
秋季长江口及邻近海域一氧化碳的分布、通量与微生物消耗速率❋
2020-11-04 08:03陈佳玉张洪海李江萍杨桂朋
中国海洋大学学报(自然科学版) 2020年12期
陈佳玉, 张洪海,2, 李江萍❋❋, 杨桂朋,2
(1.中国海洋大学化学化工学院,山东 青岛266100; 2.青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室, 山东 青岛 266237)
海洋是大气中CO的来源之一[1-3],其中海洋每年向大气释放CO中C含量的范围在3.7~600 Tg之间[2-6],是全球碳循环的重要组成部分。CO经海洋释放到大气中,随后与对流层中羟基自由基反应氧化为CO2[7-8],由于对流层中羟基自由基被大量消耗导致对流层的氧化能力减弱,使得大气中其它活性气体的浓度受到影响,因此CO被称为间接温室气体。CO作为大气中一种重要的痕量气体,无论是在全球变暖还是大气化学的研究中都有着十分重要的地位。
溶解有色有机物(Colored dissolved organic mater,CDOM)光化学生产过程和溶解有机物(Dissolved organic matter,DOM)热力学降解过程是海洋中CO的两大来源,其中CDOM的光化学生产为主要途径[2,4,5,9]。微生物消耗和海-气扩散是海水中CO去除的主要途径,其中被微生物消耗CO的含量占海水中CO总消耗量的86%[9]。
近年来,中国对CO的研究主要集中在CO的分布、海-气通量和循环过程中,调查区域以黄渤海和东海海域为主,对河口海域的研究较少。长江口及邻近海域位于我国黄海和东海交界处,属于极其典型的陆架型河口,作为河流入海处,该海域主要受到长江冲淡水的影响,同时苏中沿岸流、江浙沿岸流以及台湾暖流也对长江口及邻近海域有一定影响。此外,黄海冷水团和黑潮水也对长江口及邻近海域东部也存在一定的影响[10-11]。多种因素使得长江口及邻近海域的生态环境变得极其复杂,与周边海域大不相同。由于人类活动的增加,陆源对近海海域生态环境的影响越来越大,使得陆架海域、近岸海域以及河口海域中的CO与开阔大洋中的CO在海-气通量、生产与消耗等方面已经产生很大的不同,河口、海湾和陆架等近岸海域面积较小,在陆源输入的直接影响下,海水中CO浓度较高,进而通过海-气扩散过程向空气中排放大量的CO[9, 12-15]。本研究于2019年秋季对我国典型的河口海域—长江口及邻近海域进行了大规模调查和现场培养实验,对长江口及邻近海域中CO的分布、通量和消耗过程进行详细探讨。此项调查有助于丰富我国在河口海域CO分布与循环研究方面的内容,加深海洋对大气CO释放贡献的认识。
1 材料与方法
1.1 样品采集
于2019年10月17~26日随“浙渔科2号”对长江口及邻近海域进行现场调查研究,图1为调查区域采样站位图,共包含7个断面的57个调查站位。现场采用12 L Niskin Rosette采水器采集,共采集57份调查站位的表层海水与大气样品,同时选取A4和A6断面作为重点研究断面,并采集不同深度的海水样品,海水样品在无气泡的情况下通过硅胶管注入50 mL的样品瓶(预先经10%HCl-Milli-Q水清洗)中,样品瓶在使用前需要用海水润洗2~3次,然后将样品瓶密封,瓶中无气泡。大气样品于船走航时,用气密性良好的10 mL注射器在甲板上迎风采集,以减少船体造成的污染。大气样品与海水样品均现场测定。
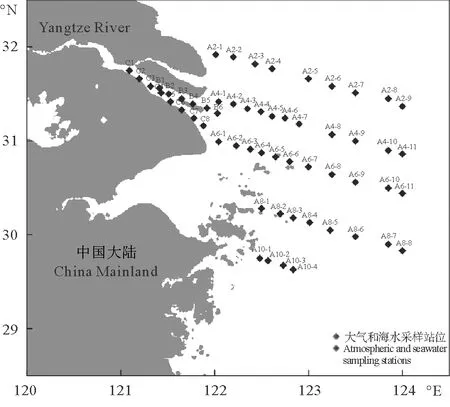
图1 秋季长江口及邻近海域大气和海水采样站位Fig.1 Atmospheric and seawater sampling stations in the Changjiang River Estuary and its adjacent area in autumn
1.2 样品分析
1.2.1 CO浓度的测定 通过顶空平衡分析法来现场测定海水和大气样品中CO的含量[16],该方法操作步骤为:通过硅胶管将现场采集的样品注入到50 mL的玻璃瓶中,保证瓶中无气泡,密封,然后通过10 mL气密性良好的玻璃注射器(VICI)注入6 mL的CO零空气,CO零空气由氮气纯化仪(先普半导体技术有限公司)给出,最后将样品瓶置于轨道式振荡器(江苏盛蓝仪器制造有限公司)上并以290 r·min-1的速度进行振荡来快速达到平衡状态,振荡时间为5 min[16],振荡结束后,样品达到气液平衡状态,用10 mL气密性良好的玻璃注射器抽取顶空气体,通过预先放置阻水滤膜(0.2 μm Nuclepore Teflon filter,φ 13 mm)的进样口注入到Ta3000痕量气体分析仪(美国Ametek公司)中进行测定。大气样品迎风采集后直接注入到仪器中进行测定。整个实验过程中,采用CO标准气体(99.6 ppbv,大连大特气体有限公司)进行单点校正。该测量方法测量的误差较低,灵敏度较高,测量数值的相对标准偏差<4.4%,最低检出限为0.02 nmol·L-1[16]。
1.2.2 CO微生物消耗速率的测定 CO的微生物消耗速率通过黑暗培养法[17-18]来测定,此方法的优点是消除了CO光化学生产的影响,具体操作步骤为:表层海水通过300 mL玻璃注射器(预先经10%HCl-Milli-Q水清洗,并用避光胶带封闭)从采水器中采集,样品采集前应先用表层海水对注射器进行润洗,重复2~3次,在整个采集过程中,注射器内不应留有气泡。样品采集完成后,立即使用三通阀对注射器进行密封,然后将注射器置于表层海水循环水浴箱中进行现场避光培养。通过连接有三通阀的塑料管将样品注入到50 mL样品瓶内,瓶内无气泡,密封,立即进行CO浓度测定,样品转移过程中要防止大气进入注射器内。在样品采集后,立刻测定CO浓度来作为CO的起始浓度,30 min后再次取培养样品测定CO浓度,作为第二个时间点的浓度,然后每隔30 min到1 h(根据测定消耗速率的情况决定)测量一次样品中CO的浓度,培养实验共选取5个时间点进行测定。表1为CO微生物消耗速率的测定站位。
1.3 CO海-气通量和饱和系数的测定
CO的海-气交换通量可以通过下式进行计算[19]:
F=k([CO]surf-[CO]eq)。
(1)
式中:F为CO的海-气通量,nmol·(m2·h)-1; [CO]surf和[CO]eq分别为CO在表层海水中的浓度和达到气液平衡后海水样品中的浓度,nmol·L-1;k为气体交换常数,m·h-1,与风速和气体施密特常数Sc(Schmidt number)有关,k可由经验公式计算得出,本文采用E2011经验公式,该经验公式适用范围广(0~18 m·s-1),且考虑到风速为0 m·s-1时,气泡破裂产生海-气扩散的贡献[20]。其中Sc的值可以通过下式得出[21]:
Sc=-0.055 3t3+4.382 5t2-140.07t+2 134。
(2)
式中:t为样品海水的温度,℃。
[CO]surf可以由达到气液平衡后顶空气体中CO的含量来确定[22]:
[CO]surf=pma(βpVw+Va)/(RTVw)。
(3)
式中:P为现场大气压力,atm;ma为样品达到气液平衡后顶空气体中CO的体积分数;β(Busen solubility coefficient)为CO在海水中的溶解度常数,海水的温度和盐度为β的主要影响因素[23];Vw为采取的海水样品体积,mL;Va为注入样品瓶中顶空气体的体积,mL;R为气体常数,0.082 06 atm·L·(mol·K)-1;T为现场海水的温度,K。
[CO]eq可以通过下式进行计算得出[24]:
[CO]eq=([CO]atm×β)/MCO。
(4)
式中:[CO]atm是现场测得的大气样品中CO的体积分数;MCO为CO的摩尔体积25.094 1 L·mol-1(标准压力和温度)[25-26]。
海水中CO的过饱和系数α可通过下式得到[24]:
(5)
1.4 叶绿素a(Chl-a)浓度的测定
从Niskin Rosette采水器中采取300 mL样品,采用玻璃纤维滤膜(Whatman GF/F,φ 47 mm,预先经450 ℃高温灼烧)进行过滤,过滤后将滤膜对折装入用锡纸包裹好的15 mL离心管中,冷冻带回陆地用于Chl-a浓度的测定。在Chl-a浓度的测定前,提前在装有滤膜的离心管中加入90%(体积分数)丙酮水溶液,随后在避光处进行萃取,时间为24 h。萃取完成后在转速为4 000 r·min-1条件下离心10 min后吸取上清液,通过F-4500荧光仪(日本日立)测定Chl-a的浓度,该方法的最低检出限为0.01 μg·L-1[27]。
2 结果与讨论
2.1 大气中CO的浓度分布
秋季长江口及邻近海域大气中CO的体积分数处在142.6~482.7 ppbv之间,平均值为(271.6±80.1) ppbv,其中最大值为最小值的3.4倍。大气中CO的最大值出现在长江口口门附近(A2-1站位);最小值出现在长江口外海海域(A8-7站位),但仍高于开阔大洋[28]。大气中CO的体积分数整体上呈现出近岸高,远岸低的趋势(见图2),近岸海域上空大气受陆源影响严重,大气中CO体积分数较高,同时采样时风向也会对结果造成一定影响。该航次调查结果高于春季东海和黄海大气CO浓度的平均值(190 ppbv)[12]以及夏季东海和南黄海CO浓度平均值(117 ppbv)[29],秋冬季节北方燃烧暖气向近岸海域上空排放大量CO,导致河口等近岸海域上空大气中CO含量明显增高,本次调查结果也与秋季黄东海调查结果接近(297 ppbv),陆源输入是影响河口海域大气中CO的一个重要因素[13-14]。
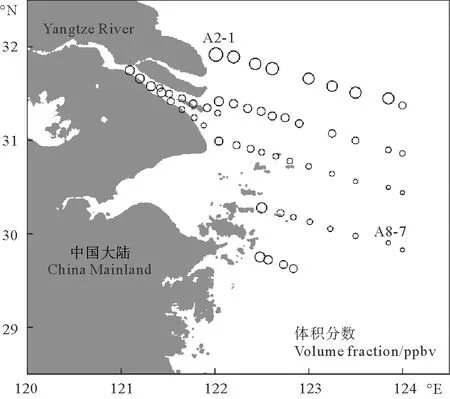
图2 秋季长江口及邻近海域大气中CO的水平分布
2.2 海水中CO的分布特征及其余环境因子的关系
2.2.1 海水中CO的水平分布 长江口及邻近海域各个站位表层海水中温度、盐度以及Chl-a浓度以及CO浓度的水平分布如图3所示,表层海水中CO浓度处在0.11~3.27 nmol·L-1之间,平均值为(0.71±0.72)nmol·L-1。秋季长江口表层海水温度范围为21.15~24.15 ℃,平均值为(22.71±0.83)℃,温度变化趋势与表层海水中CO浓度变化趋势相近,在温度高的海域,表层海水中CO浓度较低。盐度主要通过影响CO的微生物消耗过程来影响表层海水中CO的浓度,秋季长江口及邻近海域盐度范围为0.16~33.5,平均值为(22.88±13.19),调查结果显示长江口及邻近海域盐度变化十分明显,位于长江入海口附近的B、C两断面盐度接近0。在盐度较低的长江口口门以及口内海域,微生物消耗速率较低(下文会进行详细探究),长江口口门附近大气中CO浓度也较高,结果导致长江口口内以及口门附近海域表层海水中CO浓度较高。秋季长江口及邻近海域内表层海水中Chl-a浓度变化范围为0.52~5.87 μg·L-1,平均值为(1.17±1.37)μg·L-1。Chl-a浓度最大值出现在A8-7站位附近,由长江入海径流逐步形成的苏中沿岸流和台湾暖流在该区域附近形成上升流,浮游植物从上升流中获取营养盐,同时浊度较小,光照充足,浮游植物大量繁殖,初级生产力高[30-31],使得Chl-a浓度升高出现最大值。Chl-a浓度最小值出现在A4-2站位,该站位位于长江口口门附近,水深较浅,由于长江冲淡水带来的大量泥沙使得海水浊度高,光照稀少[32],对浮游植物的生长造成限制,影响初级生产力,导致Chl-a含量较低,结果表明Chl-a浓度分布趋势与CO浓度无明显关系,可以看出CO主要由CDOM光降解产生,与生物活动关系较小。本次调查研究结果与秋季东海和南黄海调查结果[14]相近,但低于东海和南黄海春季和夏季分别为2.24和2.49 nmol·L-1的调查结果[13, 29],这主要是由于秋季光照时间较短,光照强度较弱,从而导致CO的光化学生产过程减弱。[CO]surf的最大值和最小值分别出现在C3站位和A8-3站位,取样时间分别为上午和凌晨。[CO]surf分布整体上呈现出近岸高,远岸低的趋势,这主要是由于我国长江口及邻近海域人为活动为该海域生态环境提供大量营养盐和有机物,而CDOM的光降解为海洋中CO的主要来源,该现象导致近岸海水中[CO]surf明显高于远海。同时A2-5和A6-9站位附近[CO]surf相对较高,此两处高值区域均处于长江冲淡水的羽状锋区域[33]。大量营养盐随着长江冲淡水在羽状锋区域聚集,并且该区域水体泥沙量较小、透明度大、水质清澈,使得光照强度高,有利于CDOM光降解产生CO。因此,陆源输入为影响近岸海水中CO浓度分布的一个重要因素。本次调查期间白天[CO]surf平均值为0.85 nmol·L-1,夜晚[CO]surf平均值为0.30 nmol·L-1。白天调查站位[CO]surf平均值明显高于夜间调查站位[CO]surf,整体来看[CO]surf表现为中午前后浓度高,凌晨前后浓度低。造成这种现象的原因主要由于正午附近光照强度较高,CO光化学生产过程加强,CO浓度较高,可以看出表层海水中CO浓度有较为明显的周日差异。不同时间段光照强度不同,从而使CO浓度值产生较大程度的变化。太阳光的光照强度也是影响表层海水中CO浓度的一个重要因素。
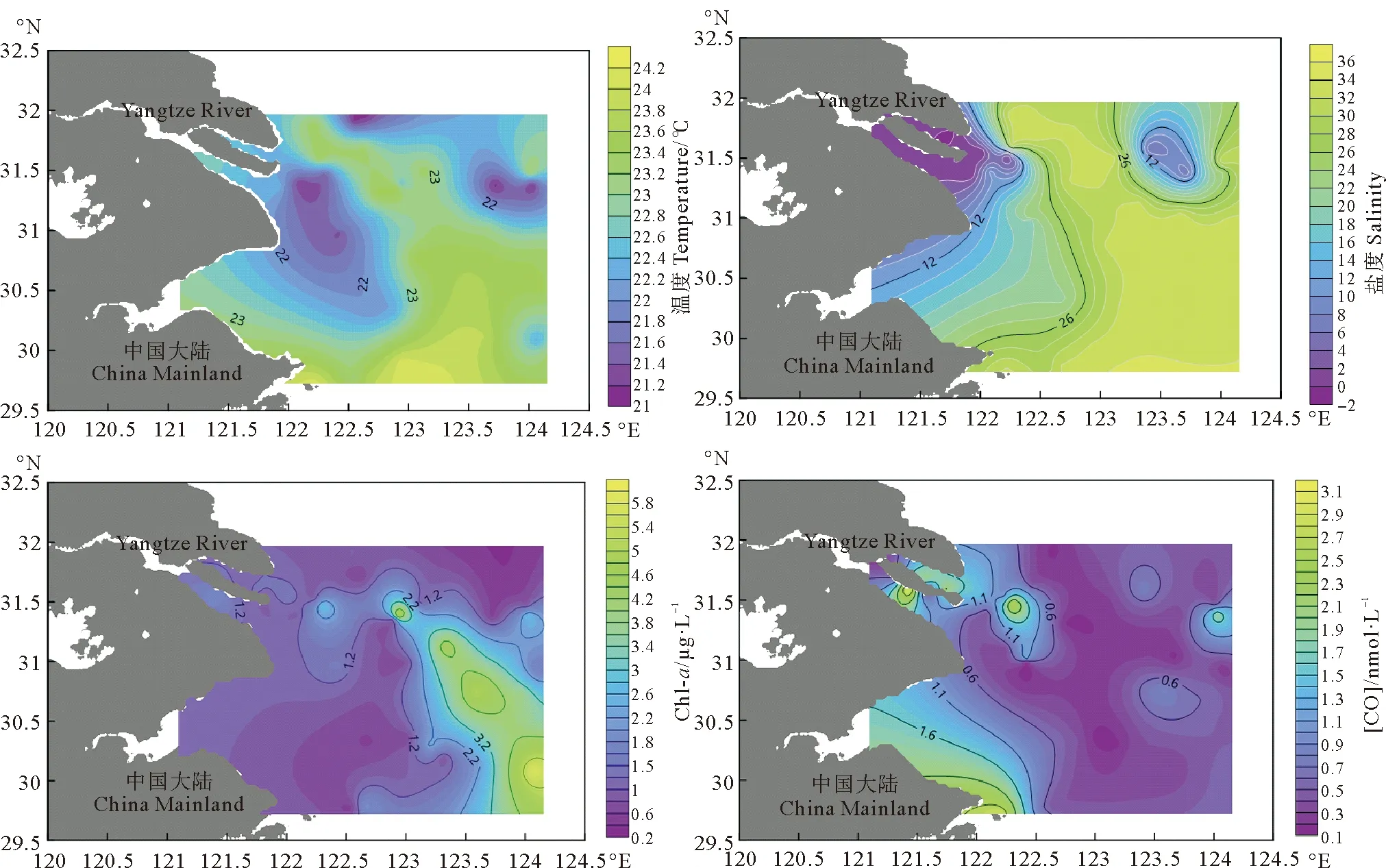
图3 秋季长江口及邻近海域表层海水温度、盐度、Chl-a和CO的水平分布
2.2.2 海水中CO的垂直分布 调查海域部分站位CO浓度的垂直分布如图4所示,表层海水中CO的浓度最高,并且随着深度增加浓度逐渐降低。该现象主要是因为表层海水受阳光照射强烈,大量CDOM在阳光照射下降解产生CO,由于海水及海水中的颗粒物会不断对太阳光进行吸收和散射,所以随着深度增加,光照强度也会逐渐减弱,导致CDOM光降解速度下降,使得底层海水中CO浓度小于表层海水,呈现出递减趋势[24],与春季和夏季东海研究结果相符。随着海水深度增加,盐度有增高趋势,与CO浓度变化趋势相反,主要是由于盐度变化会对CO微生物消耗过程造成影响。Chl-a的垂直分布与海水中CO浓度无明显关系,该结果与文献报道一致[17, 34]。
A4和A6两个受长江冲淡水影响较大的断面CO浓度、温度和盐度的垂直分布如图5所示,A4和A6断面处于典型的长江冲淡水区域,具有明显的盐度梯度,从西向东盐度逐渐增加。调查结果显示长江口口门及口门内海域CO浓度明显高于外海,主要由于长江口口门及口门内海域由长江冲淡水带来的大量CDOM光降解产生CO。在两个断面中均可以观察到CO浓度随海水深度增加而降低,部分站位如A4-3的CO浓度最大值出现在中层水体(10 m左右),这种情况的原因可能是风速过大导致海面波破碎造成表层CO通过海气界面扩散到大气中,造成表层海水中CO浓度低于深层水体中CO浓度。由于底层水体受阳光辐射较小,CO光化学产生过程较弱,从而导致CO浓度较低。温度与CO变化趋势相近,也呈现出随深度增加而降低的趋势。
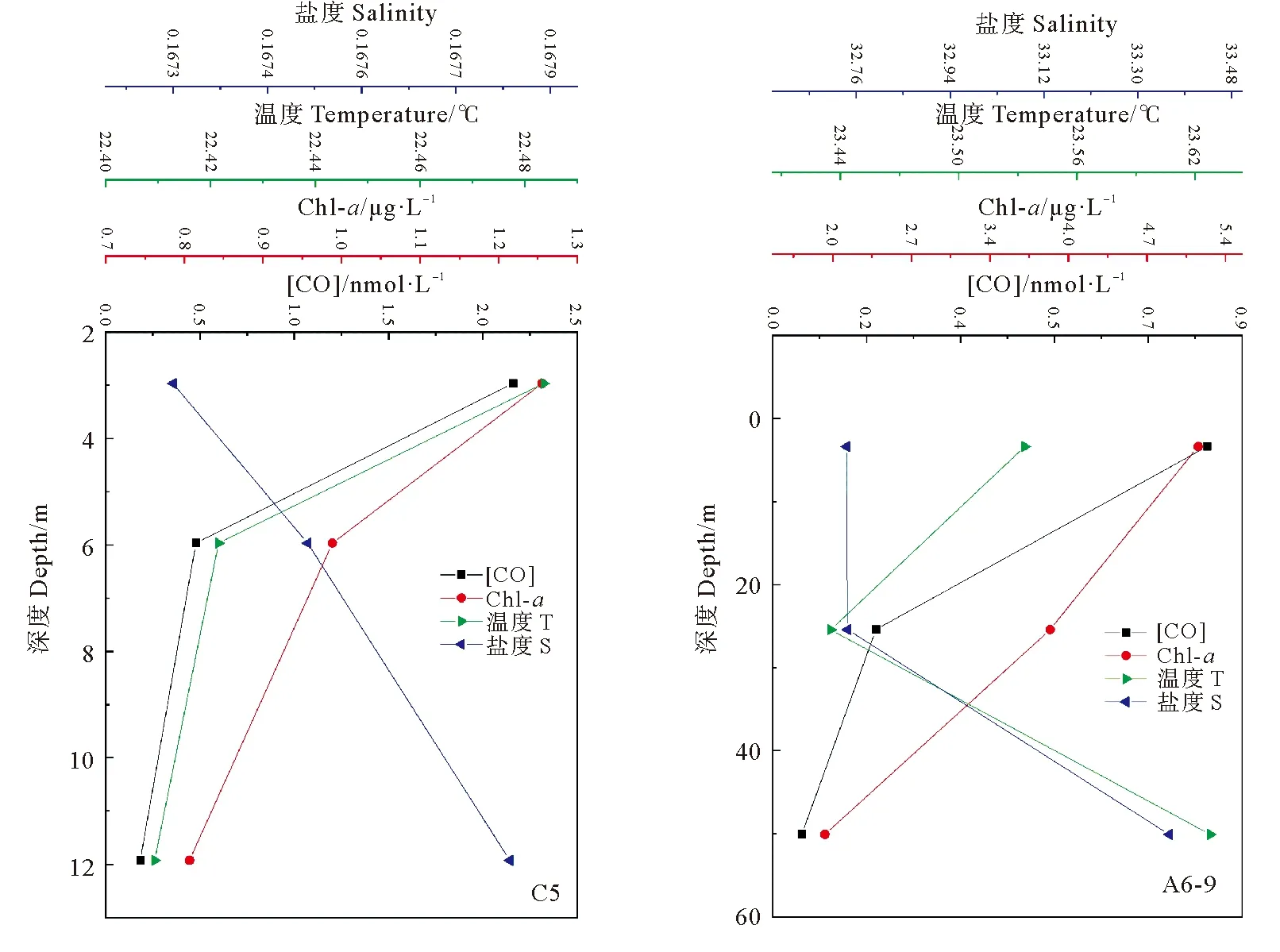
图4 C5和A6-9站位温度、盐度、Chl-a和CO浓度的垂直分布
2.3 CO微生物消耗
本航次通过现场黑暗培养法对部分重点站位CO的微生物消耗速率进行培养实验测定。结果表明,CO浓度的变化趋势随着时间的增加呈现出指数型降低,该趋势符合动力学一级反应的特征,CO微生物消耗系数(R2)、消耗速率常数(kbio)、周转时间(τ=1/kbio)、Chl-a浓度以及盐度如表1所示,kbio处在0.21~1.12 h-1之间,平均值为(0.62±0.31)h-1,调查结果高于秋季东海调查结果(0.17 h-1)[36],该现象主要是由于长江口及邻近海域受人为活动影响日益严重,致使该海域水体生态环境状况十分复杂,大量营养盐、有机物的输入严重影响微生物的种类以及微生物的丰度,进而影响kbio的数值。
盐度作为影响kbio的重要因素,本航次着重从受长江冲淡水影响严重的海域进行培养站位的选取,表层海水盐度处于0.21~33.50之间,以便于探究长江口及邻近海域CO微生物消耗速率与海水盐度的关系。调查结果表明当盐度大于19时,在A6-2站位kbio出现最大值(1.12 h-1),此处盐度较低;在A8-8站位kbio出现最小值(0.54 h-1),可以看出A8-8站位盐度值较高。而盐度低于19时,kbio在低盐站位C6出现最小值(0.21 h-1),在C2站位出现最大值(0.40 h-1)。通过数据处理得到,盐度大于19时,kbio与盐度之间呈现出随着盐度增加kbio逐渐降低的趋势,二者具有较好的相关性(R2=0.826 4,n=4,P<0.01)。同时在盐度低于19的海域kbio明显低于盐度大于19的海域,文献表明在盐度为0~19时,kbio随盐度升高而升高;在盐度大于19时,kbio随盐度升高而降低[14,24,38],与本次调查结果一致。
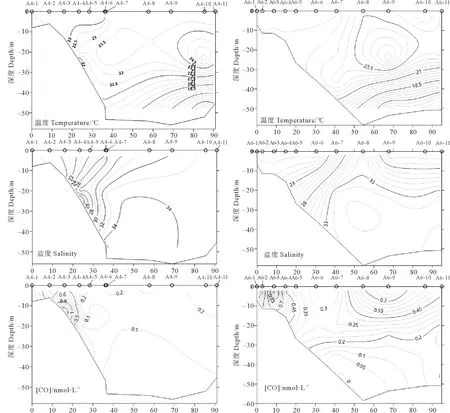
图5 A4和A6断面温度、盐度和CO浓度的垂直分布
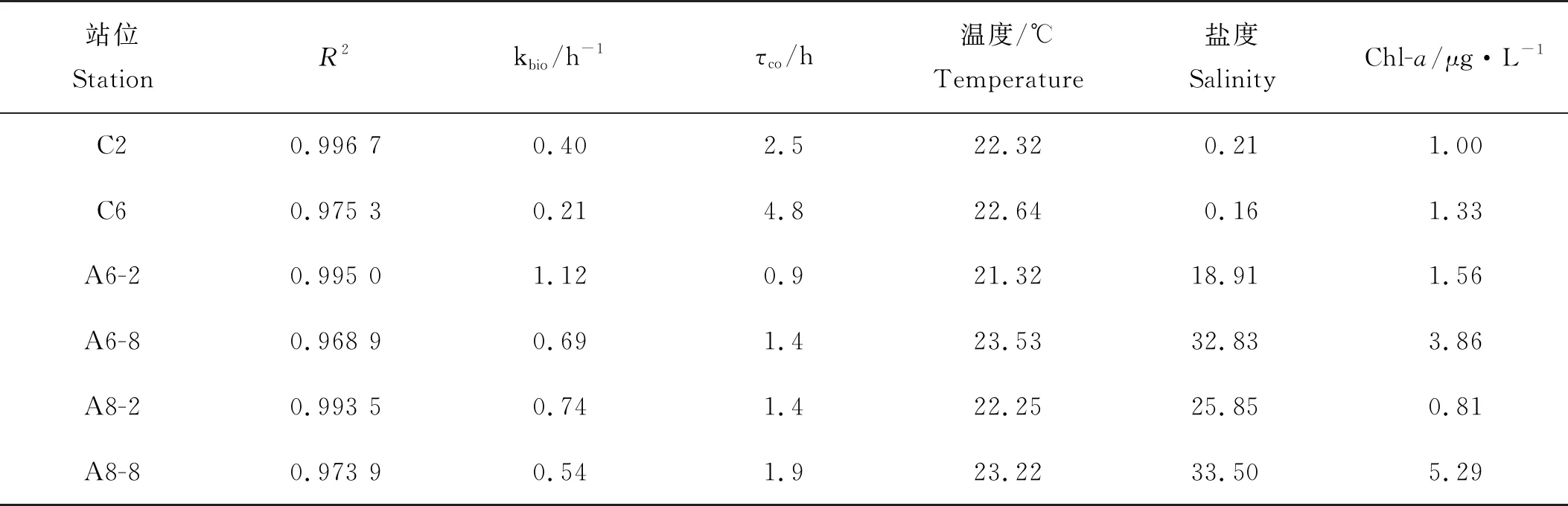
站位StationR2kbio/h-1τco/h温度/℃Temperature盐度SalinityChl-a/μg·L-1C20.996 70.402.522.320.211.00C60.975 30.214.822.64 0.161.33 A6-20.995 01.120.921.3218.911.56A6-80.968 90.691.423.53 32.833.86 A8-20.993 50.741.422.25 25.850.81 A8-80.973 90.541.923.2233.505.29
通过研究发现CO的微生物消耗过程较为复杂,影响因素多样化, Chl-a的浓度,微生物种类和丰度,微生物繁殖所需的有机物、营养盐、温度、盐度和pH等因素均会对CO的微生物消耗过程造成影响[39-41],其中许多研究还需要补充。
2.4 CO的海-气通量
长江口及邻近海域部分站位CO的过饱和系数处在0.54~14.51之间,平均值为(3.25±3.02),从数值上可以看出,海水中CO的浓度相对于大气中CO的含量绝大部分都是过饱和的,可以说明长江口及邻近海域是大气中CO的源。调查结果低于春季和夏季的调查结果,和秋季黄东海调查结果相近,春季、夏季和秋季黄东海CO过饱和系数平均值分别为16.00、29.36和3.34[13-14,29]。过饱和系数变化趋势与表层海水中CO浓度的趋势大体一致,本次调查结果显示秋季表层海水中CO的浓度低于春季和夏季表层海水中CO的浓度,与CO过饱和系数变化趋势相同,可见表层海水中CO的浓度为其主要影响因素,同时秋季温度较低对海水中CO溶解度也会造成一些影响,进而影响CO的过饱和系数。
部分调查站位CO的海-气通量与风速及过饱和系数的变化趋势如图6所示,CO的海-气通量范围处在-2.1~31.15 nmol·m-2·h-1之间,平均值为(4.84±7.54) nmol·m-2·h-1,其中最大值出现在C3站位,C3站位的风速和过饱和系数分别为5.8 m·s-1和14.51,此站位风速不是最大,但[CO]surf(3.27 nmol·L-1)较高,二者共同作用导致CO海-气通量出现最大值;最小值出现在A2-2站位,A2-2站位的风速和过饱和系数分别为6.7 m·s-1和0.54,该站位[CO]surf(0.18 nmol·L-1)较小。其中海-气通量的变化趋势还与风速的变化趋势相近,可见风速大小和海水中CO的过饱和系数以及[CO]surf为影响CO海-气通量的主要因素。在Sargasso海域以及Beaufort海域春季的调查结果分别为6.54 μmol·m-2·d-1和(6.20±6.43)μmol·m-2·d-1[21,42],春季黄东海的调查结果为6.67 μmol·m-2·d-1[13],夏季黄东海的调查结果为12.73 μmol·m-2·d-1[29],通过本次调查得到的数据与不同季节数据进行对比可以看出秋季调查结果低于夏季和春季调查结果,主要是春夏季节海水中CO的过饱和程度和风速均处于较高水平。为了加深长江口及邻近海域对全球碳循环贡献的了解,通过本航次调查得到的CO海-气通量对长江口及邻近海域(面积约为73 000 km2)的全年CO碳排放进行了估算,结果表明长江口及邻近海域每年排放约3. 0×106g C进入大气。尽管河口海域面积仅占全球海洋面积的一小部分,但是随着人类活动的增加以及陆源输入带来大量营养盐以及有机物使得河口区域生态环境十分复杂,有可能贡献出更高的CO通量。
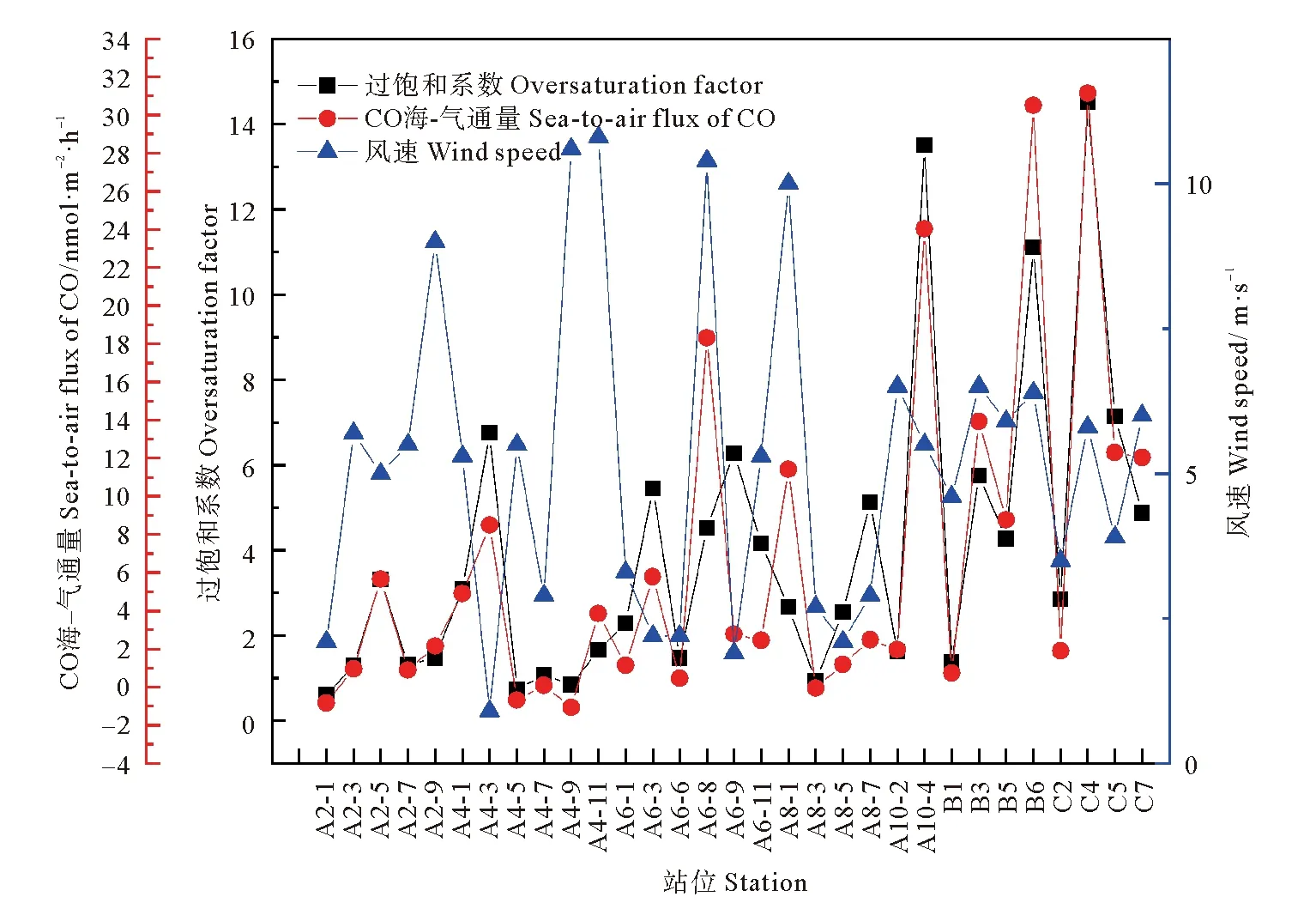
图6 CO海-气通量、风速以及过饱和系数的变化
3 结论
(1)秋季长江口及邻近海域大气和海水中CO均呈现出近岸高、远岸低的趋势,并且含量明显高于开阔大洋。此外,表层海水中CO浓度最高,随着深度增加CO浓度逐渐降低。
(2)表层海水中CO浓度呈现明显的周日差异,在光照强度较高的正午时分出现最大值,最小值出现在凌晨,表明海水中的CO主要是由CDOM光化学降解产生。
(3)秋季长江口及邻近海域CO微生物消耗速率处在0.21~1.12 h-1之间,平均值为(0.62±0.31)h-1。海水中CO微生物消耗趋势呈指数型降低,微生物消耗为海水中CO的主要去除途径。研究发现盐度是影响微生物消耗的一个重要因素,在盐度为0~19时,kbio随盐度升高而升高;在盐度大于19时,kbio随盐度升高而降低,二者具有一定的相关性。
(4)秋季长江口及邻近海域表层海水中CO含量处于过饱和状态,且具有较大的海-气释放通量,表明研究海域是大气中CO的源。
猜你喜欢
法人(2022年3期)2022-03-30
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
河南水产(2020年1期)2020-06-29
水产科学(2020年3期)2020-05-25
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
华东师范大学学报(自然科学版)(2019年2期)2019-06-11
水运管理(2017年10期)2017-11-20
中国新技术新产品(2015年3期)2015-07-19
