
交枝顶孢发酵液对根结线虫胁迫下番茄防御酶活性的影响
2020-12-14 04:12姚玉荣李林霍建飞郝永娟王万立
中国瓜菜 2020年10期
姚玉荣 李林 霍建飞 郝永娟 王万立

摘 要:为明确根结线虫胁迫下交枝顶孢发酵液对番茄南方根结线虫的作用机制,采用温室苗期人工接种方式,研究了不同浓度的交枝顶孢发酵液对南方根结线虫胁迫下番茄叶片中保护酶的影响。结果显示,接种60 d后交枝顶孢发酵液处理显著降低番茄根结指数,发酵液原液、发酵液稀释2倍液及发酵液稀释4倍液对南方根结线虫的防效为70.32%、62.18%和47.26%。线虫胁迫条件下不同浓度的交枝顶孢发酵液可显著提高根结线虫胁迫下番茄叶片SOD、POD、CAT、PPO和PAL的活性。交枝顶孢发酵液中次级代谢产物对根结线虫病有显著的防控效果。
关键词:番茄;交枝顶孢;根结线虫;发酵液;保护酶
中图分类号:S641.2 文献标识码:A 文章编号:1673-2871(2020)10-059-05
Abstract:To explore the mechanism of Acremonium implicatum fermentation to tomato under the root-knot nematode stress, the effects of nematode infection on five protective enzymes of tomatoes were investigated with or without seedling inoculation in greenhouse. Results showed that after 60 days, the root knot index of tomato treated with fermentation was significantly reduced, and the control effects of the fermentation on Meloidogyne incognita were 70.32%, 62.18% and 47.26% respectively for crude fermatation solution, 2 fold diluted one and 4 fold diluted one. In addition, compared with the control higher levels of SOD, POD, CAT, PPO and PAL activities were observed in the tomatoes pretreated with A. implicatum fermentation and inoculated with nematodes. It indicated that the secondary metabolites of A. implicatum could effectively enhance tomato resistance to root knot nematode.
Key words:Acremonium implicatum; Root knot nematode; Fermentation; Protective enzymes
根结线虫(Meloidogyne incognita)是危害番茄的重要病原之一,每年造成巨大的损失。随着我国农业产业结构调整,保护地蔬菜生产迅猛发展,使得大部分蔬菜产区根结线虫病害日趋严重。目前,生产上对根结线虫病害防治主要为化学杀线剂,但一些杀线剂因高毒、高残留而被禁用,因此亟需开发低毒、高效且环境友好的药剂。真菌次级代谢产物作为新兴的生物农药能够直接作用于植物线虫,作用快、药效高、环境相容性好而成为国内外研究的热门[1-2]。
研究发现,当植物遭受逆境胁迫时,活性氧(reactive oxygen species,ROS)急剧增加,这表明ROS在植物逆境响应中发挥着非常重要的作用。ROS主要包括超氧陰离子(O2·-)、过氧化氢(H2O2)、羟基自由基(·OH)和单线态氧(1O2)等。在植物体内ROS扮演着双重角色,低浓度的ROS可以作为信号分子参与植物生长发育;当植物遭受逆境胁迫时,ROS急剧增加,高浓度的ROS会导致膜质过氧化、破坏细胞膜结构和功能,致使植株受害[3-6]。植物体启动抗氧化酶系统来适应环境变化,抵抗逆境造成的胁迫,氧化酶系统主要包括超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)、多酚氧化酶(polyphenol oxidase, PPO)以及苯丙氨酸解氨酶(phenylalanine ammonialyase, PAL)等[7-8],这些氧化酶活性与植物对根结线虫的抗性紧密相关。研究发现,大豆胞囊线虫胁迫下抗感品种PPO、POD、PAL、SOD以及β-1,3葡聚糖酶活性均显著提高,且抗性品种增幅显著高于感病品种[9];在南方根结线虫胁迫下抗病黄瓜品种SOD、POD、PAL活性显著高于感病黄瓜品种[10]。Molinari等[11]发现,在根结线虫胁迫条件下,番茄体内活性氧积累,引起过敏性反应并诱导番茄获得抗性。
内生真菌交枝顶孢(Acremonium implicatum)对根结线虫有很好的防治效果[12-13],但在根结线虫胁迫条件下用内生真菌交枝顶孢发酵液处理后,植物保护酶活性及其防御相关机制尚不清楚。本研究以南方根结线虫胁迫条件下的番茄植株为研究对象,测定不同浓度交枝顶孢发酵液处理后,番茄叶片中SOD、POD、CAT、PPO和PAL等酶活性,以期明确在根结线虫胁迫条件下交枝顶孢发酵液可以提高番茄植株抗性的机制。
1 材料与方法
1.1 材料
供试菌株:番茄内生真菌交枝顶孢(Acremonium implicatum)菌种保藏号CGMCC8300,由中国农业科学院蔬菜花卉研究所真菌与病毒实验室提供。
供试线虫:南方根结线虫(Meloidogyne incognita),来源于天津市植物保护研究所温室辣椒苗上培养。将感染南方根结线虫的病根洗净,在解剖镜下挑取新鲜卵块,用0.1% NaClO 表面消毒,再用清水洗净置于28 ℃恒温培养箱中孵化,孵化得到南方根结线虫二龄幼虫J2,浓度调至1 000条·mL-1备用。
番茄品种:‘金棚一号,由西安皇冠蔬菜研究所提供。
1.2 发酵液的制备
将保藏的交枝顶孢转接到PDA平板中,于28 ℃恒温培养箱中培养活化。挑取活化的交枝顶孢菌丝接种于装有200 mL PDB的三角瓶中(接种量为每瓶2块d=5 mm的菌块)28 ℃,150 r·min-1培养。发酵液于10 000 r·min-1离心10 min,得到上清液。上清液用0.22 μm的微孔滤膜过滤,得到发酵滤液。
1.3 试验设计
试验于2018年3—4月在天津市现代农业科技创新基地(武清区)温室进行。试验设置8个处理,每个处理3次重复,随机区组排列。
番茄种子经过55 ℃温水浸种30 min后,置于30 ℃左右的温水浸泡6 h,28 ℃培养箱中催芽48 h,待其出芽后点到50孔育苗穴中,置于温室培养。待番茄2片真叶完全展开后移栽到花盆(外径150 mm×高125 mm)中。每个处理分为2组试验,2组试验均用交枝顶孢发酵液原液、发酵液稀释2倍液、发酵液稀释4倍液浇灌处理土壤,等量清水做空白对照。第1组番茄不接种线虫,第2组每株番茄靠近根部接种南方根结线虫1 000条,每个处理15株,3次重复。
1.4 发酵液对根结线虫的防治效果
定植60 d,调查番茄根部根结线虫发病情况并记录病级,计算根结指数及防治效果。病级分级方法—0级:根系无根结;1级:根系有少量小根结;3级:2/3根系有小根结;5级:根系布满小根结并有次生根结;7级:根系形成须根团[14]。
病情指数=Σ(病株数×相应病级数)×100/(调查的总株数×7);
防治效果/%=(空白对照病情指数-处理病情指数)×100/空白对照病情指数。
1.5 番茄叶片抗氧化保护酶活力测定
番茄苗定植60 d后测定叶片中酶活性指标。称取番茄叶片组织,按质量(g)∶体积(mL)=1∶10的比例加入磷酸缓冲液(pH=7.8);将叶片剪碎后放入匀浆中,冰浴充分研磨,离心得到上清液即为酶提取液,每个处理3次重复。可溶性蛋白含量采用BCA法测定,以牛血清蛋白做标样。可溶性蛋白含量以及抗氧化保护酶SOD、POD、CAT、PPO以及PAL的活性测定采用江苏科铭生物技术有限公司生产的BCA法蛋白含量测定试剂盒、超氧化物歧化酶试剂盒(WST-8法)、过氧化物酶试剂盒(愈创木酚法)、过氧化氢酶试剂盒(过氧化氢法)、多酚氧化酶试剂盒(邻苯二酚法)以及苯丙氨酸解氨酶试剂盒(L-苯丙氨酸硼酸缓冲液法)。具体试验步骤以及酶活计算方法参见试剂盒说明书。酶活测定所需的主要仪器包括:多功能微孔板检测仪(BioTek, Synergy H1)、台式离心机(eppendorf, 5430R)、移液器(eppendorf)等。
1.6 数据统计分析
试验数据采用Microsoft office excel 2010分析,统计分析采用SPSS21软件,采用Duncans新复极差法进行显著性分析。
2 结果与分析
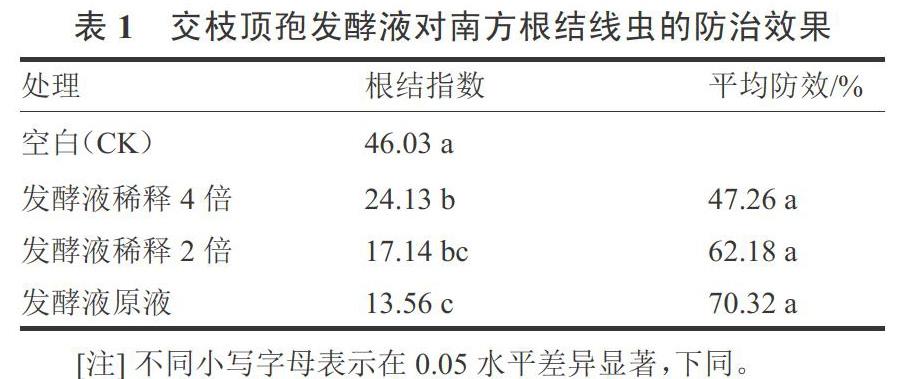
2.1 交枝顶发酵液对南方根结线虫的防治效果
内生真菌交枝顶孢发酵液处理均能够显著降低南方根结线虫对番茄的危害,其中发酵液原液及发酵液稀释2倍液对南方根结线虫防治效果较好,防效分别达到70.32%和62.18%,发酵液稀释4倍液处理的根结指数较高(表1)。
2.2 交枝顶孢代谢产物对番茄超氧化物歧化酶(SOD)活性的影响
交枝顶孢发酵液处理感染南方根结线虫的番茄叶片SOD活力結果见图1。未接种根结线虫时,交枝顶孢发酵液处理后SOD活性显著增加,施用发酵液原液处理SOD活性最高。接种根结线虫后番茄叶片SOD酶活性显著下降,在根结线虫胁迫下,施用不同浓度的交枝顶孢发酵液,番茄叶片SOD活性显著增加,且随着交枝顶孢发酵液浓度的增加SOD活力增加。试验结果表明,交枝顶孢次级代谢产物能够提高番茄叶片SOD活性,增强番茄清除活性氧能力。
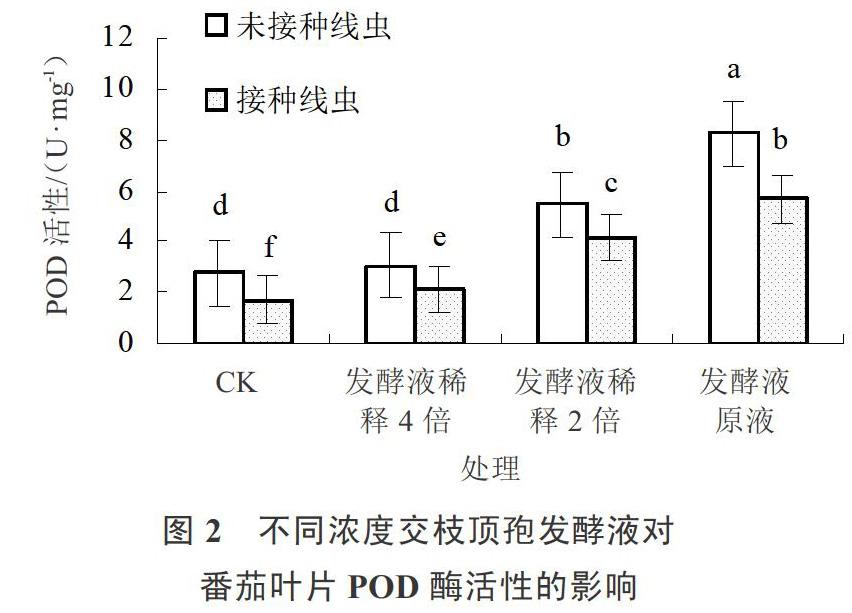
2.3 交枝顶孢代谢产物对番茄过氧化物(POD)活性的影响
交枝顶孢发酵液处理感染南方根结线虫的番茄叶片POD活力结果见图2。未接种根结线虫时,交枝顶孢发酵液处理后POD活性显著增加,施用发酵液原液处理POD活性最高。接种根结线虫后番茄叶片POD酶活性显著下降。在根结线虫胁迫下,不同浓度的交枝顶孢发酵液能够显著提高POD活性,且随着发酵液浓度的增加呈现上升趋势。
2.4 交枝顶孢代谢产物对番茄过氧化氢酶(CAT)活性的影响
交枝顶孢发酵液处理感染南方根结线虫的番茄叶片CAT酶活力结果见图3。未接种根结线虫时,交枝顶孢发酵液处理后CAT酶活性显著增加,发酵液原液处理CAT酶活性最高。接种根结线虫后番茄叶片CAT酶活性显著下降。施用交枝顶孢发酵液后,根结线虫侵染的番茄叶片CAT酶活力显著增加,且随着交枝顶孢发酵液浓度的增加CAT酶活力增加。交枝顶孢发酵液通过提高番茄CAT活性,降低H2O2对番茄的损伤,提高对根结线虫的抗性。
2.5 交枝顶孢代谢产物对番茄多酚氧化酶(PPO)活性的影响
交枝顶孢发酵液处理感染南方根结线虫的番茄叶片PPO酶活力结果见图4。未接种根结线虫时,交枝顶孢发酵液处理后PPO活性显著增加,施用发酵液稀释2倍液处理PPO活性最高。接种根结线虫后番茄叶片PPO活性显著下降。在根结线虫胁迫的条件下,发酵液对PPO酶活力的影响趋势表现为先增加后降低,发酵液稀释2倍液PPO酶活力最强。这一结果表明,发酵液浓度是根结线虫防治的关键因素,必须选择合适的浓度才能够有效防治根结线虫。
2.6 交枝顶孢代谢产物对番茄苯丙氨酸解氨酶(PAL)活性的影响
交枝頂孢发酵液处理感染南方根结线虫的番茄叶片PAL活力结果见图5。未接种根结线虫时,交枝顶孢发酵液处理后PAL活性提高,发酵液原液处理PAL酶活力最高,相比清水对照酶活力增加71.95%。接种根结线虫后番茄叶片PAL活性显著下降。在根结线虫胁迫下,不同浓度的交枝顶孢发酵液处理,番茄叶片PAL活性显著增加,且随着发酵液浓度的增加,PAL酶活力呈上升趋势。
3 讨论与结论
试验结果表明,不同浓度的交枝顶孢发酵液均能降低番茄根结指数,对根结线虫具有较好的防治效果,发酵液原液对番茄根结线虫的防效为70.32%,稀释2倍的防效为62.18%,稀释4倍的防效较低为47.26%。说明交枝顶孢发酵液中对根结线虫起防治作用的主要是发酵液中的次级代谢产物。发酵液中次级代谢产物提高了番茄叶片中防御酶SOD、POD、CAT以及PPO和PAL活性。
研究表明,SOD是植物体内清除自由基的关键酶,作为植物抗氧化酶系统防御的第一道防线,在清除自由基和延缓植物衰老方面发挥重要的作用[15-17]。POD和CAT主要起到酶促降解H2O2的作用[18-20],POD防止膜质过氧化、参与木质素积累[21-22]。PAL是酚类物质、木质素、植保素以及黄酮合成的关键酶,当植物受到病原菌侵染后,PAL活性提高,抗菌物质合成增加[23-27],促进木质素合成,强化植物细胞壁,阻碍线虫侵染[28]。PPO是酚类物质氧化成的关键酶,保护植物免受酚类物质毒害[29-30]。植物可以通过这些抗氧化酶保护系统的协同作用,提高抗性。
试验中番茄经交枝顶孢发酵液处理后,SOD、POD、CAT、PPO及PAL活性均显著提高,这与李颂等[31]对黑曲霉与番茄根结线虫病的研究结果一致。交枝顶孢发酵液使得番茄叶片中防御酶活性提高,这可能是交枝顶孢发酵液中次级代谢产物中诱导因子与番茄体内抗性反应相关,能够激活番茄防御酶活性,诱导番茄对根结线虫的相关抗性反应。在根结线虫胁迫下,细胞产生的ROS能够有效的启动黄瓜抗氧化酶系统[32]。研究发现大豆胞囊线虫胁迫导致大豆生长受阻,叶片膜质透性增加,相对导电率增加,SOD、CAT、POD等保护酶活性降低[33]。杨宇虹等[34]发现,烟草接种南方根结线虫后,抗、感病品种POD活性均提高,但抗病品种增加幅度大于感病品种;陈增齐等[35]研究发现,番茄幼苗经球孢白僵菌发酵液处理后,PAL、POD、PPO及SOD活性显著提高。
综上所述,番茄接种根结线虫后,叶片中防御酶活性降低。交枝顶孢发酵液处理后,在根结线虫胁迫条件下,番茄叶片中抗氧化酶SOD、POD、CAT、PPO以及PAL活性显著提高。说明交枝顶孢发酵液中次级代谢产物直接或间接参与番茄体内防御相关代谢过程,降低根结线虫对番茄的伤害,提高抗性。交枝顶孢发酵液如何影响防御酶活性来诱导植物抗性,仍需进一步深入研究。
参考文献
[1] OYEKANMI E O,COYNE D L,FAGADE O E,et al.Improving root-knot nematode management on two soybean genotypes through the application of Bradyrhizobium japonicum,Trichoderma pseudokoningii and Glomus mosseae in full factorial combinations[J].Crop Protection,2006,26(7):1006-1012.
[2] EL-NAGDI W M A,YOUSSEF M M A.Soaking faba bean seed in some bio-agents as prophylactic treatment for controlling Meloidogyne incognita root-knot nematode infection[J].Journal of Pest Science,2004,77(2):75-78.
[3] DENNESS L,MCKENNA J F,SEGONZAC C,et al.Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species-and jasmonic acid-dependent process in Arabidopsis[J].Plant Physiology,2011,156(3):1364-1374.
[4] 王娟,李德全.逆境条件下植物体内渗透调节物质的积累与活性氧代谢[J].植物学通报,2001(4):459-465.
[5] MILLER G,SHULAEV V,MITTLER R.Reactive oxygen signaling and abiotic stress[J].Physiologia Plantarum,2008,133(3):481-489.
[6] MITTLER R.Oxidative stress,antioxidants and stress tolerance[J]. Trends in Plant Science,2002,7(9): 405-410.
[7] AHMED S,NAWATA E,HOSOKAWA M,et al.Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging[J].Plant Science,2002,163(1):117-123.
[8] 贺文婷,彭德良.植物对线虫胁迫的生理生化响应机制[J].植物保护,2007(2):11-15.
[9] 李海燕,段玉璽,陈立杰,等.大豆胞囊线虫3号生理小种胁迫下不同抗性大豆品种的生化响应[J].大豆科学,2014,33(5):783-786.
[10] 叶德友,钱春桃,王暄.南方根结线虫侵染对黄瓜植株生长和抗氧化系统的影响[J].植物保护学报,2012,39(4):321-326.
[11] MOLINARI S.Changes of catalase and SOD activities in the early response of tomato to Meloidogyne attack[J].Nematologia Mediterranea,1999,27(1):167-172.
[12] TIAN X L,YAO Y R,CHEN G H,et al.Suppression of Meloidogyne incognita by the endophytic fungus Acremonium implicatum from tomato root galls[J].International Journal of Pest Management,2014,60(4):239-245.
[13] YAO Y R,TIAN X L,SHEN B M,et al.Transformation of the endophytic fungus Acremonium implicatum with GFP and evaluation of its biocontrol effect against Meloidogyne incognita[J].World Journal of Microbiology and Biotechnology,2015,31(4):549-556.
[14] 中华人民共和国国家标准农药田间药效试验准则(一)[M].北京:中国标准出版社,2000.
[15] LIU J W,ZHANG R H,ZHANG G C,et al.Effects of soil drought on photosynthetic traits and antioxidant enzyme activities in Hippophae rhamnoides seedlings[J].Journal of Forestry Research,2017,28(2):255-263.
[16] 窦俊辉,喻树迅,范术丽,等.SOD与植物胁迫抗性[J].分子植物育种,2010,8(2):359-364.
[17] MARTINEZ C A,LOUREIRO M E,OLIVA M A,et al.Differential responses of superoxide dismutase in freezing resistant Solanum curtilobum and freezing sensitive Solanum tuberosum subjected to oxidative and water stress[J].Plant Science,2001,160(3):505-515.
[18] 李强,姚霞,孙楷,等.不同光质对茅苍术生长、抗氧化酶活性及挥发油含量的影响[J].中国实验方剂学杂志,2018,24(10):27-32.
[19] ZHANG Y N,YAN X,GUO H M,et al.A novel protein elicitor BAR11 from Saccharothrix yanglingensis Hhs.015 improves plant resistance to pathogens and interacts with catalases as targets[J].Frontiers in Microbiology,2018,9:700.
[20] SAHEBANI N,HADAVI N.Induction of H2O2 and related enzymes in tomato roots infected with root knot nematode (M. javanica) by several chemical and microbial elicitors[J].Biocontrol Science and Technology,2009,19(3):301-313.
[21] DINIZ I,AZINHEIRA H,Figueiredo A,et al.Fungal penetration associated with recognition,signaling and defence-related genes and peroxidase activity during the resistance response of coffee to Colletotrichum kahawae[J].Physiological and Molecular Plant Pathology,2019,105:119-127.
[22] 袁庆华,桂枝,张文淑.苜蓿抗感褐斑病品种内超氧化物歧化酶、过氧化物酶和多酚氧化酶活性的比较[J].草业学报,2002,11(2):100-104.
[23] TREUTTER D.Significance of flavonoids in plant resistance and enhancement of their biosynthesis[J].Plant Biology,2005,7(6):581-591.
[24] TONNESSEN B W,MANOSALVA P,Lang J M,et al.Rice phenylalanine ammonia-lyase gene OsPAL4 is associated with broad spectrum disease resistance[J].Plant Molecular Biology,2015,87(3):273-286.
[25] 马俊彦,杨汝德,敖利刚.植物苯丙氨酸解氨酶的生物学研究进展[J].现代食品科技,2007,23(7):71-74.
[26] FRASER C M,CHAPPLE C.The phenylpropanoid pathway in Arabidopsis[M].The Arabidopsis Book,2011.
[27] 郭鹏飞,雷健,罗佳佳,等.柱花草苯丙氨酸解氨酶(SgPALs)对生物胁迫与非生物胁迫的响应[J].热带作物学报,2019,40(9):1742-1751.
[28] RITTER H,SCHULZ G E.Structural basis for the entrance into the phenylpropanoid metabolism catalyzed by phenylalanine ammonia-lyase[J].The Plant Cell,2004,16(12):3426-3436.
[29] SHETTY N P,KRISTENSEN B K,Newman M A,et al.Association of hydrogen peroxide with restriction of Septoria tritici in resistant wheat[J].Physiological and Molecular Plant Pathology,2003,62(6):333-346.
[30] BOECKX T,WINTERS A L,WEBB K J,et al.Polyphenol oxidase in leaves:is there any significance to the chloroplastic localization[J].Journal of Experimental Botany,2015,66(12):3571-3579.
[31] 李颂,段玉玺,朱晓峰,等.黑曲霉次生代谢产物对番茄抗根结线虫病效果的影响[J].中国蔬菜,2011(4):44-49.
[32] APEL K,HIRT H.Reactive oxygen species:metabolism,oxidative stress,and signal transduction[J].Annual Review of Plant Biology,2004,55:373-399.
[33] 陳洁敏,韩效国.孢囊线虫对大豆生理生化特性的影响[J].莱阳农学院学报,1996(2):8-11.
[34] 杨宇虹,秦西云,刘敬业.过氧化物酶与烟草南方根结线虫病的抗性研究[J].烟草科技,1997(3):46-47.
[35] 陈增齐,段玉玺,陈立杰,等.球孢白僵菌Snf907发酵液对番茄根结线虫病的控病效果及防御酶活性影响[J].农药,2009,48(4):304-306.
猜你喜欢
家教世界·V家长(2020年5期)2020-06-29
红领巾·探索(2020年2期)2020-05-19
食品界(2019年8期)2019-10-07
作文大王·笑话大王(2018年12期)2018-03-23
湖北农业科学(2017年5期)2017-04-12
科技与创新(2016年22期)2017-03-30
山东农业科学(2017年3期)2017-03-29
创新作文(5-6年级)(2015年9期)2015-10-21
江苏农业科学(2015年9期)2015-10-20
湖南大学学报·自然科学版(2015年6期)2015-07-20
