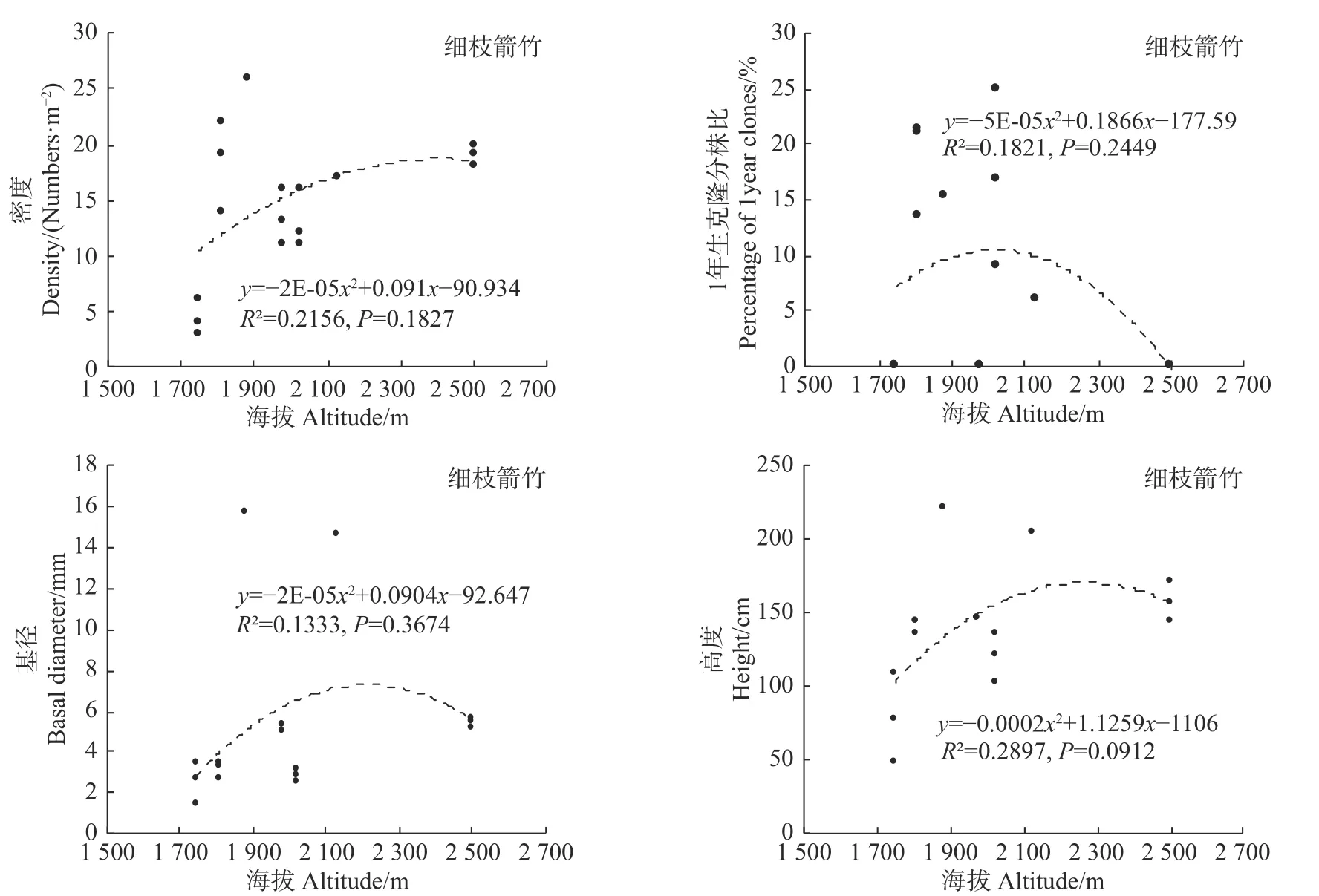
图5 海拔对细枝箭竹的影响Fig.5Effects of altitude on Fargesia stenoclada
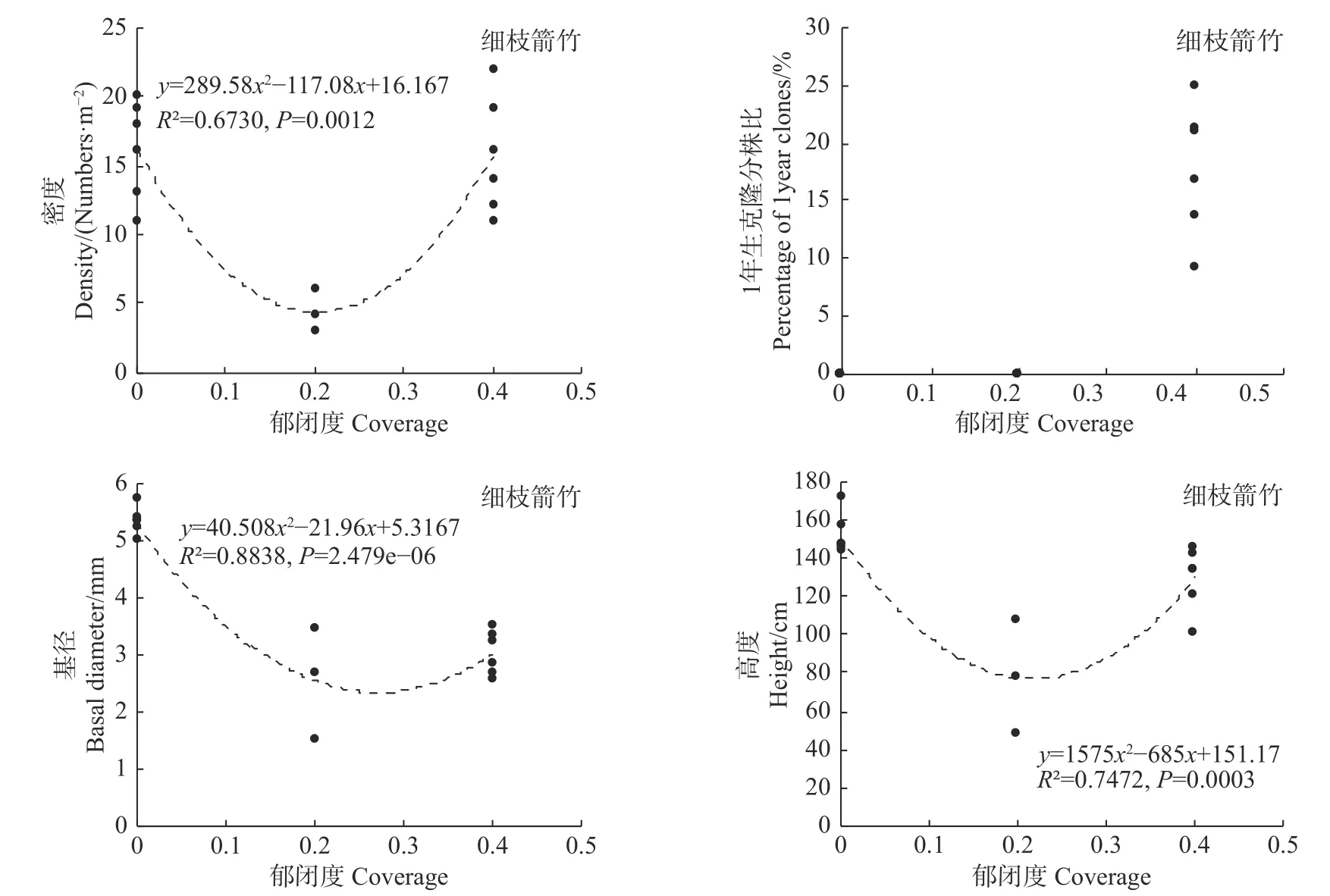
图6 郁闭度对细枝箭竹的影响Fig.6Effects of canopy density on Fargesia stenoclada
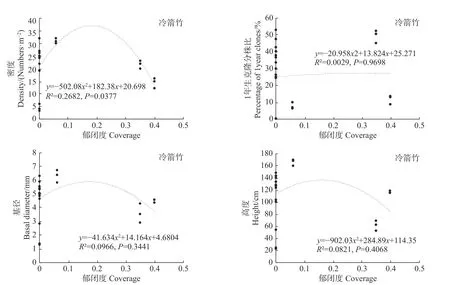
图7 郁闭度对冷箭竹的影响Fig.7Effects of canopy density on Bashania fangiana
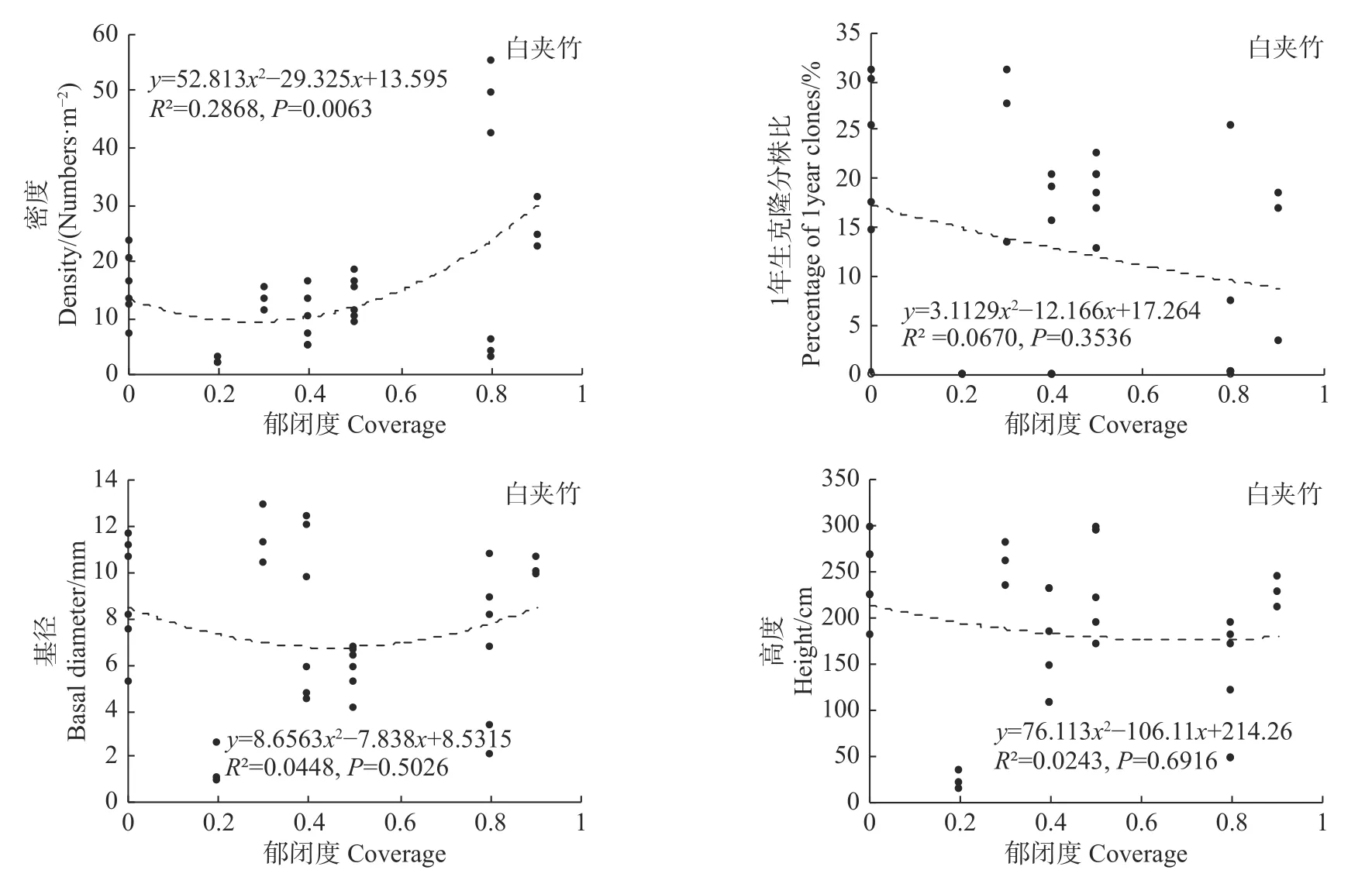
图8 郁闭度对白夹竹的影响Fig.8Effects of canopy density on Phyllostachys bissetii
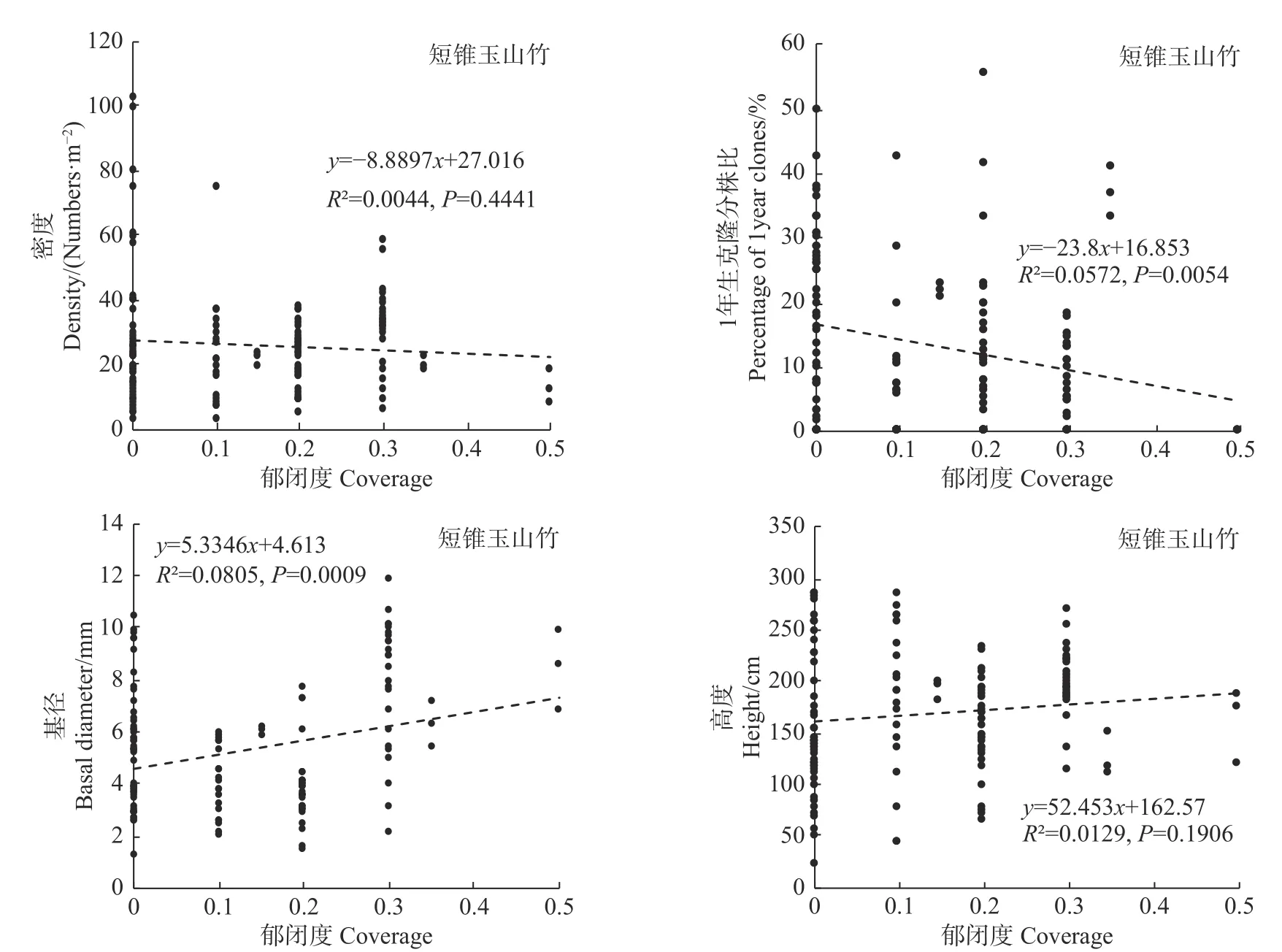
图9 郁闭度对短锥玉山竹的影响Fig.9Effects of canopy density on Yushania brevipaniculata
已有研究发现,相同竹种的植株基径和株高存在显著的正相关性[7]。本研究发现短锥玉山竹基径-株高变化系数较高,说明在相同粗壮程度下,短锥玉山竹能长更高以获得更多阳光,进而易于在取食竹群落中不断积累优势,成为优势竹种。
本研究发现,尽管全光照下细枝箭竹有较高的密度、基径和高度,但由于全光照不利于大熊猫的活动,因而在管理中建议倾向于维持在0.4左右的郁闭度为宜。另外,研究发现该区域大部分竹类偏好郁闭度较低的生境,因此在管理中可考虑对茂密森林(尤其是栖息地内的人工纯林)进行适当透光疏伐或以林窗形式疏伐而形成林窗,使栖息地的乔木层平均郁闭度保持在一个相对较为适宜的范围内(以0.4~0.7为宜)。
竹子具有周期性开花死亡的特点,大熊猫在较为单一竹种组成的生境中,为避免将来可能会面临由竹类周期性开花死亡造成的食物相对短缺问题,可以适当进行不同竹种的镶嵌种植以提高竹类的多样性,同时也可以种植不同竹鞭年龄的竹子以形成异龄竹林。此外,在实际管理中还应该与科研单位建立合作关系,探索大熊猫取食较多的短锥玉山竹管理模式。
猜你喜欢
宁夏大学学报(自然科学版)(2022年2期)2022-07-19
新农村(浙江)(2022年3期)2022-03-22
小哥白尼(野生动物)(2021年7期)2021-11-20
花卉(2021年9期)2021-05-15
热带作物学报(2020年6期)2020-08-04
福建林业(2020年5期)2020-03-18
幼儿教育·父母孩子版(2020年8期)2020-03-04
家教世界·创新阅读(2019年5期)2019-06-14
热带林业(2019年4期)2019-03-05
学苑创造·A版(2018年7期)2018-08-07
