
45份红三叶种质表观性状变异研究
2020-11-18 04:53张鹤山王志勇陈志宏熊军波陆娇云
江西农业大学学报 2020年5期
张鹤山 ,王志勇,陈志宏,熊军波,田 宏,陆娇云,刘 洋*
(1.湖北省农业科学院畜牧兽医研究所,湖北武汉 430064;2.信阳农林学院园艺学院,河南信阳 464000;3.全国畜牧总站,北京 100125)
【研究意义】红三叶(Trifolium pratenseL.)是世界上栽培最为广泛的豆科牧草之一[1],我国西北部、西南部及长江流域进行大面积栽培利用。据统计,2017 年我国主要红三叶栽培地区甘肃红三叶栽培面积3.61万hm2,湖北3万hm2,重庆5.6万hm2[2],湖北省仅巴东地区就分布有7万余hm2红三叶人工草地[3]。红三叶在我国高海拔地区与黑麦草(Lolium perenneL.)、苇状羊茅(Festuca arundinaceaSchreb.)等禾草混种可建成优良人工草地,在四川、贵州、湖北等地与玉米等作物间作,改良土壤,提高作物产量,供作优良绿肥,为当地草地畜牧业生产发展和增加农牧民经济收入发挥了重要作用。除此之外,红三叶在医药及保健品领域亦被开发利用[4-6]。红三叶起源于地中海地区,但在世界各地均有分布,并适合多种土壤气候条件[7],我国新疆、贵州、云南等地有野生(逸生)资源分布。红三叶的这种广泛适应性主要是由于物种本身拥有许多适应当地环境条件的基因型[8]。实际上,红三叶是异花授粉植物,存在配子体自交不亲和现象,使得群体内或群体间具有显著的性状差异,这种差异也为红三叶育种亲本材料的选择提供机会[9]。我国目前登记的“巴东”、“岷山”、“巫溪”和“贵州”等4个红三叶品种就是由地方资源经选择培育而成的。因此,对红三叶种质表观性状变异的分析是进一步利用红三叶种质的基础。【前人研究进展】植物的表型性状是植物内在组织生长分化的外在表现,是植物适应环境变异最直接的体现[10],也是检测遗传变异和多样性的最直接简便的方法[11]。国内外学者对红三叶品种间或群体内的表观性状变异及多样性进行了许多研究。Bulinska-Radomska 等[12]利用21个表观性状分析了三个草属15个种的变异关系,发现种间差异水平在0.05~0.60;Greene 等[13]利用15 个表观性状对33 个红三叶种质材料进行了变异分析,发现14 个性状在不同材料间具有显著差异;Paula等[9]利用17个表观性状对大田和温室的57份红三叶栽培或野生材料进行表观性状变异分析,表明材料间具有比较大的性状变异,并以花期将材料划分为不同的5个类群。在国内,张鹤山[3]对湖北地区的25个野生红三叶群体间和群体内11个表观性状进行了变异分析,变异系数范围为5.93%~34.57%;王建丽[14]对20份引进红三叶资源的9个生长方面的农艺性状进行了变异分析,发现顶生叶片大小变异系数最大,为26.16%。这些研究都表明红三叶无论群体间还是群体内都存在较大的性状变异和丰富的群体遗传多样性。【本研究切入点】表观性状变异分析是进行遗传多样性分析、筛选优良种质资源的有效措施。我国生产中使用的红三叶品种主要来自国外,由于地域差异,引进种质有时会表现出优异的特性。如何筛选出优异种质材料用于红三叶育种是亟需解决的问题。【拟解决的关键问题】本研究拟对红三叶表观性状的变异进行研究,并对种质材料进行聚类分析,旨在筛选有利用价值的种质材料,为红三叶育种提供基础亲本材料和科学依据。
1 材料与方法
1.1 试验材料
试验在2018 年进行。试验材料为来自国内外的45 份红三叶种质,由全国畜牧总站提供,材料信息见表1。

表1 红三叶材料来源信息Tab.1 Information on origin of red clover
1.2 试验方法
采用随机区组设计,每个材料3个重复小区,每个小区种植30株,行距50 cm,株距60 cm。所有材料在2016年秋季种植,各性状在植株生长第3年时测定。
试验地点位于武汉市江夏区牧草种质资源圃,地理位置东经114°10′,北纬30°18′,海拔31 m,属亚热带北缘季风气候,光照充足,热量丰富,雨水充沛,年平均温度16.7 ℃,年降水量1 200 mm左右,无霜期270 d;该地区为丘陵岗地,土壤类型为丘陵黄土,酸性;土壤有机质含量1.8%,pH 值5.69,速效氮、磷、钾的含量分别为88.2 mg/kg、22 mg/kg和87 mg/kg。
1.3 测定项目与方法
测定方法参照《三叶草种质资源描述规范和数据标准》[17]执行,具体指标及方法如下:
茎长度:在开花期,以整个小区植株为观测对象,随机抽取开花的植株10株,测量植株主茎长度,单位为cm。
茎分枝:在开花期,以整个小区植株为观测对象,随机抽取开花的植株10株,采用目测法计数每个植株主茎分枝的数目,单位为枝/株。
茎节数:在开花期,以整个小区植株为观测对象,随机抽取开花的植株10株,采用目测方法观测植株主茎第一个节到主茎末梢的节数,单位为节。
茎节长度:在开花期,以整个小区植株为观测对象,随机抽取开花的植株10株,测量主茎第四个节间的长度,单位为cm。
茎直径:在开花期,以整个小区植株为观测对象,随机抽取开花的植株10株,卡尺测量主茎第四个节间的直径,单位为cm。
顶生叶片长度:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部顶生小叶从叶颈至叶尖的绝对长度,单位为cm。
顶生叶片宽度:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部顶生小叶最宽处的绝对长度,单位为cm。
侧生叶片长度:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部侧生小叶从叶颈至叶尖的绝对长度,单位为cm;
侧生叶片宽度:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部侧生小叶最宽处的绝对长度,单位为cm。
叶形指数:顶生小叶长度与顶生小叶宽度的比值。
叶柄长度:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部叶柄的长度,单位为cm。
叶柄直径:在开花期,在试验小区内随机抽取开花的植株10 株,分别测量每一株中部叶柄中间部位的直径,单位为mm。
单株花序数:在盛花期,在试验小区内随机抽取开花植株10 株,采用目测法观测计数每株上的花序数,单位为个。
每花序小花数:在盛花期,在试验小区内随机选取主茎从上往下数第二个节上的花序10 个,计数花序上的小花数,单位为朵。
花序长度:在盛花期,在试验小区内随机抽取开花植株10株,测量花序的长度,单位为cm。
花序宽度:在盛花期,在试验小区内随机抽取开花植株10 株,测量花序最宽处的直径,单位为cm。
种子长度:采集风干后成熟干籽粒为观测对象,随机抽取10粒干种子,测量最长处的长度,单位为mm。
种子宽度:采集风干后成熟干籽粒为观测对象,随机抽取10粒干种子,测量最宽处的长度,单位为mm。
种子千粒质量:采集风干后成熟干籽粒为观测对象,随机取样,4 次重复,每个重复1 000 粒种子,用1/1000天平称量,单位为g。
1.4 数据处理
用Excel 软件计算各性状平均值、标准差和变异系数;用SPSS 软件进行方差分析、主成分分析和聚类分析。
2 结果与分析
2.1 表观性状变异分析
2.1.1 群体间性状变异 45份红三叶种质材料的19个表观性状变异情况见表2。所有性状在群体间均产生了不同程度的遗传分化,变异系数范围为6.65%~20.57%。其中,茎节长度在不同材料群体间具有最大程度的分化,变化范围为3.1~4.3 cm,平均值10.2 cm,变异系数为20.57%;其次为单株花序数,变异系数为20.41%,指标变幅为75~136朵;分化程度最小的性状是顶生叶片长度,变异系数仅为6.65%,其次为顶生叶片宽度,变异系数6.72%。经方差分析(F 检测),19 个表观性状在不同群体间的差异均达到显著水平(P<0.05),其中12 个性状(茎长度、茎分枝、茎节数、茎节长度、茎直径、侧生叶片宽、叶柄长度、叶柄直径、单株花序数、花序小花数、花序长度以及种子宽度)的变异系数均超过10%,也即说明这些性状在红三叶不同群体间具有较大程度的变异。因此,为了更有效地研究红三叶种质的多样性,以这12个性状数据为基础对后续的相关性、聚类以及主成分进行分析。

表2 红三叶群体间及群体内性状变异分析Tab.2 Analysis of variance for all morphological traits of red clover
2.1.2 群体内性状变异 红三叶种质群体内各性状均具有显著的变异分化。为研究红三叶种质群体内的性状分化程度,随机选择两个种质为例进行性状变异分析(CF0022184 和CF022236),结果见表2。两个种质群体内各性状均具有不同程度的变异,变异系数范围分别为3.98%~29.41%和1.99%~31.58%且差异达到显著水平(P<0.05)。种质CF022184 群体内性状分化程度最大的是单株花序数,变异系数为29.41%,其次为茎节数,变异系数为24.14%;变异系数最小的是种子千粒质量,仅为3.98%。19个性状中有11 个性状变异系数大于10%。种质CF022236 群体内变异系数最大的是叶柄长度,变异系数为31.58%,其次为茎直径(22.89%);茎长度、茎分枝等14 个性状变异系数大于10%,说明该群体内存在广泛的性状变异。在两个种质内,变异程度最低的性状都为种子千粒质量,这与张鹤山等[3]对湖北地区野生红三叶种质研究结果一致。
2.2 表观性状间相关性分析
基于材料间变异系数大于10%的12 个性状指标分析了各性状间的相关关系,结果见表3。不同表观性状间具有不同程度的正向或负向相关性。茎长度与茎节数和叶柄长度呈显著正相关(P<0.05),说明红三叶株高(茎长度)越高,其茎节数越多、叶柄越长。茎分枝与其他性状有正或负相关性,但均没有达到显著水平。茎节数和叶柄长度及叶柄直径具有极显著正相关;茎节长度和花序小花数及茎直径呈显著正相关,但与叶柄直径显著负相关;叶柄长度与花序小花数呈显著正相关;而种子宽度与花序长度具有显著正相关关系,说明花序越长,种子越宽。
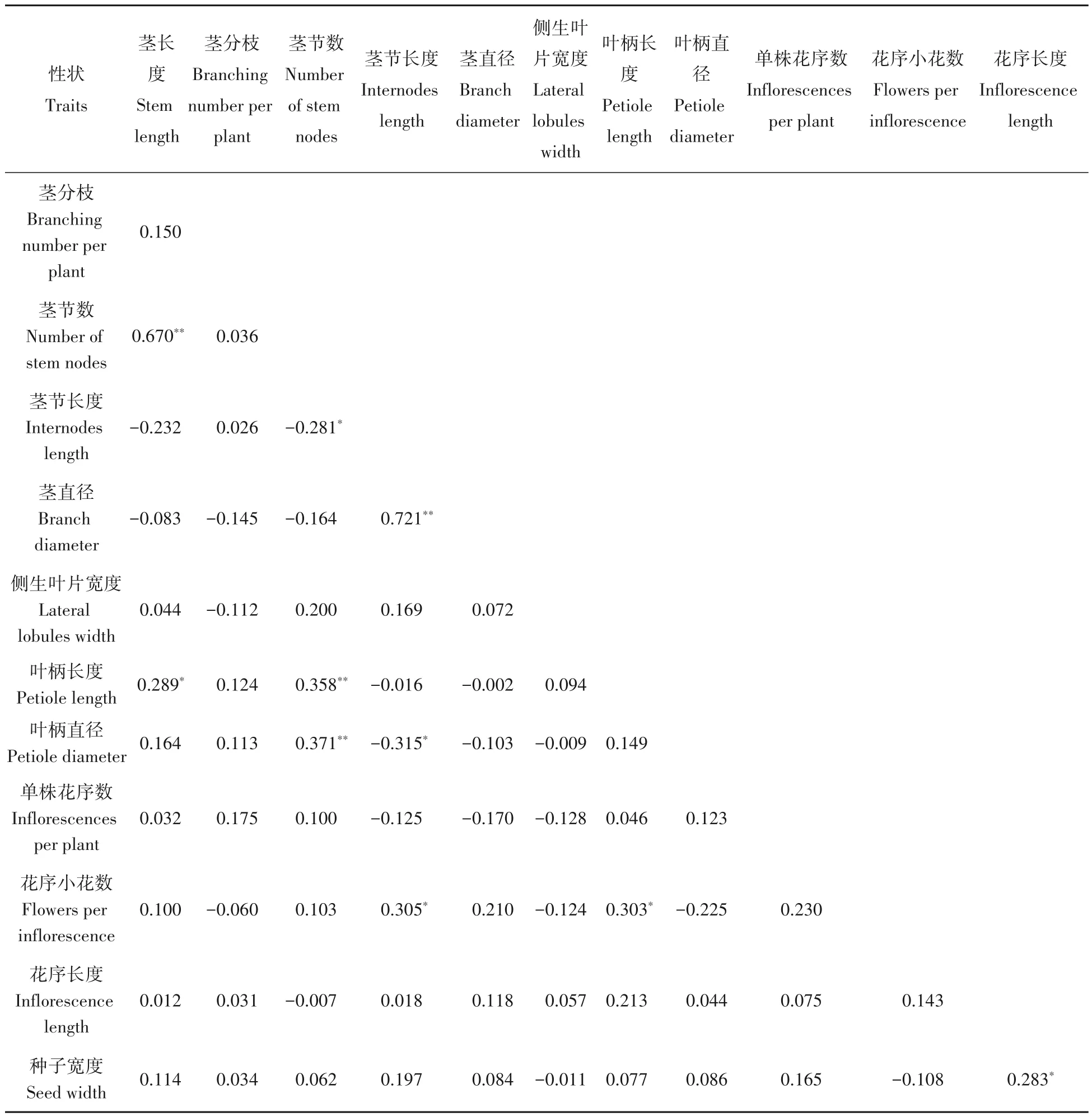
表3 45份红三叶材料表观性状间遗传相似系数Tab.3 Genetic similarity coefficient between traits in 45 populations of red clover
2.3 聚类分析
聚类分析表明,在欧式距离15.0 处可将45 份红三叶种质资源分为4 个类群(图1)。第I 类群包括34 份种质,几乎囊括所有来源国的红三叶材料,来自中国的3 份红三叶种质包含在这一类群中;第II 类群包含4份种质,来自匈牙利、俄罗斯、罗马尼亚、英国4个不同的国家;第III类群有4份种质,来自美国、英国、匈牙利3个国家;第VI类群包括3份种质,来自美国、英国和捷克斯洛伐克。可以看出,基于12个表观性状的聚类分析并没有将同一地区的种质聚在同一个类群,表明了红三叶种质材料丰富的形态多样性。

图1 基于表观性状的45份红三叶遗传距离Fig.1 Genetic distance among 45 red clover populations based on morphological data
各类群表观性状特征见表4。相较于其他类群,第I 类群种质表观性状指标处于平均水平,植株偏矮,单株花序数和单花序小花数较多分别达到107个和115朵,种子生产潜力较大;第II类群种质特征为植株高大、叶片大、花序多,其平均茎长度为92.9 cm,平均侧生小叶宽度2.7 cm,单株花序数为118个/株,种子宽度为1.24 mm,说明了该类群种质具有较高的地上生物产量和种子生产性能;第III类群整体形态特征为植株高,单株分枝能力强;第VI 类群表观特征较为明显,属于植株矮小的一类,平均茎长度仅为57.3 cm,并且单株花序数和花序小花数均较低,说明其牧草生产和种子生产潜力价值较低。

表4 红三叶种质每个类群表观性状的平均值Tab.4 Mean of morphological traits in each cluster obtained in a subset of populations of red clover germplasm
3 讨论
红三叶为配子体自交不亲和植物,其群体间或群体内存在较大的性状变异分化潜力,这些具有不同特征的种质材料为红三叶育种工作提供了丰富的原始亲本材料,是品种选育工作的基础。结果表明,45 份红三叶群体间和群体内都存在大的表观性状变异,这与前人研究结果一致[3,14,18-19]。但是,具体在表观性状上,同一性状在不同材料间的变异程度大小是不一样的。试验结果表明,红三叶群体间变异分化最大的性状是茎节长度和单株花序数,变异系数分别为20.57%和20.41%,这与他人研究结果有所不同。王建丽等[14]对来自俄罗斯的20 份红三叶表观性状分析证明顶生小叶大小性状变异程度最大,而孟丽娟等[19]发现红三叶不同种质间叶面积分化程度最大。由此可见,红三叶种质间的变异大小不仅与材料有关,还可能受生长环境和生态区域的影响。
表观性状评价是红三叶种质资源筛选的重要依据[20],但红三叶表观形态性状复杂多样,为其评价工作带来难度。前人研究表明,红三叶植株叶面积和开花期具有较高的正相关性,叶片越大,开花期越晚[22],但单株花序数与其他性状并没有显著相关性[9]。本研究中发现茎长度和茎节数、叶柄长度呈显著正相关,但单株花序数与其他表观性状不具显著相关性。
通过表观性状可以有效地将具有同一性状属性的红三叶种质聚为一类,许多研究也证明了这种方法的有效性[13,22]。本研究以12个表观性状将45份红三叶种质资源划分为4个类群,每个类群具有明显差异的形态特征,如第II 类群种质其植株高大、单株花序数多,第III 类群种质具有更强的单株分枝能力。研究表明,单株花序数和单株茎分枝数是影响红三叶种子生产的重要因子[15,23],因此本研究结果可以为高种子产量红三叶品种选育提供帮助。尽管通过聚类分析可将具有相近表观性状的红三叶种质材料划分到同一个类群,但就种质来源地而言,来自同一地域的种质材料并未被聚在同一或相近的类群。实际上,这种现象在异花授粉植物如紫花苜蓿(Medicago sativa)[21]和红三叶(Trifolium pratense)[9]中已经被发现。一方面,可能是因为选育过程中大多采用综合品种的方法,无论地方品种还是自然材料,其种质为许多杂合单株组成混合群体[24];另一方面,异花授粉的特性也会引起个体差异大于品种间差异的现象,抵消了来源地生态环境的影响[19]。聚类分析能够突出同类的特点和异类之间的差异,在红三叶育种和生产中,可以根据育种目标选择相应特点的亲本材料,减少育种过程中的盲目性,提高育种效率。
4 结论
红三叶种质无论群体间还是群体内都存在较大程度的表观性状变异分化,茎节长度在各群体间的变异系数最大,为20.57%,而顶生叶片长度变异分化程度最小,变异系数仅为6.65%。红三叶株高(茎长度)越高,其茎节数越多、叶柄越长;茎节长度和花序小花数及茎直径呈显著正相关,而种子宽度与花序长度具有显著正相关关系。45份红三叶种质材料根据植株形态可划分为4个类群,第I类群包含34个种质,占所有种质的75%,其表观特征为植株高度中等,种子生产潜力较大;第II类群和第III类群均包含4个种质,其特征是植株高大,具有较高的地上生物产量生产能力;第VI类群包括3个种质,其明显特征是植株低矮,平均株高57.3 cm。可见,种质材料间具有显著的群体特征和性状差异,可为红三叶育种和生产提供科学参考。
猜你喜欢
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14
河北果树(2021年4期)2021-12-02
生态学报(2021年3期)2021-03-31
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
福建基础教育研究(2019年10期)2019-05-28
心肺血管病杂志(2018年11期)2018-12-18
果农之友(2018年5期)2018-10-19
广东农业科学(2017年5期)2017-08-29
科学种养(2017年6期)2017-06-13
