
金丝楸优良无性系遗传多样性和亲缘关系的AFLP 分析
2020-12-08 08:03马庆国王治军徐慧敏李明昆李慧民张俊佩张建国
林业科学研究 2020年5期
马庆国,王治军,徐慧敏,李明昆,李慧民,张俊佩*,张建国
(1. 林木遗传育种国家重点实验室,国家林业和草原局林木培育重点实验室,中国林业科学研究院林业研究所,北京 100091;2. 洛阳农林科学院,河南 洛阳 471000;3. 洛宁县林业技术指导站,河南 洛阳 471700;4. 和田地区林业和草原局,新疆 和田 848000;5. 喀什地区林业技术推广站,新疆 喀什 844000)
楸树(Catalpa bungei C. A. Mey.)原产中国,是紫葳科(Bignoniaceae)梓属(Catalpa Scop.)的高大落叶乔木,由于其对环境的适应性较好、生长速度迅速、木材材质优良,同时具用材和观赏两方面的特性,是理想的复合农林业建设树种[1]。金丝楸是楸树中的类型之一,在河南、山东、江苏、安徽等省有自然分布,由于它干型好、生长快、材性好、木材剖面色泽金黄,因此,在楸树适生区域备受青睐。河南是我国金丝楸的主产区之一,深入研究金丝楸优异种质资源,认识其遗传多样性水平、明确其亲缘关系,可以为金丝楸种质资源的保护和利用提供理论支持。
楸树种质资源研究始于20 世纪70 年代,早期主要是采用形态学的方法,姚庆渭等[2]研究了分布于亚洲东部和北美洲的梓树属不同种的形态学特征,建议设立梓树组、大梓树组和楸树组,其中,楸树组包括楸树和灰楸两个种。潘庆凯等[3]首次将楸树和灰楸2 个种的种质资源从生产利用角度划分成了不同类型并进行了详尽的描述。河南省楸树研究组在1983 年至1985 年期间对楸树和灰楸资源开展了全国范围的调查,将我国的楸树种质资源在种以下划分成了11 个类型,这是我国目前最为系统的楸树资源调查和分类研究[4]。分子标记技术的广泛应用使得楸树种质资源的研究已经深入到了分子水平。Li 等[5]采用叶绿体ndhF 序列和核糖体DNA序列对紫葳科梓属的系统发育关系进行了探讨。石欣等[6]采集了156 个楸树不同类型的单株材料,利用 ISSR 分子标记分析发现,其遗传变异主要存在于类型内,且聚类分析发现,10 个类型可划分为4 类。Wang 等[7]从楸树转录组中设计并筛选了30 对EST-SSR 引物,这些引物可应用于楸树及其近缘种的遗传分析。方乐成等[8]应用 SSR 分子标记解析楸树种质资源遗传多样性和亲缘关系,构建了其核心种质。这些研究的开展为楸树种质资源的保存保护和开发利用提供了参考,但在分子水平上对其遗传多样性的研究相对较少,采用 AFLP 技术进行金丝楸遗传多样性分析的研究还未见报道。
扩增片段长度多态性(AFLP)具有重复性好、可靠性强、灵敏度高等优点[9],是进行种质资源遗传多样性和遗传结构研究的有效分子标记方法,目前已在很多多年生木本植物中成熟应用。为了明确金丝楸优良无性系的遗传多样性,解析其与楸树种下不同类型及梓树属其它种之间的亲缘关系,本研究以75 个金丝楸优良无性系单株和梓树属不同类型为研究对象,应用 AFLP 技术分析它们的遗传多样性,建立DNA 指纹图谱,揭示金丝楸优良无性系和楸树不同类型间的遗传关系,为金丝楸品种鉴定提供依据,同时也为确定金丝楸的分类地位、种质资源保护及可持续利用和新品种选育提供参考。
1 材料与方法
1.1 材料
供试材料为定植于我国河南省洛宁县的75 个金丝楸优良无性系、‘豫楸1 号’、光叶楸、灰楸和梓树(图1、表1),采其健康、无病虫为害的嫩叶,低温运输、保存并立即提取其基因组DNA。
1.2 方法
1.2.1 DNA 提取 采用改良的CTAB 法[10]提取其基因组DNA。
1.2.2 引物筛选和重复性验证 从64 对EcoRⅠ+3/Mse Ⅰ+3 引物组合(上海生工)中筛选出10 对(表2)用于本研究。选取‘豫楸1 号’、梓树等5 个样品采用10 对AFLP 引物进行扩增,试验重复3 次,验证其谱带质量和可重复性。
1.2.3 AFLP 分析 对供试样品采用EcoR Ⅰ/MseⅠ内切酶(NEB 公司)进行双酶切,酶切、连接、预扩增、选择性扩增程序参考李辛雷等[11]描述的方法进行。
1.2.4 电泳检测 将甲酰胺与分子量内标(100∶1)混合均匀后取9 μL 进行上样,再将PCR 产物稀释10×后吸取1 μL 加入上样板,使用3730XL 测序仪进行毛细管电泳检测。
1.2.5 数据分析 基于采样地点的经纬度利用Google earth 和Arcmap 生成采样地点分布图。利用GeneMarker 2.2 软件将79 个材料10 对荧光引物毛细管电泳的检测结果与分子量内标进行比较,得到片段大小,然后根据无带和有带的情况转换为“0/1” 矩阵。采用SPSS 22.0 软件的simple match 模块检验AFLP 试验的可重复性。用NTSYSpc 2.11F软件计算DICE 遗传相似系数,并基于相似系数矩阵采用UPGMA 法(非加权算术平均法) 进行聚类分析,通过 Tree plot 模块生成聚类图;用EIGEN模块进行基于Jaccard 遗传相似系数的主坐标分析(PCA)[12],以进一步验证并分析聚类结果。使用POPGENE32 version 1.32 软件[13]计算多态性比例PPBs (percentage of polymorphic bands),并在假定哈迪-温伯格平衡的前提下计算其有效等位基因数(Ne)、基因多样度(H)和Shannon 信息指数(I)[14]。利用SPSS 22.0软件基于遗传距离和地理距离进行Mantel 检验[15]。

图1 供试样品采集地点Fig. 1 Geographic location of samples used in the study

表1 供试金丝楸优良无性系及其来源 Table 1 Superior clones of Jinsiqiu used in the AFLP analysis and their origin
2 结果与分析
2.1 多态性分析
本研究筛选10 对EcoR Ⅰ/Mse Ⅰ引物组合对金丝楸优良无性系基因组DNA 进行AFLP 分析,扩增取得了较好的效果(表2,图2),扩增谱带清晰、位置一致性良好(重复性>92%)。总体水平上,10 对引物组合共扩增出1 177 条带,其中,1 130 条带是多态性带,平均多态性比例为96.01%;对75 个金丝楸无性系而言,总的条带数是1 041条,其中,多态性带811 条,平均的多态性比例是77.89%。其中,引物组合E-TCC/ M-CTT 所检测的多态性比例最高为总体水平100% 及金丝楸93.55%,总条带数分别为111 条(总体)和93 条(金丝楸);引物组合E-AAG/ M-CAA 所获得的总条带数最多,为149 条(总体)和128 条(金丝楸), 其多态性比例为94.63%( 总体) 和75.78%(金丝楸)。
2.2 遗传相似系数
利用10 对选择性引物组合进行AFLP 检测,79 个样品的DICE 遗传相似系数范围为0.413 4~0.905 9,平均为0.811 6。其中,JSQ-54 和JSQ-76的遗传相似系数最大为0.905 9,JSQ-37 和梓树间的遗传相似系数最小,为0.413 4。
2.3 有效等位基因数、基因多样度及Shannon 信息指数分析
基于等位基因频率计算有效等位基因数(Ne)、基因多样度(H)及Shannon 信息指数(I)是进行遗传多样性水平评估的重要方式[12]。表3 表明:10 对引物所检测到的不同位点的有效等位基因数范围为1.00~1.99,平均为1.48±0.03(总体)和1.36±0.04(金丝楸);基因多样度为0.00~0.49,平均为0.28±0.01( 总 体 ) 和0.21±0.02( 金 丝 楸 ) ;Shannon 信息指数为0.00~0.69,平均为0.43±0.02(总体)和0.32±0.04(金丝楸)。75 个金丝楸无性系的有效等位基因数、基因多样度和Shannon 信息指数高于对白榆无性系[16]、核用银杏[17]和古银杏资源[18]的研究结果(Ne=1.33,H= 0.20,I=0.33;Ne=1.33, H=0.21, I=0.34; Ne=1.26、 H=0.17,I=0.27),低于利用ISSR 分子标记对观赏石榴[19]的研究结果(Ne=1.94,H=0.38,I=0.55)和采用SSR 标记对西藏光核桃[20]的研究结果,与利用ISSR 技术对楸树种质资源[6]的分析(Ne=1.48,H=0.29,I=0.45)及王庆军等[21]对对山东石榴品种(Ne=1.38,H=0.23,I=0.35)的研究较接近。

表2 AFLP 分析引物组合及其选择性扩增结果 Table 2 Primers used for AFLP and amplification results of Jinsiqiu

图2 部分金丝楸供试样品的荧光AFLP 电泳图谱(引物组合为E-ACT/M-CAA)Fig. 2 Electrophoresis patterns of partly assayed materials of Jinsiqiu superior clones(Primer combination: E-ACT/ M-CTA)

表3 75 个金丝楸优良无性系遗传多样性度量指标Table 3 Genetic diversity of 75 superior clones of Jinsiqiu
2.4 聚类分析
分析聚类图(图3)发现:当阈值设定为0.46 时,可将79 个供试样品分成2 个大组(A 和B):A 组中仅有梓树(ZiShu),B 组包含了其他78 个单株。分析B 组成员发现:75 个金丝楸无性系首先聚在一起,然后与光叶楸(GYQ)和灰楸(HuiQiu)聚在一起,‘豫楸1 号’则单独聚为一组。
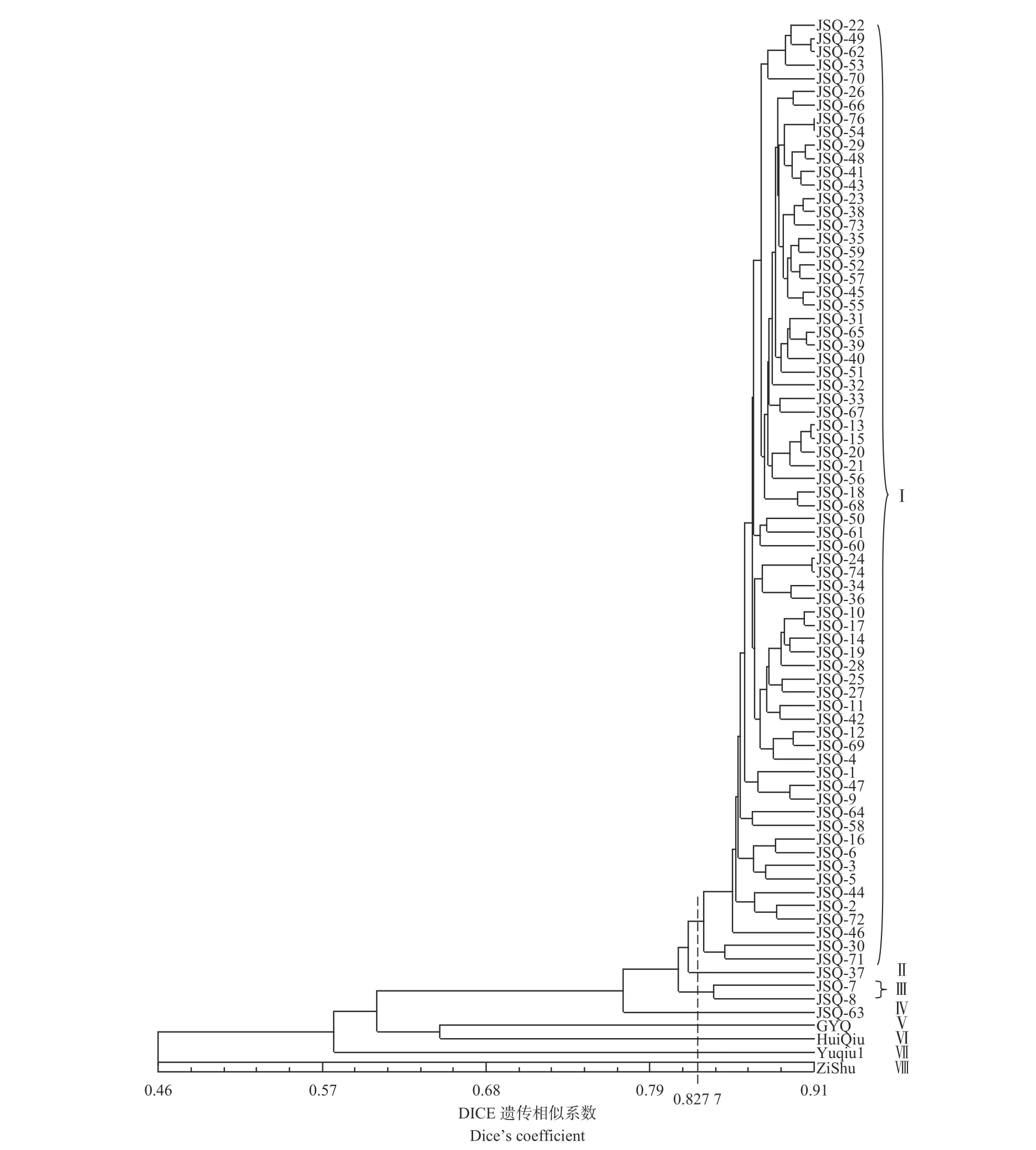
图3 基于AFLP 分析的金丝楸优良无性系UPGMA 聚类图Fig. 3 UPGMA dendrogram of Catalpa bungei superior clones based on genetic distance
当以DICE 遗传相似系数平均值(0.827 7)划分时,可将79 个供试材料分为8 个组(分别标记为Ⅰ~Ⅷ),其中,Ⅰ组成员最多,包含71 个金丝楸无性系,其遗传相似系数为0.806 6~0.905 9,JSQ-30 和JSQ-58 间的遗传相似系数最小为0.806 6, JSQ-54 和JSQ-76 间的遗传相似系数最大为0.905 9,这些无性系的亲缘关系较密切。Ⅲ组包含2 个无性系,即JSQ-7 和JSQ-8(DICE 遗传相似系数0.837 3),它们的母树均采自罗岭乡核桃沟村,亲缘关系较近。JSQ-37 和JSQ-63 分别构成了单独的组(Ⅱ、Ⅳ)。JSQ-37 的母树采自罗岭乡前阴坡村,JSQ-63 的母树采自兴华乡瓦寨庙村,这2 个金丝楸无性系表现出了与其它无性系较远的亲缘关系。GYQ、HuiQiu、Yuqiu1 和ZiShu作为种内和种间对照各自聚为一组(Ⅴ、Ⅵ、Ⅶ和Ⅷ),与金丝楸亲缘关系较远。GYQ 是楸树中不同于金丝楸的一个类型[4],它单独聚为一组。值得注意的是,在本研究中与近缘种灰楸相比,普通楸树品种‘豫楸1 号’与金丝楸的亲缘关系更远。
通过NTSYSpc 2.11F 软件EIGEN 程序进行的PCA 分析表明:前2 个主坐标Dim-1 和Dim-2 所能解释的相关性为84.94%,PCA 分析结果与聚类结果基本一致(图4),这2 种分析相结合可以更全面地反映金丝楸优良无性系的遗传多样性和亲缘关系。进一步对各无性系的遗传距离和地理距离进行相关分析,结果呈极显著正相关(图5),表明地理隔离对楸树遗传多样性有显著的影响。结合聚类分析结果,金丝楸无性系当中的63 号与其他遗传距离最远,值得重点推广。

图4 基于AFLP 数据的金丝楸优良无性系主坐标分析(PCA)Fig. 4 PCA analysis of Catalpa bungei superior clones based on AFLP data

图5 金丝楸优良无性系地理距离和遗传距离的相关分析Fig. 5 Mantel test for the correlation between geographical distance and genetic distance of Catalpa bungei superior clones
3 讨论
楸树是原产我国的特色用材和观赏树种,金丝楸作为其中一个类型具有材色金黄、干性优良等特点,深受消费者喜爱,金丝楸良种的选育也在逐步开展[22],然而,金丝楸的优良品种目前还非常少,对其种质资源遗传多样性和亲缘关系开展分子水平上的研究,有利于深入认识金丝楸种质资源、进一步筛选优秀和特异种质资源以及制定保护和可持续利用策略,同时也可以为金丝楸杂交育种中的亲本选配提供技术支撑。
目前,国内外关于楸树分子标记方面的研究罕有报道,本文采用荧光AFLP 技术,筛选10 对EcoR Ⅰ+3 和MseⅠ+3 引物组合分析75 个金丝楸优良无性系,并以‘豫楸1 号’、光叶楸、灰楸和梓树等不同类型为对照进行遗传结构和亲缘关系分析,获得了高效、稳定的检测结果,供试样品间各引物组合检测到的多态性比例最低为E-ACT/MCTT 的62.86%,最高为E-TCC/M-CTT 的93.55%,表明本研究中所采用的荧光AFLP 技术通过毛细管电泳进行检测能够获得较大的信息量,这与采用同一方法对核桃的研究结果一致[12,14]。同时,本研究中,AFLP 方法检测信息量大、检测效率较高,检测到的大量多态性位点对于金丝楸种质资源的高效、准确鉴定具有重要参考价值。另外,对有效等位基因数、基因多样度及Shannon 信息指数等遗传多样性水平表征指标进行了分析,并与其它树种或本树种的以往研究进行比较,呈现出不同程度的差异性,这可能是由于样品的规模和覆盖度、采用的方法和算法差异等方面的原因导致的。
楸树在暖温带和亚热带范围内广泛分布,但是由于其野生种群很少且自花不育,加之栽培中以无性繁殖为主,一个地区往往只有一个单一无性系存在[6],本研究供试75 个金丝楸无性系之间的遗传相似系数为0.742 4~ 0.905 9, 平均值为0.856 8,无性系间的遗传距离普遍较小,只有JSQ-7、JSQ-8、JSQ-37 和JSQ-63 等4 个无性系表现出比较显著的遗传分化。说明金丝楸无性系间的遗传基础较窄、亲缘关系较近。多年生木本植物种内较大的遗传差异对于杂交育成优良品种具有重要意义[23-24],以4 个具有显著遗传分化的金丝楸无性系作为亲本更易于获得有利变异,可以在今后的金丝楸优良新品种选育中加以利用。与种内其它类型相比较,金丝楸树冠狭窄,树皮呈现块状纵裂,枝条短粗,叶片较大、全缘或1~3 浅裂,尤其是木材呈现明显的特征,其心材呈现黄色,心、边材交界处金黄色,晚材部分的薄壁细胞排列呈与髓射线垂直的大波浪状。本研究聚类分析显示:光叶楸与灰楸首先聚为一组(遗传相似系数为0.649 3),然后与75 个金丝楸聚在一起,之后再与普通楸树即‘豫楸1 号’聚在一起,另一个近缘种梓树则与之亲缘关系更远。因此,形态学性状与AFLP 分析结果均表明金丝楸与普通楸树存在较明显的差异,同时,考虑到本研究中样品数量和技术方法的局限性,在后续研究中宜扩大样品范围、采用高通量技术作更加深入的探讨。
4 结论
本研究以楸树不同种和类型为对照在分子水平上分析了金丝楸优良无性系的遗传多样性水平和亲缘关系, 显示出金丝楸无性系遗传背景较狭窄,各无性系的遗传距离和地理距离呈极显著正相关,表明地理隔离对楸树遗传多样性有明显的影响。形态学性状观察和AFLP 分子标记分析显示:金丝楸与普通楸树存在较显著的差异性,为金丝楸分类地位的确定和品种鉴定提供了新的证据,对于金丝楸优良品种的培育具有重要的理论价值。
猜你喜欢
作物学报(2022年2期)2022-11-06
——致秋天的花楸树
北方人(2021年19期)2021-10-29
毛纺科技(2021年8期)2021-10-14
花卉(2020年22期)2020-01-08
苏州杂志(2019年5期)2019-12-03
山东林业科技(2019年2期)2019-06-03
天然产物研究与开发(2018年10期)2018-11-06
诗歌月刊(2017年10期)2018-01-28
广东农业科学(2017年5期)2017-08-29
语文教学与研究(读写天地)(2012年8期)2012-05-08
