
大鼠耳蜗传出神经系统发育中降钙素基因相关肽和乙酰胆碱酯酶的时空分布
2020-11-15 03:51丁大连张建辉杨琨蒋海燕亓卫东时海波刘芳李鹏RichardSalvi
听力学及言语疾病杂志 2020年6期
丁大连 张建辉 杨琨 蒋海燕 亓卫东 时海波 刘芳 李鹏 Richard Salvi
某些哺乳类动物,如:人类或常用实验动物豚鼠,耳蜗感觉上皮和周边神经系统在出生前已经基本发育完善,因而从一出生就具备了基本正常的听觉反应[1,2],但是大鼠或小鼠的耳蜗在出生时尚未发育完全,一般要在出生后15天才开始出现部分听觉反应,在出生后18天以上才开始具备接近正常的听觉反应能力[3,4];因此,处于婴幼儿阶段的大鼠或小鼠或许更适合于研究听觉的发生和耳蜗的后期发育过程。
耳蜗与中枢的神经联系包含着传入和传出两个系统,其中传入神经系统是将耳蜗内感觉上皮产生的听觉冲动信号经中枢听觉传入通路传送到各级听觉中枢直至大脑听觉皮层被机体感知,传出神经系统则是位于脑干上橄榄核的由高位听觉中枢下行纤维调控的参与维持听觉敏感度和参与调控和保护听觉器官的耳蜗传出神经元。从耳蜗传出神经元发出的大部分神经纤维下行分布到耳蜗的感觉上皮,少部分传出神经纤维分布到耳蜗核。耳蜗传出神经主要包括调控外毛细胞的内侧橄榄耳蜗(medial olovocochlear, MOC)和调控内毛细胞底部传入神经纤维的外侧橄榄耳蜗(lateral olovocochlear system, LOC)两支不同的传出神经系统[5~7]。传出神经系统通过神经递质的释放使神经冲动信号在神经元与效应器之间或神经元与神经元之间传递,传出神经的受体包含多种神经活性物质,例如:乙酰胆碱、降钙素基因相关肽(calcitonin gene-related peptide,CGRP)、r-氨基丁酸(GABA),多巴胺、脑啡肽及强啡肽等,目前接受胆碱能受体的乙酰胆碱酯酶(acetylcholinesterase,AchE)和参与伤害性信息传递的CGRP被认为是耳蜗内传出神经递质的主要受体,因此,AchE和CGRP的组织化学反应产物被较多地作为耳蜗传出神经末梢的特异性标记[8~13]。为了了解耳蜗传出神经在耳蜗后期发育过程中抵达耳蜗的确切时间和AchE及CGRP这两种耳蜗传出神经末梢标记蛋白在耳蜗螺旋器中表达的异同,本研究观察了出生后3~15天及出生后4个月的大鼠耳蜗中AchE和CGRP的时空分布,报告如下。
1 材料与方法
1.1实验动物及分组 受试动物为出生后0、3、5、7、10、12、15和120天的Sprague Dawley大鼠(Charles River Laboratories, Wilmington, MA)各5只,所有动物的使用和饲养程序均按照美国国立卫生研究院发布指南并得到布法罗大学实验动物管理委员会的批准。
1.2全耳蜗基底膜取材 所有受试大鼠的颞骨在指定的出生后不同时间被取出并立刻浸入10%福尔马林PBS溶液固定3小时,然后在解剖显微镜下常规分离取出全耳蜗基底膜[10~14]。
1.3免疫组织化学染色方法 在荧光显微镜下观察耳蜗神经纤维及末梢的免疫组织化学染色方法与先前所采用的方法[12,14]基本相同。应用Mouse anti-Tublin抗体标记出生后不同时间大鼠耳蜗的听神经纤维及其神经末梢,应用Rabbit anti-CGRP抗体标记出生后不同时间大鼠耳蜗传出神经末梢的CGRP,应用Mouse anti-AchE抗体标记出生后不同时间大鼠耳蜗传出神经末梢的AchE,同时应用ToPro-3复染出生后不同时间大鼠耳蜗基底膜细胞的细胞核。用于免疫荧光标记的耳蜗基底膜在固定后被取出,经PBS漂洗后浸入用含有1% Triton X-100和5%羊血清或驴血清的PBS溶液稀释100倍的一抗溶液(Mouse anti-Tublin抗体,或Rabbit anti-CGRP抗体,或Mouse anti-AchE抗体),在4 ℃冰箱中孵育24小时;然后用PBS溶液充分漂洗样品后,将样品浸入用含有1% Triton X-100和5%羊血清或驴血清的PBS溶液稀释200倍的与一抗和血清相对应的不同荧光标记的二抗溶液孵育2小时,再用ToPro-3染色液标记细胞核30分钟,最后将基底膜平整铺放在载玻片上的甘油滴中,盖上盖玻片,在共聚焦荧光显微镜下,应用不同的激发光分别探测绿色荧光标记的免疫组化产物(λabc/λem=488/520 nm)和红色荧光标记的免疫组化产物(λabc/λem=550/570 nm)以及蓝色荧光标记的细胞核(λabc/λem=642/661 nm)。应用Confocal LSM Image Examiner和Adobe Photoshop 5.5软件对共聚焦图像资料进行不同图像层次和不同荧光信号的采集、合并及处理。
1.4酶组织化学染色方法 应用铜-铅-硫胆碱染色标记技术显示AchE的酶组织化学标记方法与先前所采用的方法[10,11,13]相同。有关AchE的酶组织化学染色技术和耳蜗铺片方法简单介绍如下:首先在室温下将分离取下的耳蜗基底膜浸入到0.1 mol/L醋酸盐缓冲液(pH6.0)浸泡30分钟,然后移入到新鲜配制的AchE孵育液[包含5 mg溶于6.5 ml 0.1 mol/L醋酸盐缓冲液(pH6.0)的碘化乙酰硫胆碱,0.5 ml 0.1 mol/L枸橼酸钠溶液、1 ml 30 mmol/L硫酸铜溶液、1 ml双蒸馏水,1 ml 5 mmol/L铁氰化钾溶液]温育30分钟,用0.1 mol/L醋酸盐缓冲液(pH6.0)漂洗基底膜样品三次后移入1%硫化铵溶液显色1分钟;然后用0.1 mol/L硝酸钠溶液漂洗五次,再将样品浸入0.1%硝酸银溶液1分钟;经0.1 mol/L硝酸钠溶液漂洗1分钟后再用0.1 mol/L醋酸盐缓冲液(pH6.0)漂洗1分钟;最后常规将耳蜗基底膜铺放在载玻片上的甘油滴中,盖上盖玻片,在普通光学显微镜下观察并拍照。
2 结果
2.1耳蜗传入和传出神经纤维在耳蜗基底膜上的时空分布 应用Tublin免疫组织化学方法显示的是耳蜗基底膜上所有的传入和传出神经丝及其末梢的分布,虽然Tublin标记并不能区分这些神经纤维到底是传入神经还是传出神经,但是却可以根据穿越耳蜗Corti隧道(Corti’s tunnel)的具有耳蜗传出神经隧道螺旋束(tunnel spiral bundle)解剖学特点的神经纤维,来判断来自内侧橄榄耳蜗传出神经系统的隧道放射束开始穿越Corti隧道的确切时间。结果显示,耳蜗传出神经的隧道放射束是在大鼠出生后第七天才穿越耳蜗Corti隧道并与外毛细胞建立起突触联系(图1)。
2.2耳蜗基底膜上CGRP的时空分布 根据CGRP免疫组织化学反应标记结果,发现直到出生后第三天,耳蜗基底膜上无CGRP表达(图2a、2b);然而,在大鼠出生后第五天开始CGRP出现在内毛细胞周围和底部(图2c),并且这种特异性表达在耳蜗内毛细胞周围的CGRP将保持终生(图2d~h)。但是值得特别注意的是,在耳蜗传出神经内侧螺旋束的神经纤维并未见CGRP表达,在耳蜗传出神经上隧道放射束的神经纤维和末梢也未见CGRP表达(图2c~h)。
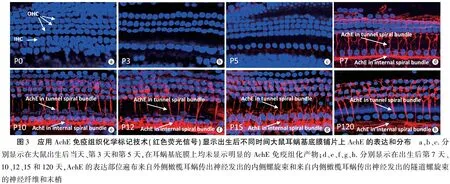
2.3耳蜗基底膜上AchE的时空分布 根据AchE荧光免疫组织化学反应标记结果,直到大鼠出生后第五天AchE尚未进入Corti器(图3a~c);然而,在大鼠出生后第七天AchE却已经同时出现在耳蜗传出神经内侧螺旋束和隧道螺旋束的神经纤维和末梢(图3d~h);应用铜-铅-硫胆碱染色标记AchE的酶组织化学反应产物在普通光学显微镜下显示相同结果(图4a~h)。
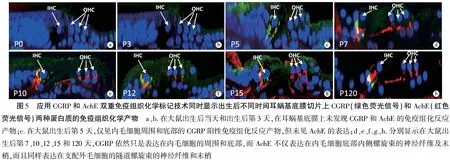
2.4耳蜗基底膜切片上的CGRP和AchE双重标记 为了更好证明CGRP和AchE在耳蜗Corti器的表达部位和出现时间都不相同,除了上述耳蜗基底膜单个蛋白标记的表面制备铺片结果之外,还应用CGRP和AchE双重免疫组织化学方法同时显示大鼠耳蜗基底膜上耳蜗传出神经的CGRP和AchE,并从耳蜗基底膜切片的角度再次证实CGRP和AchE在大鼠耳蜗基底膜上的表达位置和表达时间完全不同(图5)。值得注意的是,如果在内毛细胞底部的CGRP和AchE定位完全相同,表达绿色荧光信号的CGRP和表达红色荧光信号的AchE在重叠部位显示成黄色荧光信号;但是如果两种蛋白的表达位置不相同,则绿色荧光信号和红色荧光信号不会重叠成黄色。从图5d~h都可以看出,绿色荧光信号与红色荧光信号并未重叠,提示CGRP和AchE在内毛细胞底部内侧螺旋束的表达可能并不是在完全相同的部位。
3 讨论
3.1耳蜗传出神经抵达Corti器的时间晚于传入神经系统 根据本实验应用Tublin抗体标记神经丝的免疫组化产物的观察结果,可以确定耳蜗I型和II型传入神经纤维及末梢在大鼠出生的时候已经与耳蜗内外毛细胞建立起突触联系,这与以往的观察结果[14]相一致。但是由脑干上橄榄核耳蜗传出神经元发出的耳蜗传出神经隧道螺旋束的神经纤维在大鼠出生后第七天才抵达Corti器。说明耳蜗传出神经纤维末梢与耳蜗毛细胞之间的突触联系远远滞后于耳蜗的传入神经系统。


3.2CGRP在耳蜗基底膜上的时空分布 据文献报道,大鼠耳蜗内的CGRP在出生后第四天首先出现在大鼠耳蜗底回传出神经内侧螺旋束的神经末梢[15,16],这与本实验在大鼠出生后第三天未发现CGRP但是却在出生后第五天发现内螺旋束神经末梢中强烈表达CGRP的现象相吻合。虽然有文献报道CGRP随后也会出现在外毛细胞底部的传出神经末梢[8,9,17],但是本实验发现相对于内侧螺旋束神经末梢中强烈表达的CGRP免疫组化产物,出现在外毛细胞底部的微弱荧光信号与样品的背景色相同,因此,认为大鼠耳蜗Corti器中的CGRP主要定位在内毛细胞底部内侧螺旋束的神经末梢,但是既不表达在传出神经内侧螺旋束的神经纤维,也不出现在支配外毛细胞的隧道螺旋束的神经纤维及末梢。

CGRP作为神经肽的降钙素肽家族成员之一遍布全身各个系统,参与调控机体各种复杂的生理功能。CGRP虽然在耳蜗内侧螺旋束的传出神经末梢有强烈的表达,但充其量只能说明出现在耳蜗传出神经内侧螺旋束神经末梢的CGRP或许只是在这里参与疼痛感受或者参与钙稳态维持,却并不一定意味着CGRP可以作为耳蜗传出神经系统特有的独特蛋白,因为CGRP的表达毕竟遍及全身各个器官,而且并没有特异性表达在整个耳蜗传出神经系统。
3.3AchE在耳蜗基底膜上的时空分布 作为传出神经系统的特异性标志酶之一,AchE的功能是将乙酰胆碱分解为胆碱和乙酸以阻止胆碱能神经递质对突触后膜的过度兴奋作用。Zheng等曾将AchE用于证明耳蜗传出神经切除手术的成功与否,当耳蜗传出神经随着前庭下神经被成功切除而消失之后,耳蜗内的AchE不复存在[13]。本实验结果再次证实大鼠耳蜗内的AchE遍布调控内毛细胞底部传入神经纤维的外侧橄榄耳蜗传出神经纤维及其末梢和调控外毛细胞的内侧橄榄耳蜗传出神经纤维及其末梢,这显然与仅仅表达在内侧螺旋束神经末梢的CGRP完全不同。从这个现象可以看出,虽然AchE和CGRP都被认为是耳蜗传出神经的标志性蛋白质,但是前者表达在所有传出神经的细胞体和神经纤维及其末梢,而后者仅仅出现在耳蜗传出神经内侧螺旋束的神经末梢。如此看来,标记AchE显然更有利于显示全耳蜗的传出神经系统。本实验结果证实,无论应用铜-铅-硫胆碱的酶组织化学染色技术还是应用免疫组织化学方法标记AchE,都使我们有可能准确判断耳蜗传出神经纤维在出生后何时入住到大鼠耳蜗Corti器。本研究发现AchE标记的耳蜗传出神经纤维在大鼠出生后第五天尚未抵达Corti器,直到出生后第七天才在基底膜上同时形成内毛细胞下方的内侧螺旋束和穿越Corti隧道与外毛细胞建立突触联系的隧道螺旋束,可见出生后第六天很可能是耳蜗传出神经开始进入大鼠耳蜗基底膜的关键时刻。至于耳蜗传出神经纤维中AchE的表达会不会有可能稍许滞后于神经丝结构的出现,根据本研究显示穿越Corti隧道的呈典型放射状排布的隧道螺旋束传出神经纤维解剖学特征(图1d~h,图3d~h,图4d~h),可以确定大鼠耳蜗传出神经在出生后第五天之前确实没有抵达Corti隧道,而AchE的表达位置与耳蜗传出神经纤维及其末梢的位置完全相同。
3.4传出神经纤维趋向性生长的引导信号 本实验发现,CGRP在大鼠出生后第五天已经出现在内毛细胞的下方,而AchE标记的耳蜗传出神经纤维在大鼠出生后第七天才在基底膜上形成内侧螺旋束和隧道螺旋束并与内毛细胞底部的I型传入神经纤维和外毛细胞的细胞体建立起突触联系,说明CGRP蛋白在耳蜗传出神经纤维进入耳蜗Corti器的前两天就已经提前出现在内毛细胞的下方。既然用AchE标记的具备胆碱能受体的耳蜗传出神经纤维在大鼠出生后第七天之前尚未抵达耳蜗基底膜,说明预先出现在内毛细胞下方的CGRP并不是伴随着耳蜗传出神经末梢在Corti器的就位才开始出现,由此引出一系列需要思考的问题,那就是CGRP到底是不是来源于耳蜗传出神经系统?预先出现在内毛细胞下方的CGRP会不会只是一种由内毛细胞或与内毛细胞相联系的I型传入神经末梢释放的诱导内侧螺旋束耳蜗传出神经纤维的引导信号?这是值得深入思考和探索的下一步研究课题。在神经纤维生长的过程中,位于轴突生长尖端的生长锥具有“嗅探”引导信号的能力,从而使神经纤维的生长方向遵循着精确路径找到靶目标并与之建立起精准的突触联系。例如,内耳感觉毛细胞在胚胎期与传入神经建立联系的分化发育过程,既要依赖于毛细胞分泌神经营养因子的吸引,同时,也要依赖于神经节对假复层上皮的引导信号[18],而耳蜗传出神经在寻找靶目标的过程可能同样离不开分化中Corti器细胞释放出的诸如钙离子或乙酰胆碱类物质的引导信号。据文献报道,出生后当天的大鼠耳蜗内毛细胞就已经表现出胆碱能传出输入的生理学反应[19]。鉴于从脑干上橄榄核耳蜗传出神经元发出的传出神经纤维此时尚未进入到耳蜗Corti器,那么这些最早出现在内毛细胞的乙酰胆碱很可能并不是由耳蜗传出神经释放,而是由处于早期分化发育过程中的内毛细胞释放并成为一种引导耳蜗胆碱能传出神经纤维生长方向的引导信号。除了内毛细胞之外,分化发育中的耳蜗支持细胞也可能参与了对神经突触发生的引导和调控作用,例如:在耳蜗的早期分化过程中,位于内侧螺旋沟的Kolliker 细胞及内毛细胞周围的支持细胞在释放ATP的过程中通过激活内毛细胞内的嘌呤能受体而导致了钙离子的释放[20]。鉴于CGRP的功能与钙的稳态调节有关,其表达可能会趋向于钙波释放活跃的靶细胞;而AchE的功能与乙酰胆碱的降解有关,其表达也有可能会趋向于乙酰胆碱释放活跃的靶目标。如此看来,内毛细胞或位于内侧螺旋沟的Kolliker细胞在早期分化发育阶段释放出的钙离子和乙酰胆碱类生物化学信号是否有可能成为诱导耳蜗传出神经纤维趋向性生长的引导信号,还有待进一步的实验研究予以澄清。
猜你喜欢
现代仪器与医疗(2022年2期)2022-08-11
中国典型病例大全(2022年13期)2022-05-10
中国典型病例大全(2022年9期)2022-04-19
中国听力语言康复科学杂志(2021年6期)2021-12-21
中国听力语言康复科学杂志(2021年6期)2021-12-21
听力学及言语疾病杂志(2020年4期)2020-12-20
复旦学报(医学版)(2020年2期)2020-04-17
电脑报(2020年4期)2020-03-25
发明与创新·中学生(2019年3期)2019-03-20
数学大王·低年级(2015年6期)2015-07-09
