
放流三疣梭子蟹月间回捕率及繁殖贡献率研究
2021-01-28 10:34刘滨玮姜洁明翁歆之杨颜溢伍明鸣刘文雷王子维王连顺秦玉雪
水产科学 2021年1期
刘滨玮,刘 帅,姜洁明,翁歆之,杨颜溢,伍明鸣,刘文雷,王子维,王连顺,刘 奇,秦玉雪
(1.大连海洋大学 水产与生命学院,辽宁 大连 116023; 2.设施渔业教育部重点实验室,辽宁 大连 116023; 3.台州市水产技术推广总站,浙江 台州 318000 )
三疣梭子蟹(Portunustrituberculatus)俗称梭子蟹、飞蟹,属甲壳纲、十足目、梭子蟹科、梭子蟹属,为大型经济蟹类,生理生命可达2~3年,但3龄者较少,雄性最大的个体体质量可达710 g;雌性个体体质量可达730 g[1]。广泛分布于西北太平洋沿岸海域,包括日本、韩国、朝鲜、中国、越南及马来西亚沿海[2-3]。因肉质鲜美,口感细腻,营养丰富,1981年开始被列为我国水产养殖的对象[4-8]。1986—1988年我国营口地区首次开展人工放流,放流C2期稚蟹共60余万只[9]。
辽东湾位于渤海东北部,是一个半封闭的浅海湾,水交换能力较弱,水循环周期约为15 a[10]。沿岸有辽河、大凌河、六股河等多条河流汇入[11],带来了大量营养盐,湾内初级生产力较高,饵料生物充足,具有河口地区典型的生境特征,曾是包括三疣梭子蟹在内多种经济动物的重要索饵场和产卵场[12-13]。三疣梭子蟹属杂食性底栖动物,在生态系统中扮演着调节饵料生物、小型动植物密度,以及提高水域资源价值的关键生态角色,是开展资源增殖与养护的适宜种类[14]。伴随20世纪90年代辽东湾沿岸经济崛起[15],三疣梭子蟹渔业资源迅速衰退,在2004—2008年,辽东湾盘锦海域难以形成有效的梭子蟹渔汛,故依靠天然群体的自然繁殖来恢复种群规模并不现实[16-17]。为恢复梭子蟹渔业资源量,保证渔业生产的可持续发展和维持辽东湾生态系统平衡,自2012年开始,辽东湾海域以每年4—5月间海捕受精的雌性亲蟹作为产卵群体,人工孵化出F1代群体,开展规模化增殖放流。2015年,全省共放流C2期稚蟹1000余万只。目前,三疣梭子蟹已成为辽宁省第二大规模人工增殖种类。
近年来,在我国沿海地区广泛开展了三疣梭子蟹放流工作,取得了一定经济效果。然而截至目前,对三疣梭子蟹增殖放流缺乏有效监控和定量分析。笔者利用6对高多态性微卫星DNA分子标记,对辽东湾盘锦海域三疣梭子蟹增殖放流开展大规模追踪调查,以期评估放流后短期时间内各月间回捕到的放流蟹占当月回捕蟹群体的比例(月间回捕率)以及雌性亲蟹对回捕到的放流子代群体的贡献率程度(繁殖贡献率),为将来建立科学的放流体系,构建负责任增殖放流新模式提供基础资料。
1 材料与方法
1.1 三疣梭子蟹增殖放流
2015年增殖放流承担单位使用50只海捕已受精雌蟹作为繁殖亲蟹,经暂养、人工繁育等步骤,共产生约500万只C2期稚蟹,2015年6月,将上述稚蟹放流到盘锦沿岸海域。于当年7月28日—10月3日在盘锦沿岸以地笼网形式开展回捕调查,共获得1671只三疣梭子蟹样本,剪其螯足,剥离肌肉冷冻运回实验室,立刻用无水乙醇多次洗涤后长期保存。
1.2 放流子代三疣梭子蟹个体识别
1.2.1 DNA抽提
采用吸附柱法(TIANamp Marine Animals DNA Kit,中国)从50只雌性繁殖亲蟹与1671只回捕蟹的螯足中提取DNA,共计对1721只三疣梭子蟹样本开展个体识别。同时采用含量为1.00%的琼脂糖凝胶电泳(DYY-8C型,中国)来检测DNA提取质量,使用核酸定量分析仪来检测DNA浓度(K2800,中国)。
1.2.2 微卫星引物
共使用6对具有高多态性的微卫星DNA分子标记开展试验。在预试验阶段,以上引物可靠性均经过野生群体验证。引物序列参考文献[18-21]的试验设计。每对引物均在5′端添加相应的荧光修饰接头(表1)。

表1 多态性微卫星位点的特征
1.2.3 聚合酶链式反应与毛细管电泳
聚合酶链式反应流程和体系参照文献[13],但每个聚合酶链式反应总体积设置为15 μL,为了保证检测的可靠性,使用上述6对引物对1721只三疣梭子蟹(50只亲本,1671只回捕个体)样本单独扩增。扩增产物经琼脂糖凝胶电泳检测合格后,使用ABI3730XL基因分析仪(Applied Biosystems,美国)基因测序仪开展毛细管电泳,并基于GeneMapper(4.0版本)软件绘制荧光信号峰图,最后进行人工校正。在进行毛细管电泳时,设置多组阳性、阴性和空白对照,保证所绘制的基因型数据的准确性和可重复性(图1)。
1.3 统计分析
1.3.1 亲子鉴定
首先使用Microchecker 2.2.3软件[22]检测基因型绘制错误和无效等位基因频率情况,采用亲缘关系分析软件Cervus 3.0.7[23]开展亲子鉴定及遗传多样性分析,所分析的遗传参数包括:等位基因数、观测杂合度、期望杂合度、多态信息含量、同时计算非亲排除率,分子标记的累积排除率,根据鉴定成功的位点数量计算鉴定准确率,最终计算出亲本对子代的贡献率。各亲本间的繁殖贡献率差异经SPSS 22.0中的非参数卡方检验,P<0.05认为差异显著。

图1 微卫星位点代表性分型结果Fig.1 Typical genotype of the microsatellite loci
1.3.2 各月间回捕到的放流蟹占当月回捕蟹群体的比例
2015年6月放流后,于7—10月在盘锦沿岸主要采用地笼网调查的方式开展回捕调查,调查地点见图2。并基于亲子鉴定结果,绘制出盘锦海域放流三疣梭子蟹月间回捕率变化图,查明各月间回捕到的放流三疣梭子蟹占当月回捕蟹群体的比例变化趋势。

图2 盘锦海域回捕调查范围Fig.2 Investigation area for recapture swimming crab in coastal Panjin area
1.3.3 雌性繁殖亲蟹与回捕蟹群体遗传多样性比较
比较雌性繁殖亲蟹(50只)与回捕蟹群体(1671只)的遗传多样性差异以探讨放流事件中雌性繁殖亲蟹的遗传多样性层次。使用Arlequin 3.5软件[24]对上述2个群体开展哈迪温伯格平衡检验;使用GenAlEx 6.503软件[25-26]分析等位基因数、有效等位基因数、观测杂合度、期望杂合度和无偏期望杂合度;使用HP Rare 1.0软件[27]分析等位基因丰度。多重检验经sequential Bonferroni校正显著性[28]。雌性繁殖亲蟹与回捕蟹群体间的遗传指标差异经SPSS 22.0中的配对样本t检验,P<0.05认为有统计学差异。
2 结 果
2.1 微卫星扩增片段分析
本试验所使用的6对高多态性微卫星引物在三疣梭子蟹群体中均能获得稳定的、高强度的毛细管检测信号(图1)。无效等位基因频率检测显示,所有位点均未发现明显的无效等位基因干扰(<0.02)(表2、表3),证实这些引物可以用于后续的遗传多样性分析与亲子鉴定。试验发现,6对微卫星引物共在2个群体中获得519个等位基因,其中在雌性繁殖亲蟹群体成功扩增出176个等位基因;在回捕蟹群体扩增出343个等位基因,此外等位基因出现最多的微卫星引物为Pot51,最少的为Ptri1。两群体中多态信息含量平均值均大于0.9,也印证了所使用的微卫星引物的高多态性。

表2 三疣梭子蟹雌性繁殖亲蟹微卫星DNA标记特征参数

表3 三疣梭子蟹回捕群体微卫星DNA标记特征参数
2.2 雌性繁殖亲蟹与回捕蟹群体遗传多样性分析
雌性繁殖亲蟹与回捕蟹两群体开展了遗传学参数分析,计算了等位基因、有效等位基因、等位基因丰度、多态信息含量、期望杂合度、无偏期望杂合度和观测杂合度等参数。以上参数在雌性繁殖亲蟹与回捕蟹群体中成正态分布(P>0.05,Shapiro-Wilk test,SPSS),故使用配对样本t检验雌性繁殖亲蟹与回捕蟹群体遗传多样性差异发现,除回捕蟹群体的等位基因(57.167)显著大于雌性繁殖亲蟹群体(29.333)外,其他遗传学参数在两群体间均无显著差异(P>0.05,SPSS),证实三疣梭子蟹增殖放流所使用的雌性繁殖亲蟹遗传多样性较高。同时就整体而言,雌性繁殖亲蟹群体尽管样本数量远不及回捕蟹群体,但基因频率符合哈迪温伯格平衡(P>0.05,SPSS),进一步证实所使用的雌性繁殖亲蟹群体种质较好(表4)。

表4 雌性繁殖亲蟹与回捕群体遗传参数显著性检验
2.3 亲子鉴定分析
因在三疣梭子蟹人工放流过程中,父本基因型未知,故使用Cervus软件中的母本鉴定开展亲子鉴定。亲子鉴定结果显示,当使用4个微卫星分子标记时,累积排除率为0.9979,亲子鉴定的准确率为84.17%;微卫星分子标记为5个时,累积排除率为0.9993,亲子鉴定的准确率为90.83%;微卫星分子标记为6个时,累积排除率为0.9997,亲子鉴定的准确率为95.83%(表5,图3),最终从回捕群体中共发现120只放流子代。
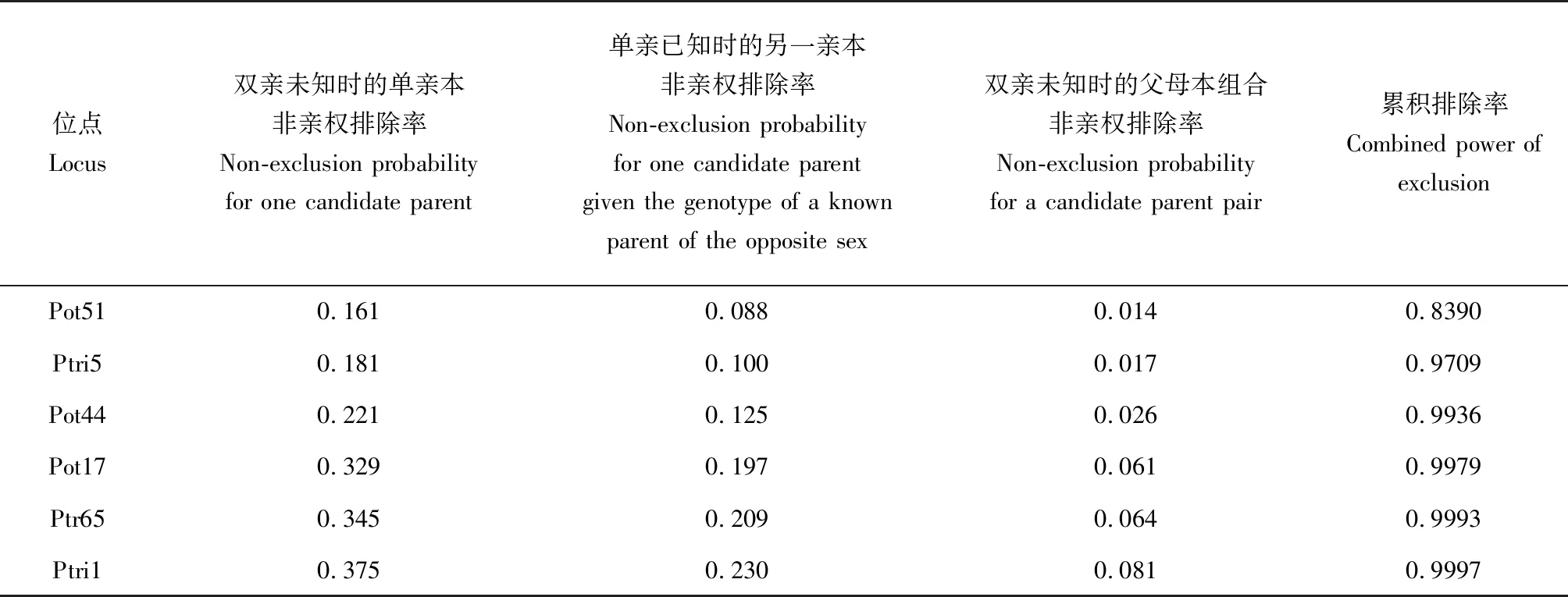
表5 亲权鉴定参数

图3 不同微卫星位点数的亲子鉴定准确率Fig.3 Identification accuracy of different microsatellites for parental authority
2.4 繁殖贡献率分析
本试验结果显示,在所检测的样本中,有22只雌性繁殖亲蟹在1671只回捕蟹群体中鉴定到其所产子代,占所检测的雌性繁殖亲蟹总样本的44%。但各雌性繁殖亲蟹对子代的繁殖贡献率存在显著差异(χ2=142.050,自由度=6,P<0.001,非参数卡方检验)。在鉴定到的120只回捕子代当中,有87只来自3只雌性繁殖亲蟹,其中亲蟹1贡献率为51.67%,亲蟹2贡献率为15.00%,亲蟹3贡献率为5.83%,其他亲蟹对子代的贡献率仅维持在0.83%~4.17%(图4)。
2.5 盘锦海域放流三疣梭子蟹月间回捕分析
本次三疣梭子蟹回捕调查历时4个月,其中7月共回捕到359只,基于分子标记,从回捕蟹群体中识别出当年放流群体为59只,即7月放流蟹占回捕蟹群体的比例为16.43%;8月共回捕到115只,从回捕群体中识别出当年放流群体为3只,即8月放流蟹占回捕蟹群体的比例为2.61%;9月共回捕到1167只,从回捕群体中识别出当年放流群体为56只,即9月放流蟹占回捕蟹群体的比例为4.80%;10月共回捕到30只,从回捕群体中识别出当年放流群体为2只,即10月放流蟹占回捕蟹群体的比例为6.67%(图5)。

图4 雌性亲蟹的繁殖贡献率Fig.4 The rate of reproduction contribution in female broodstock swimming crab P. trituberculatus
3 讨 论
本试验共使用6对高多态性微卫星引物,对2015年辽东湾盘锦海域三疣梭子蟹增殖放流开展回捕调查,通过对放流个体开展个体识别的方式解析了放流后短时间内回捕到的放流群体占调查中回捕群体的回捕率(月间回捕到的放流三疣梭子蟹占当月回捕蟹群体的比例)以及雌性亲蟹对放流子代群体的繁殖贡献率。

图5 月间回捕率曲线Fig.5 Monthly recapture rate curve
3.1 微卫星位点
本试验中,所有微卫星位点均表现出极高的多态性。以验证微卫星DNA标记的多态性信息含量为例,6个位点的多态性信息含量平均值均在0.9以上,显示出所应用的位点具有极佳的遗传信息丰富性[29]。在精确的亲子鉴定过程中,选择高多态性的微卫星位点是重要一环;此外微卫星位点在基因组中的随机分布性、扩增片段的稳定与可重复性,毛细管电泳的高强度信号也是成功鉴定的关键因素[30]。大量的前期试验发现[13,31],上述6个微卫星位点无疑具备以上特征,适宜用于三疣梭子蟹放流后的回捕追踪。
3.2 雌性繁殖亲蟹与回捕蟹群体遗传多样性
前期研究发现,增殖放流过程中亲本的种质质量是影响放流群体遗传多样性的关键因素[32-33]。如果使用养殖群体或者遗传多样性较低的海捕群体作为亲本来大量繁育放流群体,很可能造成放流群体遗传多样性的下降,甚至威胁当地固有群体的遗传多样性[34]。本次试验,因回捕蟹群体的数量远多于雌性繁殖亲蟹群体,造成回捕蟹群体的等位基因显著多于雌性繁殖亲蟹群体(P>0.05),是合理的。因在计算过程中已经考虑到了群体大小的差异性,解析如等位基因丰度等这些遗传参数,开展不同群体大小的基因多样性比较是可行的[27]。笔者发现,在有效等位基因、等位基因丰度、期望杂合度、无偏期望杂合度和观测杂合度这些遗传参数中,雌性繁殖亲蟹与回捕蟹两群体无显著差异(P>0.05)。鉴定结果显示,雌性繁殖亲蟹群体与当地回捕蟹群体的遗传多样性维持在较高水平,观测杂合度和期望杂合度均高于莱州湾和鸭绿江的野生群体[21,35]。盘锦地区放流用三疣梭子蟹雌性亲蟹具有较高的遗传多样性,有利于维持放流后当地固有群体的遗传多样性。然而每次增殖放流活动中亲蟹的使用数量也极大地影响放流后当地固有群体的遗传多样性与亲缘关系,尤其是甲壳类广泛存在有繁殖贡献率不平衡现象[36]。试验发现,在鉴定到的120只回捕子代当中,有87只来自3只雌性繁殖亲蟹,其中亲蟹1贡献率为51.67%,亲蟹2贡献率为15.00%,亲蟹3贡献率为5.83%,其他雌性亲蟹对子代的贡献率仅维持在0.83%~4.17%,且有28只繁殖雌蟹没有定位到对应子代。在以往的研究中,也检测了放流前雌性繁殖亲蟹对即将放流子代的繁殖贡献率,发现雌性亲本对子代均有贡献,最高为29.61%,最低为3.35%,不同亲本之间的贡献率也存在显著差异(P<0.05)[13]。三疣梭子蟹繁殖亲本间繁殖贡献率极不平衡这种情况,尽管目前没有太多直接证据,但是参阅其他水产种类的相关研究,尤其是甲壳动物,多数是由于广泛存在的多次交配,精子竞争,雌性性别选择所致[37-40]。在人工放流过程中繁殖雌蟹是人为选择的,这也可能会造成繁殖亲本间,繁殖贡献率不平衡的现象。因此在人工放流中,三疣梭子蟹这种繁殖贡献率极不平衡现象更需要使用较多的雌性亲蟹来参与繁育(图4)。目前使用较少的繁殖亲本繁育大量子代用于放流的现象在我国普遍存在,如盘锦海域的资源修复,故今后亟需在增殖放流标准中对亲本数量加以科学规划,以维持放流群体整体的遗传多样性[32-33,41]。此外在本次试验中,回捕群体中放流蟹识别比例不高,也可能会存在某些雌性亲蟹的对应子代样本采集不到的现象,从而一定程度上影响了雌性亲蟹繁殖贡献率。另一个值得注意的是,回捕群体不符合哈迪温伯格平衡,这可能由于盘锦当地常年的人工增殖放流(2012—2015年),极大地影响了三疣梭子蟹群体的自然选择,也改变了等位基因频率[42]。
3.3 亲权鉴定
微卫星标记因具有高多态性、基因组内随机分布性、重复检测稳定性等特征,是亲权关系鉴定中的常用遗传学检测方法[43]。

本试验采用最大似然法(LOD)在回捕群体中开展放流个体的谱系追踪,为了提高鉴定准确率,LOD值不仅大于0,且达到了95%的置信区间,以保证个体识别结果的可靠性,故最终从1671只回捕蟹中发现放流个体为120只,籍以此评估放流三疣梭子蟹的月间回捕率(各月间回捕到的放流蟹占当月回捕蟹总群体的比例)为2.61%~16.43%,表明人工放流三疣梭子蟹对当地群体产生了一定的补充和经济效益。本试验回捕率试验结果与蔡珊珊[32]所论述的在山东海域547只回捕蟹中,发现约15%为放流蟹的试验结果相近;但与谢周全等[48-50]的研究结果不同,其原因在于谢周全等[48-50]所述回捕率指的是通过对比增殖放流前后相对资源量变动来评估得到的放流群体对目标海区三疣梭子蟹资源的贡献率。
综上,本试验成果将深化对三疣梭子蟹增殖放流的认知,为这一物种开展科学的增殖放流提供有益指导。
4 结 论
本试验利用微卫星DNA分子标记技术跟踪了辽东湾盘锦海域三疣梭子蟹增殖放流过程,发现放流后苗种短期月间回捕到放流蟹占当月回捕蟹群体的比例为2.61%~16.43%。在鉴定到的120只回捕子代当中,有87只来自3只雌性繁殖亲蟹,其中亲蟹1贡献率为51.67%,亲蟹2贡献率为15.00%,亲蟹3贡献率为5.83%,其他亲蟹对子代的贡献率仅维持在0.83%~4.17%,故雌性繁殖亲蟹间存在明显的繁殖贡献率不平衡现象。
猜你喜欢
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
渔业科学进展(2022年2期)2022-04-11
水产科学(2022年2期)2022-03-20
南京师范大学学报(工程技术版)(2021年3期)2021-10-22
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
现代营销·经营版(2017年3期)2017-05-17
消费电子(2017年4期)2017-05-04
作文周刊·小学五年级版(2016年1期)2016-06-29
中学生英语·阅读与写作(2008年7期)2008-12-19
