
外源性神经激肽B对雌性大鼠生殖轴GnRH和Kisspeptin表达的影响
2021-01-15 04:48李海玲司文宇康铁柱姚郅璆李福宝方富贵
中国农业大学学报 2021年1期
李海玲 司文宇 宋 爽 康铁柱 姚郅璆 叶 菁 李福宝 方富贵
(安徽农业大学 动物科技学院,合肥 230036)
中枢神经是通过下丘脑-垂体-性腺(Hypothalamo-pituitary-gonadal,HPG) 轴来调控性腺,其信息传递取决于3个主要来源的神经和激素信号之间的动态相互作用:下丘脑前部,促性腺激素释放激素(Gonadotropin-releasing hormone, GnRH)合成并以脉冲形式分泌;垂体前叶,GnRH脉冲刺激垂体分泌促性腺激素;以及性腺,通过分泌性类固醇并产生配子响应促性腺激素的营养作用[1]。神经激肽B(Neurokinin B,NKB)是速激肽家族的一员[1],其主要通过神经激肽-3受体(NK3R)发挥生物学作用。NKB神经元及其受体主要分布于动物的下丘脑中[2-3],如弓状核(Arcuate nucleus,ARC)、正中隆起(Median eminence,ME)和视前区(Preoptic area,POA)等,此外在卵巢、胎盘、睾丸及前列腺中也均能检测到NKB及其受体NK3R分布[4-5]。
Kiss1是开启和控制性成熟的关键基因,Kisspeptin由Kiss1基因编码表达,主要与 kiss1 受体结合发挥作用,研究表明Kisspeptin/Kiss1R信号传导是GnRH和促性腺激素分泌的主要刺激因素[6]。研究发现NKB调节女性的GnRH和促黄体生成素(Luteinizing hormone,LH)分泌[7],且主要依赖Kisspeptin调节雌激素对LH分泌的反馈来诱导卵母细胞成熟,进而促进卵泡发育[8]。Navarro等[9]发现施用于大鼠侧脑室的NKB激动剂Senktide显著增加血清中LH的水平,并且在ARC中表达c-fos的Kiss1神经元数量增加10倍。同时NKB和Kisspeptin神经元均表达 NK3R,因此NKB可能作用于Kisspeptin神经元影响GnRH分泌[10]。NKB自发现以来,研究者们发现其在调控生殖的中枢机制中起着关键性的作用,不仅可作用于 GnRH 神经元影响 GnRH 的分泌,进而影响卵泡刺激素(Follicle stimulating hormone,FSH)和 LH 的分泌[12],而且还能与 Kisspeptin 相互作用,共同调节生殖内分泌系统[7]。有关NKB 在大鼠、小鼠、山羊和猴子等动物生殖轴上分布的报道多数仅涉及生殖轴中的某一个或某几个部分的组织、器官,而NKB对大鼠整个生殖轴上 NKB、GnRH 和 Kisspeptin 的分布及其表达量的影响报道甚少。因此,本研究通过侧脑室外源注射NKB探讨NKB对雌性大鼠生殖轴GnRH和Kisspeptin表达的影响,以期为NKB调节雌性动物生殖功能提供一定的科学依据。
1 材料与方法
1.1 试验材料
40只60 d的普通级Sprague-Dawley(SD)雌性大鼠,购自清源实验动物公司。
NKB购自Anaspec公司,批号23026;Trizol试剂和EnergicScript®FirstStrandcDNA合成试剂盒,购自上海闪晶分子生物科技有限公司;SYBR®Green Master Mix试剂盒,购自南京诺唯赞生物科技有限公司;小鼠源GnRH单克隆抗体(批号sc-32292)、驴抗小鼠IgG-FITC二抗(批号sc-2099)、山羊抗兔IgG-R二抗(批号sc-2780),购自上海优宁维生物科技有限公司;兔源Kisspeptin初级多克隆抗体和4′,6-二脒基-2-苯基吲哚,购自Abcam(上海)贸易有限公司。
1.2 试验设计
将大鼠随机分为试验组(n=20)和对照组(n=20)。试验组的大鼠侧脑室注射10 μL(600 pmol)[11]溶解于生理盐水的NKB溶液,对照组大鼠侧脑室注射等体积的生理盐水。所有大鼠均在侧脑室后20 min处死。每组10只大鼠断颈处死,快速分离出完整的下丘脑并冻存于-80 ℃冰箱,用于GnRH和Kiss1mRNA表达量的检测;每组另外10只大鼠从心脏的心尖处插入针头,用止血钳固定后剪破右心耳,通过左心室灌注生理盐水直至血水变清,然后灌注4%的多聚甲醛直至尾巴变硬,取下丘脑、垂体和卵巢固定于4%的多聚甲醛中,用于GnRH和Kisspeptin蛋白的免疫荧光检测。
1.3 侧脑室注射
大鼠麻醉后固定于脑室立体定位仪(BW-SDA903)上,头颅顶部剪毛消毒后,用手术刀沿皮肤中线切开表皮暴露出前囱的位置。以前囱为中心原点,距前卤点后方1.5 mm,左侧1.0~1.5 mm的位置进行定位,随后用牙科钻在定位点钻孔,将吸有不同药物的微量进样针固定于定位仪上并沿此孔垂直下针8.6~9.0 mm。药物注射完毕后停留5~10 s后再缓慢将微量进样针退出。拔针后缝合切口并进行消炎处理。术中注意观察大鼠生命特征,做好保暖措施。
1.4 下丘脑GnRH和Kiss1 mRNA转录水平测定
根据说明书,使用Trizol试剂提取下丘脑总RNA。NanoDrop分光光度计在260 nm处定量总RNA。通过甲醛凝胶电泳评估总RNA的质量,选取质量合格的RNA样品用于逆转录。使用EnergicScript®FirstStrandcDNA合成试剂盒,根据说明书的要求用1 μg RNA在20 μl的总反应体积中进行逆转录。
使用Primer Premier 5.0和AlleleID 6.0软件,根据GenBank数据库中GnRH,Kiss1和β-actin基因的核苷酸序列(GenBank登录号分别为:NM_012767.2,NM_181692.1和NM_031144.3)设计引物。引物由生工生物工程(上海)股份有限公司合成,其信息见表1。

表1 RT-qPCR引物信息Table 1 RT-qPCR primers information
RT-qPCR反应体系:2 μL的cDNA模板,10.4 μL 的SYBR®Green Master Mix混合物,0.4 μL的正向引物和0.4 μL的反向引物以及6.8 μL的双蒸水(ddH2O)。RT-qPCR条件:首先在95 ℃下预变性5 min,然后在95 ℃ 10 s和60 ℃ 30 s的40个循环中进行扩增,最后在95 ℃下15 s,60 ℃ 1 min 和95 ℃ 15 s来确定溶解曲线。通过 2-ΔΔCt法对基因相对表达量进行统计学分析。
1.5 生殖轴中GnRH和Kisspeptin的免疫荧光共定位
将4%多聚甲醛中固定的下丘脑、垂体和卵巢取出放于质量浓度300 g/L蔗糖的磷酸盐缓冲盐水(phosphate-buffered saline,PBS)中脱水。次日取出组织用滤纸吸取多余的水分后,将OTC包埋剂(Tissue OCT-Freeze Medium,OTC)包埋组织,随后放入冰冻切片机内加速冻凝。调整下丘脑切片厚度为7 μm,垂体和卵巢切片厚度为5 μm,收集完整的切片于防冻溶液中并储存在-20 ℃直至用于免疫荧光。
将下丘脑切片与小鼠源GnRH单克隆抗体(1∶500 稀释)和兔抗大鼠Kisspeptin多克隆抗体(1∶500 稀释)在4 ℃下孵育过夜,垂体和卵巢切片仅与兔抗大鼠Kisspeptin初级多克隆抗体(1∶500稀释)在4 ℃下孵育过夜。次日,切片在室温下用PBS洗3次(10 min/次)后用驴抗小鼠IgG-FITC(1∶ 200稀释)和山羊抗兔IgG-R(1∶200稀释)一起孵育2 h。然后室温下用PBS洗3次(10 min/次)并加入4′,6-二脒基-2-苯基吲哚(DAPI;蓝色核染色),温育10 min。最后用50%甘油封片固定。为验证本次试验没有非特异性染色,将切片同时设置阴性对照,阴性对照用PBS代替鼠GnRH单克隆抗体或兔抗大鼠Kisspeptin初级多克隆抗体。
1.6 统计分析
分别取试验组和对照组封片后的组织切片,在荧光显微镜下选用绿光(FITC)、红光(R)、蓝光(DAPI)等荧光通道进行观察、拍照,然后用Image-Pro Plus 6.0软件对荧光分布图片进行半定量荧光强度分析,得到其平均荧光强度值。用 Image J软件统计下丘脑不同核团上单位面积GnRH和Kisspeptin阳性细胞的数量。
试验数据采用配对样本T检验进行分析,所有数值表示为平均值±标准差(Mean±SEM),运用SPSS Statistics软件进行统计学处理和相关性分析。P<0.05表示显著性差异。
2 结果与分析
2.1 NKB对大鼠下丘脑GnRH和Kiss1 mRNA转录水平的影响
由图1可以看出,与对照组相比,侧脑室注射NKB组的GnRH的mRNA表达水平显著升高(P<0.05),Kiss1mRNA水平显著降低(P<0.05)。
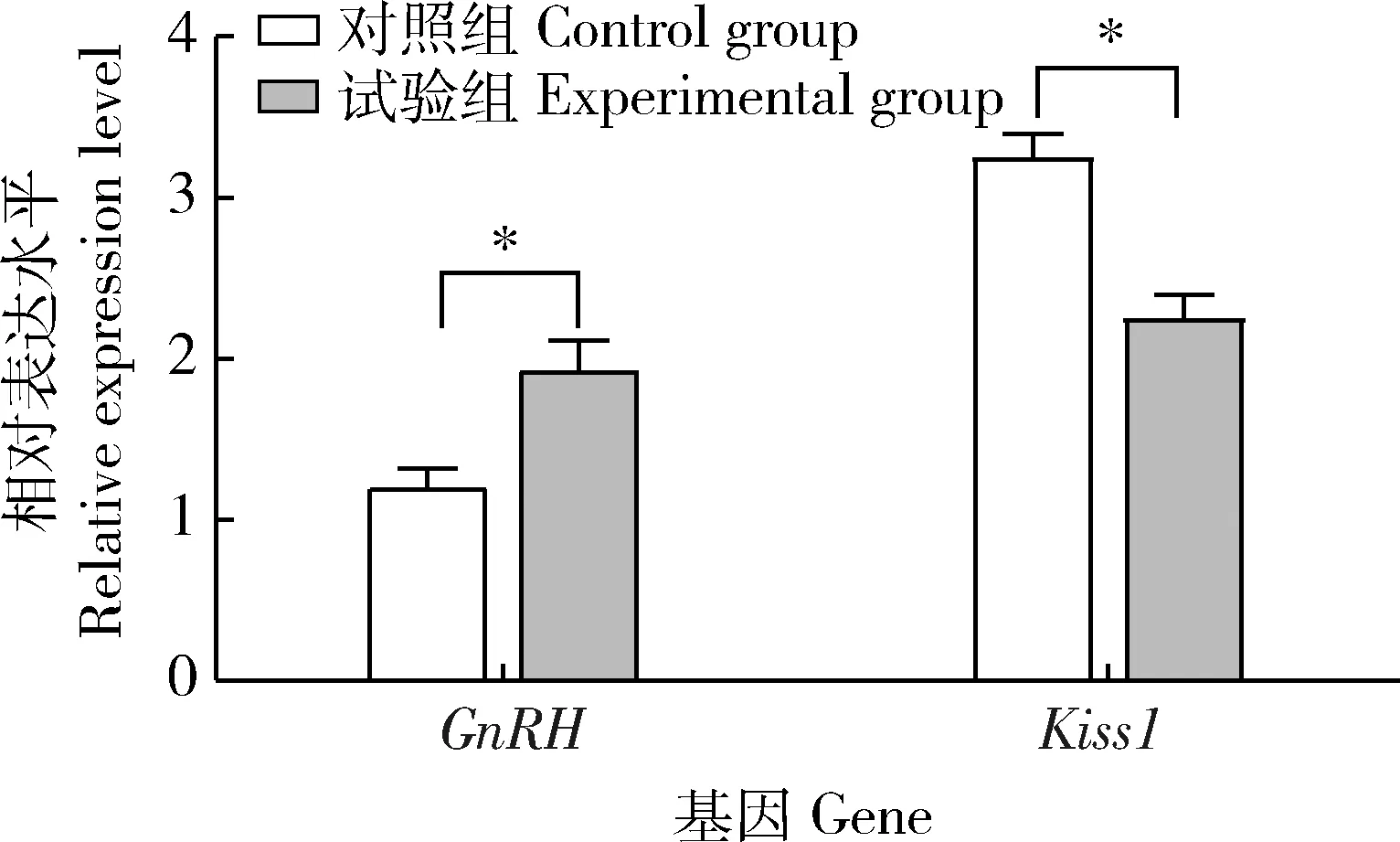
*表示显著差异(P<0.05)。下同。* indicate significant differences (P<0.05). The same as below.图1 NKB对大鼠下丘脑GnRH和Kiss1 mRNA转录水平的影响Fig.1 Effect of NKB on GnRH and Kiss1 mRNA transcription level in rat hypothalamus
2.2 NKB对大鼠下丘脑GnRH和Kisspeptin分布的影响
试验组中,GnRH在ARC、POA、ME和室旁核(Paraventricular nucleus,PVN)存在不同强度的免疫荧光表达,其中在ARC处呈现密集的广泛分布的强荧光表达,而在POA、ME和PVN处为分散的点状较弱荧光;Kisspeptin在ARC、POA、ME和前腹侧室旁核(Anterior ventral paraventricular nucleus,AVPV)有不同强度的免疫荧光表达,其中在ARC、ME和AVPV处为广泛而密集的强荧光表达,而在POA处为稀疏的较弱荧光(图2(a));对照组中,GnRH仅在ARC和ME中存在广泛分布的强荧光表达,Kisspeptin在ARC和AVPV中存在免疫荧光表达(图2(b))。对下丘脑不同核团中的GnRH和Kisspeptin阳性细胞数进行统计(图3和图4),与对照组相比,试验组中GnRH阳性细胞数量在ARC,POA和ME中增加,但仅在POA核团中呈现显著增加(P<0.05)(图3)。但试验组的Kisspeptin阳性细胞数在ARC、PVN、AVPV和POA中均显著低于对照组(P<0.05)(图4)。
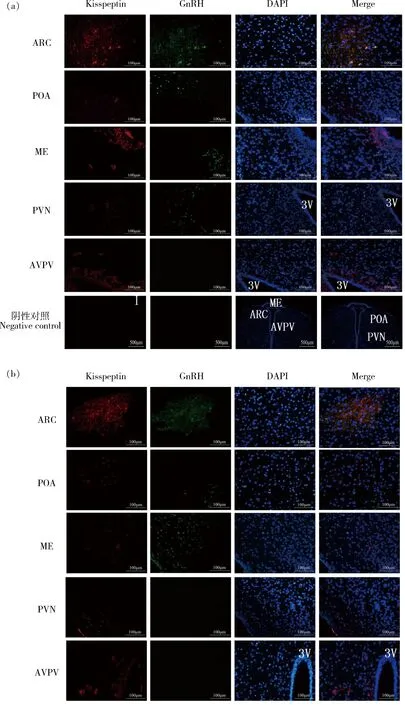
ARC:弓状核; POA:视前区; ME:正中隆起;PVN:室旁核;AVPV:前腹侧室旁核;阴性对照:用PBS代替鼠GnRH单克隆抗体或兔抗大鼠Kisspeptin初级多克隆抗体;3V:第三脑室。ARC:Arcuate nucleus; POA: Preoptic area; ME: Median eminence; PVN: Paraventricular nucleus; AVPV: Anterior ventral paraventricular nucleus; Negative control; PBS instead of GnRH MAb; 3V: The third ventricle。图2 NKB对试验组(a)和对照组(b)大鼠下丘脑GnRH和Kisspeptin分布的影响Fig.2 Effects of NKB on the distribution of GnRH and Kisspeptin in rat hypothalamus for experimental and control group
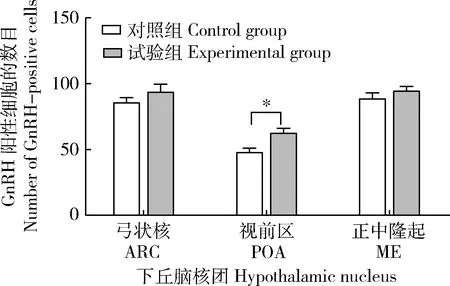
图3 NKB 对大鼠下丘脑弓状核(ARC),视前区(POA)和正中隆起(ME)GnRH阳性细胞数的影响Fig.3 The effect of NKB on the number of GnRH-positive cells in the arcuate nucleus (ARC), preoptic area (POA) and median eminence (ME) of hypothalamus in rats
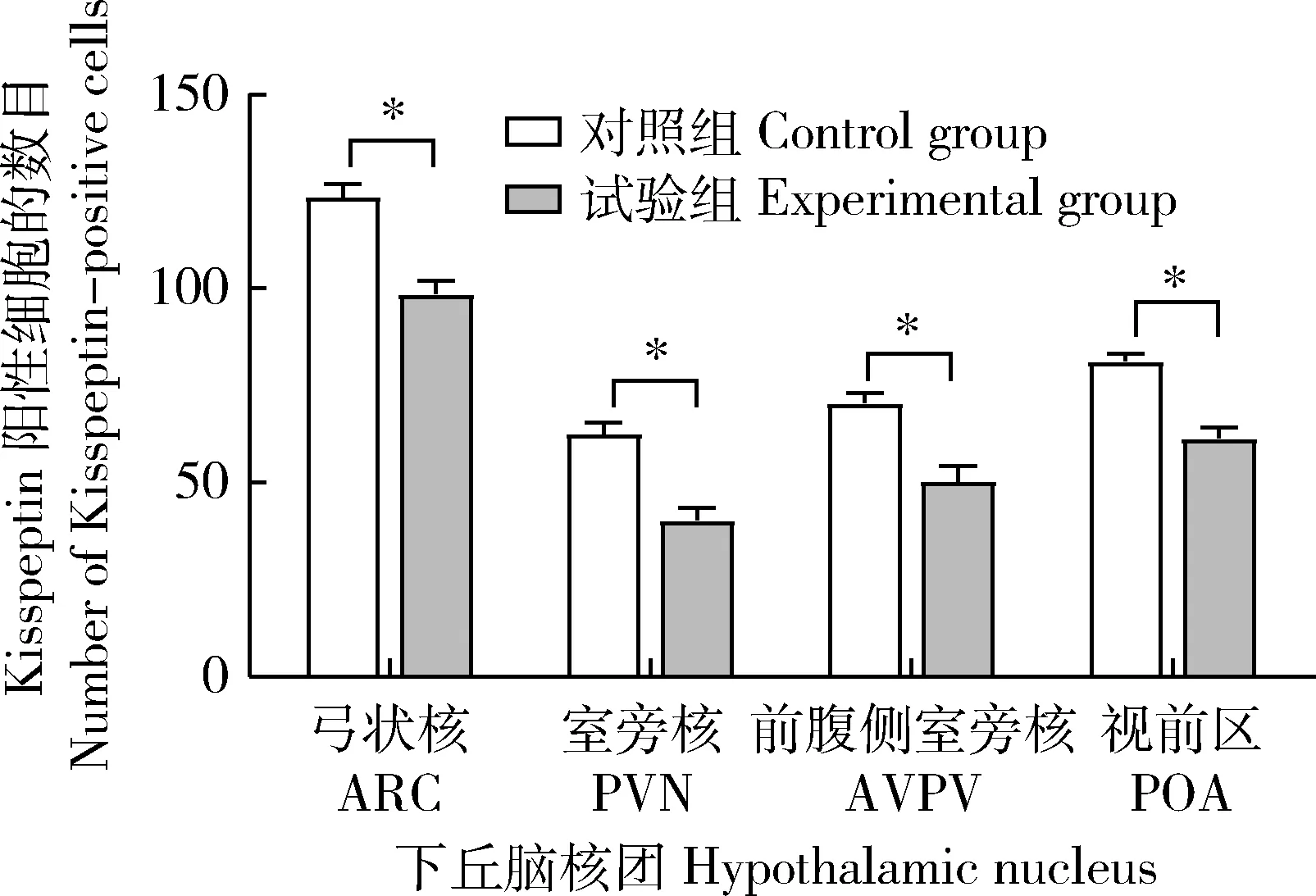
图4 NKB对大鼠下丘脑弓状核(ARC),室旁核(PVN),前腹侧室旁核(AVPV)和视前区(POA)Kisspeptin阳性细胞数的影响Fig.4 The effect of NKB on the number of Kisspeptin-positive cells in the arcuate nucleus (ARC),paraventricular nucleus (PVN), anterior ventral paraventricular nucleus (AVPV) and preoptic area (POA) of hypothalamus in rats
2.3 NKB对大鼠垂体和卵巢Kisspeptin分布的影响
Kisspeptin主要分布在成年雌性大鼠的腺垂体(图5),且试验组Kisspeptin平均荧光强度显著低于对照组(P<0.05)(表2);在成年大鼠的卵巢上也存在有Kisspeptin的荧光表达(图6),主要分布于卵巢间质和卵泡膜细胞上,但2组之间没有显著性的差异(P>0.05)。
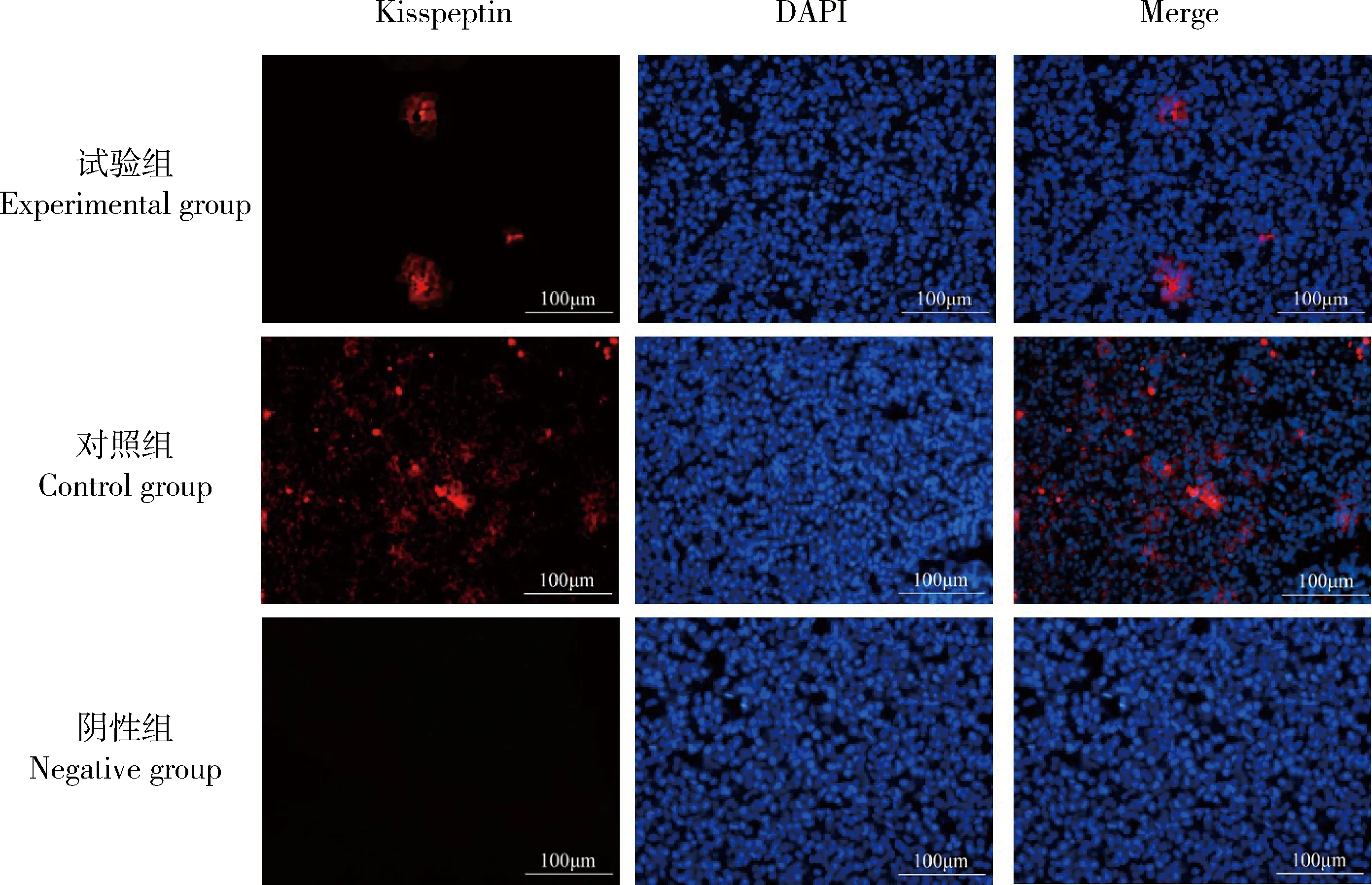
图5 NKB对大鼠腺垂体Kisspeptin分布的影响Fig.5 Effect of NKB on Kisspeptin distribution in rat adenohypophysis
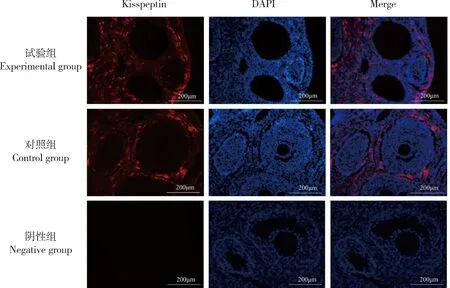
图6 NKB对大鼠卵巢Kisspeptin分布的影响Fig.6 Effect of NKB on the distribution of Kisspeptin in rat ovary

表2 NKB对大鼠垂体和卵巢上Kisspeptin表达的平均荧光强度的影响Table 2 The effect of NKB on the mean fluorescence intensity of Kisspeptin in the pituitary and ovary of rats
3 讨 论
中枢神经通过下丘脑-垂体-性腺轴实现对性腺的调控,生殖轴的信息传递路线即GnRH神经元接受其上游机制的调控后,脉冲性释放GnRH,其作用于垂体,调节促性腺激素的释放,促性腺激素又作用于性腺,调节生殖功能及性激素的分泌[10]。下丘脑是动物中枢神经系统中的重要器官,可以维持正常代谢并调控动物的生殖活动。下丘脑GnRH的脉冲式分泌是HPG轴功能正常启动和维持的关键[9]。近年来,研究者对NKB、Kisspeptin及其受体在人、猕猴、猪、大小鼠等多种动物下丘脑定位分布已做了大量的研究。目前大多数研究者认为 NKB 和 Kisspeptin 主要共表达于弓状核[12]。本研究免疫荧光结果显示,Kisspeptin在ARC、PVN、AVPV和POA中均有不同强度的荧光表达,且与对照组相比,试验组ARC、PVN、AVPV及POA中Kisspeptin阳性细胞数显著降低。这与不同生理时期大鼠体内下丘脑Kisspeptin免疫荧光分布表达结果相似[13]。前期研究表明,给雄鼠同时注射NKB和Kisspeptin,其LH分泌量高于单独注射Kisspeptin[14],另外研究者在动物和人类中均发现,NKB 神经元同时表达Kisspeptin和强啡肽(Dynorphin,Dyn)神经元,而弓状核的 Kiss-1、Tac2能够增加 LH 的分泌,表明NKB可以增强Kisspeptin在生殖发育的促进作用。小鼠[15]和山羊[16]侧脑室注射NKB激动剂后Kiss1mRNA表达量和LH脉冲释放增加,提示TAC3可能为Kiss1的上游基因,通过直接或间接作用调控Kisspeptin神经元从而调控生殖功能。本研究中,试验组大鼠下丘脑的Kiss1 mRNA表达量与对照组相比显著降低,该结果与小鼠[15]和山羊[16]侧脑室内注射NKB激动剂后Kiss1mRNA表达量增加不一致,表明不同物种可能会对NKB对Kiss1的表达产生影响。与对照组相比,试验组GnRHmRNA的转录水平增加,Kiss1mRNA转录水平却显著降低,该结果与注射Kisspeptin促进GnRH的结论[17]相反。对于这种差异,可能的原因:一是可能与样品来源有关,因为GnRH与Kisspeptin在下丘脑不同核团分布不同,本试验中样品为整个下丘脑,并非下丘脑某个核团,导致GnRH与Kisspeptin的表达不一致;二是因为Kisspeptin是GnRH上游调控因子[18],其基因转录水平未必同时增加,可能Kisspeptin增加在GnRH之前;三是NKB还可能直接或者间接通过其他通路如Dyn等影响GnRH释放,ARC中的单个细胞群(在绵羊中)产生3种神经肽:Kisspeptin,Dyn和NKB,并且这3 种神经肽都与GnRH分泌相关[19]。因此,NKB调节Kisspeptin和GnRH的确切机制仍需进一步探究。
垂体包括神经垂体和腺垂体,其中腺垂体是内分泌系统的重要调控枢纽,受下丘脑释放激素和靶腺激素的双重调节[20]。本研究表明,外源性给予NKB可使腺垂体中Kisspeptin高表达。这与在雌性恒河猴垂体上Kisspeptin的定位研究结果一致,即Kisspeptin主要存在于雌性恒河猴的垂体前叶及中叶[7]。本研究中侧脑室注射NKB导致腺垂体Kisspeptin表达的变化,可能是通过影响下丘脑GnRH和Kisspeptin的表达来实现的。有研究表明下丘脑ARC上的NKB通过传出神经元进入垂体门脉循环影响垂体前叶LH和FSH释放[21-22]。Billings等[23]在母羊上发现NKB受体激动剂可以促进LH分泌,且Navarro等[11]研究发现NKB受体激动剂对雌性大鼠LH分泌有持续性的兴奋作用。也有研究表明NKB通过脑垂体的GH3细胞系促进催乳素的分泌[24]。提示NKB可以通过腺垂体参与生殖轴的调控作用,也可以直接作用于脑垂体,影响促性腺激素的分泌,进而调控生殖。
雌性动物卵巢的功能受下丘脑腺垂体系统的调节。本试验侧脑室注射NKB,不仅影响了大鼠下丘脑和垂体Kisspeptin的表达,还发现试验组卵巢的卵泡膜、颗粒细胞和间质细胞Kisspeptin的荧光强度高于对照组。该结果与性腺切除小鼠侧脑室注射NKB激动剂后Kiss1mRNA表达增加相似[25-26],即NKB增加Kiss1的含量进而促进Kisspeptin的表达。已有的研究表明雌性大鼠卵巢上具有Kisspeptin分布,并且在排卵前Kiss1 mRNA的表达增加,表明Kisspeptin可能参与卵泡形成、黄体生成和排卵的过程[27]。Patterson等[28]发现先天性促性腺激素缺乏症(Isolated hypogonatropic hypogoadism,IHH)患者都存在GPR54 基因的突变,发现给GPR54基因缺陷的动物模型注射GnRH后可以缓解停滞的性成熟症状,由此认为启动初情期的关键因子之一是GPR54,Kisspeptin-GPR54 系统在青春期发育过程中发挥着重要的作用。Seminara等[29]发现,在未成熟雌性小鼠脑室内长期注入 Kisspeptin 可引起生殖轴的提前发育,如阴门的提前开放、卵巢和子宫重量增加,这可能是由于它的内源性受体GPR54信号与GnRH所释放的激活信号偶联的缘故,而Kisspeptin很有可能是通过激活GnRH来实现对生殖轴的调节作用。本试验中也可能是NKB直接调节下丘脑GnRH和Kisspeptin,进而影响垂体的分泌来调控卵巢的功能。已知雄激素由卵泡膜细胞和间质细胞合成,并通过卵泡细胞或颗粒细胞最终转化为雌激素,与此同时颗粒细胞还可以产生孕酮[30],提示NKB可能通过颗粒细胞、卵泡膜细胞和间质细胞调控性激素分泌,从而对生殖轴产生作用,调控雌性大鼠的生殖活动。
猜你喜欢
世界科学技术-中医药现代化(2021年5期)2021-11-05
中国妇幼健康研究(2019年2期)2019-03-26
现代园艺(2018年2期)2018-03-15
中老年健康(2017年9期)2017-12-13
中成药(2017年6期)2017-06-13
中国民族医药杂志(2016年7期)2016-05-09
湖南中医药大学学报(2015年1期)2016-01-06
磁共振成像(2015年1期)2015-12-23
郑州大学学报(医学版)(2015年2期)2015-02-27
中学科技(2014年11期)2014-12-25
