
基于两个陆地棉低世代群体定位纤维品质相关QTL
2021-02-05 09:37郭晓豪王寒涛魏鑫张晶晶付小康马亮魏恒玲喻树迅
棉花学报 2021年1期
郭晓豪,王寒涛,魏鑫,张晶晶,付小康,马亮,魏恒玲,喻树迅*
(1. 河南科技学院,河南 新乡453000;2. 中国农业科学院棉花研究所/ 棉花生物学国家重点实验室,河南 安阳455000)
棉花是主要的天然纤维作物,每年世界棉花纤维消费量约为2700 万t[1],其中陆地棉是种植面积最广,具有产量高、适应性强的优点,占世界棉花产量的95%以上(http://www.cotton.org/)。近年来,纺织技术的不断革新和人民生活的不断提高对纤维品质提出了更高的要求,优质育种显得尤为重要[2-4]。 因此,在保持高产的基础上,培育高纤维品质的陆地棉品种是摆在育种家面前的重要课题。
纤维品质性状受微效多基因控制,是典型的数量性状,传统的表型选择在棉花的遗传改良中发挥了积极作用。 随着生物技术的发展,通过分子标记辅助育种可从分子水平上选择目标基因,进而实现目标性状的改良[5-9]。 在棉花纤维品质性状的QTL(Quantitative trait locus, 数量性状位点)定位上,前人已经开展了较多的研究:伊海法[10]采用优质母本和高产父本构建F2和F2:3群体,利用SSR(Simple sequence repeat,简单重复序列)标记进行QTL 定位,检测到35 个与纤维品质相关的QTLs;乔文青等[11]利用纤维品质差异显著的陆地棉亲本构建F2群体及F2:3群体,以已有的遗传图谱为基础定位到157 个与纤维品质有关的QTLs;Fang 等[12]利用陆地棉重组自交系群体构建分子标记连锁图,将纤维强度基因定位到62.6 kb 区间内。这些QTLs 的发现将为后续的育种工作提供重要的参考。
目前,关于纤维品质QTL 定位的报道不少,定位到的QTLs 很多, 但是借助这些QTLs 开展精细定位仍存在一定的困难,原因在于很少获得稳定的QTLs 区段。因此,仍然需要在精细定位上做补充, 以及就如何获得更多更稳定的QTLs 进行探索。
本研究利用陆地棉高强纤维品系中棉所679与农垦5 号构建F2和F2:3群体, 采用双亲间149对多态性SSR 引物对F2群体进行基因分型,根据基因型信息,使用QTL IciMapping 软件构建连锁图谱,并对纤维长度(Fiber length,FL)、长度整齐度指数 (Uniformity index,UI)、 断裂比强度(Breaking tenacity,BT)、 马克隆值(Micronaire,MIC)和断裂伸长率(Breaking elongation,BE)共5 个纤维品质性状进行QTL 定位。
1 材料与方法
1.1 亲本与群体构建
以高品质陆地棉品种中棉所679(简称CCRI 679)与低品质材料农垦5 号(Nongken 5,简 称NK-5)为亲本,于2016 年夏天在中国农业科学院棉花研究所老所部试验地(河南省安阳县)配制杂交组合, 同年冬天在海南自交加代,2017 年4月在老所部试验地播种亲本及F2群体。随机挑选200 个F2单株在现蕾期取幼叶样品,于-80 ℃冰箱保存。 7 月份开花期每个单株做20 个自交铃,10 月轧花收种保存。 2018 年4 月在东场部试验地种植F2:3家系,每个家系种1 行,2 个重复。 所有群体均设置4 m 行长,0.8 m 行距,0.25 m 株距。 田间管理与大田生产相同。
1.2 表型检测
F2自交铃按单株收获,F2:3群体每行随机挑选20 铃收获,并分别考种,纤维样品送原农业部棉花品质监督检验测试中心(河南安阳)检测纤维品质,包括前述的5 项指标。
1.3 基因型检测
双亲及F2单株的DNA 提取采用改良的CTAB(Cetyltrimethylammonium bromide,十六烷基三甲基溴化铵)法[13]。 利用6 688 对SSR 引物在双亲间进行多态性分析,共得到149 对清晰易辨的多态性引物,用这些多态性引物在F2群体间鉴定个体的基因型。 聚合酶链式反应(Polymerase chain reaction,PCR)体系与检测方法参照张军等[14]的方法。 F2与F2:3家系均基于F2基因型进行QTL 定位。
1.4 数据分析
使用IBM SPSS 21.0 软件对F2及F2:3群体表型信息进行描述性统计分析。 使用QTL IciMapping 4.1 软件[15]分析标记的偏分离情况、根据F2基因型数据绘制连锁图谱、根据图谱信息和表型数据进行QTL 定位。 在冗余标记分析中,设置缺失率为50%(缺失率高于50%的标记将被删除),偏分离的阈值设置为0.01(偏分离检验中阈值低于0.01 的标记将被删除), 选择删除BIN 标记(当多个标记的基因型信息完全一致时, 保留1个标记,删除冗余标记);连锁图谱构建中,设置连锁系数为4.0。 QTL 定位采用加性- 完备区间作图法(Inclusive composite interval mappingadditive mapping,ICIM-ADD), 设置似然比对数(Logarithm of odds,LOD)为2.5[16]。 F2:3群体定位以F2基因型为基础,以F2:3家系2 个重复表型的平均值(如无特殊说明,后文中对F2:3性状的描述与分析均指2 个重复的平均值)做QTL 定位[17-19],软件及方法与F2相同。
2 结果与分析
2.1 表型数据分析
F2及F2:3群体纤维品质性状的描述性统计见表1。 以均值分析,2 个群体在长度整齐度指数和马克隆值上无明显差异,而在纤维长度、断裂比强度和断裂伸长率上存在一定差异。 相比F2群体,F2:3群体纤维长度增加了1.14 mm, 增幅为4.02%;断裂比强度增加了2.16 cN·tex-1,增幅为7.06%;断裂伸长率降低了0.99 百分点。 F2:3群体在纤维长度、长度整齐度指数、断裂比强度、马克隆值的方差均小于F2群体, 表现出一定的稳定性。 为了进一步了解这些性状的分布情况,对2个群体的所有性状绘制了直方图(图1)。 在分布上,除了F2马克隆值与F2:3断裂伸长率,其余性状的偏度均在-0.5~0.5, 说明基本服从正态分布,F2马克隆值与F2:3断裂伸长率呈现一定程度的偏态分布,这可能与受到数量性状主效显性基因的影响有关。
2.2 连锁图谱构建
利用6 688 对SSR 引物鉴定双亲之间的多态性,共得到149 对多态性引物,多态性比例为2.23%。 通过对F2群体的基因型检测, 得到了200 个单株的149 个多态性位点的基因型数据。在绘制连锁图谱前, 使用QTL IciMapping 软件的BIN 功能,对这些标记做进一步筛选:淘汰基因型数据完全相同的重复标记和偏分离标记。通过BIN 功能,共删除11 个冗余标记,8 个偏分离标记。 以130 对多态性标记构建连锁图谱,最终得到包含119 个标记、28 个连锁群、 总长为1 173.5 cM(centiMorgan)的遗传连锁图谱。 其中,最大的连锁群全长为153.2 cM,包含14 个标记; 最小的连锁群长为3.3 cM, 包含2 个标记;平均每个连锁群包含4.25 个标记,标记间平均距离9.86 cM。
2.3 QTL 定位与分析
根据连锁图谱对F2和F2:3群体的5 个纤维品质性状做QTL 定位。 其中,F2群体共检测到9个QTLs,F2:3群 体 共 检 测 到11 个QTLs, 这 些QTLs 分布在11 个连锁群上(表2 和图2)。 根据Cotton FGD(http://www.cottonfgd.org/)网站收录的标记信息以及将标记序列与参考基因组对比的结果, 其中6 个连锁群分别锚定到A02、A05、A09、D04、D09 与D11 上;3 个连锁群确定了染色体编号但无法区分A、D 亚组:LG1 无法确定在A12 或D12 上,LG2 无法确定在A08 或D08上,LG3 无法确定在A10 或者D10 上;2 个连锁群LG4 与LG5 未能锚定到染色体。

表1 F2 及F2:3 群体纤维品质性状的描述性统计Table 1 Descriptive statistic characteristics of fiber quality traits in F2 and F2:3 populations
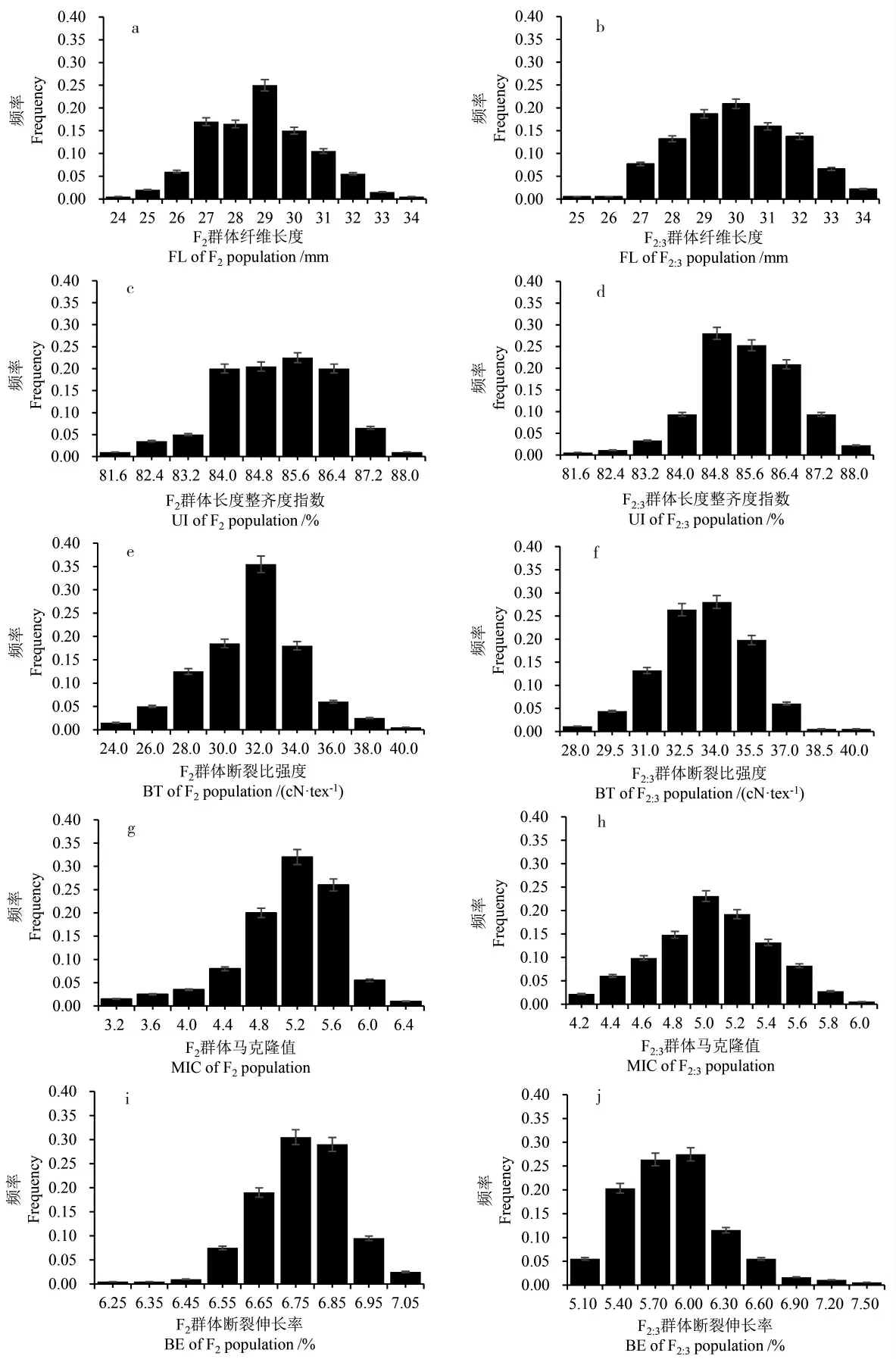
图1 F2 及F2:3 群体纤维品质性状的分布Fig. 1 The distribution of fiber quality traits in the F2 and F2:3 population
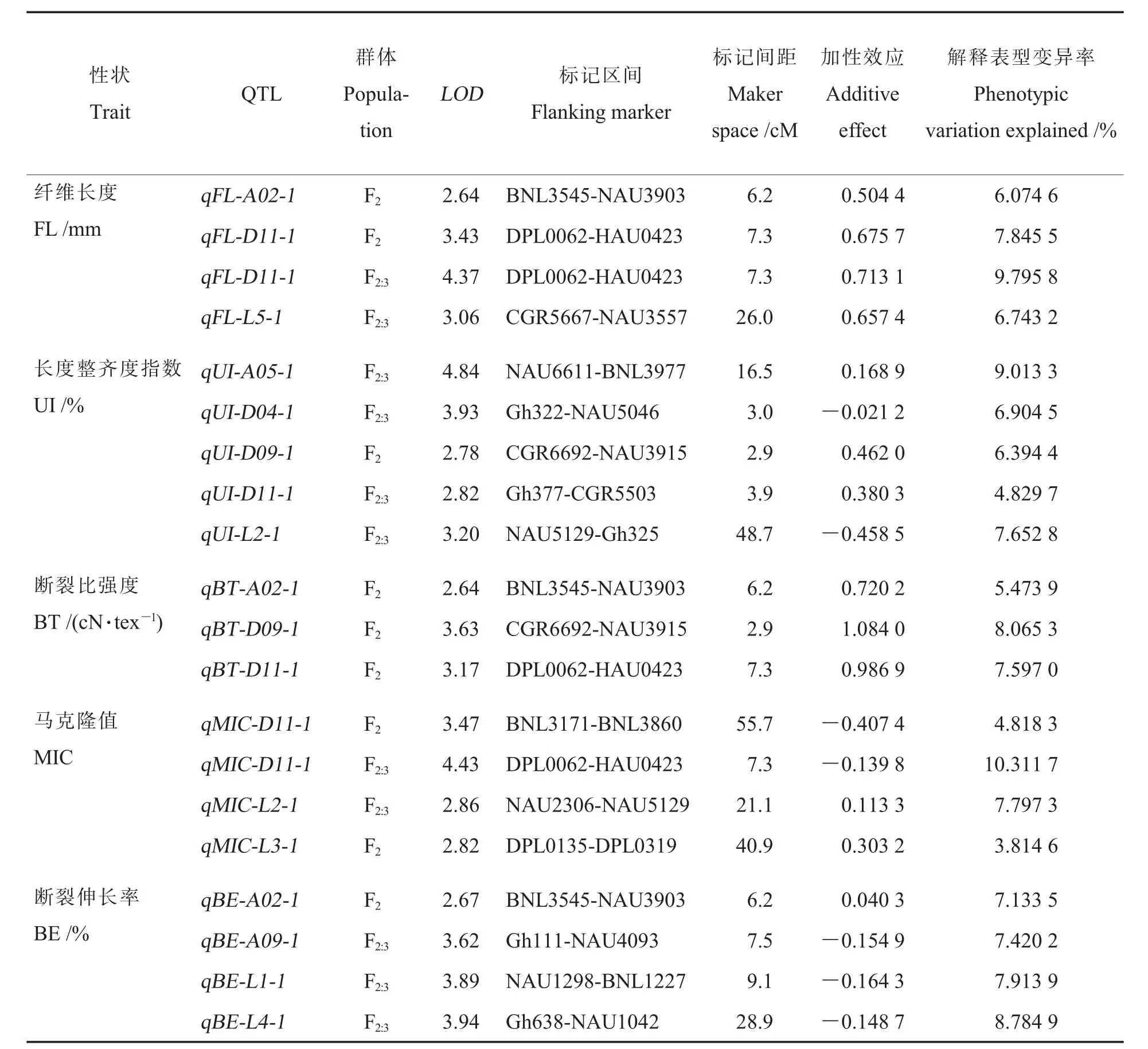
表2 F2 及F2:3 群体纤维品质相关QTL 的定位Table 2 Mapping of QTLs related to fiber quality in F2 and F2:3 populations
共检测到3 个与纤维长度相关的QTLs,分布在3 个连锁群,解释了6.074 6%~9.795 8%的表型变异。 这些QTLs 的增效基因均来自中棉所679, 这与中棉所679 作为高纤维长度亲本相吻合。其中,qFL-D11-1 在F2群体与F2:3群体中均能检测到,是本研究中仅有的同一性状在2 个群体中定位到同一区段的QTL。 除了纤维长度外,F2中断裂比强度相关QTL(qBT-D11-1)和F2:3中马克隆值相关QTL(qMIC-D11-1)也定位到了该区段。为了进一步了解该区段标记在D11 染色体上的物理位置信息, 对4 个紧密连锁的标记NAU3127、DPL0062、HAU0423、BNL3171 在参考基因组进行比对, 发现其位置顺序与遗传图谱相一致, 再次验证了该区段内标记遗传位置的可靠性。
共检测到5 个与长度整齐度指数相关的QTLs, 其中F2群体中检测到1 个,F2:3群体中检测到4 个。5 个QTLs 分布在5 个不同的连锁群,解释了4.829 7%~9.013 3%表型变异,qUI-A05-1解释表型变异率最大, 其加性效应值为0.168 9。在5 个QTLs 中,F2:3群 体 的 qUI-D04-1 和qUI-L2-1 的增效基因来自亲本农垦5 号,其余则来自中棉所679。

图2 连锁图谱构建与QTL 定位Fig. 2 Linkage map construction and QTL mapping
共检测到3 个与断裂比强度相关的QTLs,3个QTLs 均来自F2群体,分布在3 个不同的连锁群。 这些QTLs 解释了5.473 9%~8.065 3%的表型变异, 其中qBT-D09-1 解释的表型变异率最大,其加性效应值为1.084 0。 3 个断裂比强度相关的QTLs 的增效基因均来自于亲本中棉所679,这与中棉所679 作为高纤维强度亲本相吻合。
共检测到4 个与马克隆值相关的QTLs,其中2 个来自F2群体,2 个来自F2:3群体。 4 个QTLs 分布在3 个连锁群, 解释了3.814 6%~10.311 7%表型变异, 其中来自F2:3群体的qMIC-D11-1 解释的表型变异率最大, 其加性效应 值 为-0.139 8。 4 个QTLs 中,F2群 体 的qMIC-D11-1 与F2:3群体的qMIC-D11-1 的增效基因来自于亲本农垦5 号,其余来自中棉所679。
共检测到4 个与断裂伸长率相关的QTLs,其中1 个来自F2群体,3 个来自F2:3群体。 4 个QTLs 分布在4 个不同的连锁群,解释了7.1335%~8.784 9%的表型变异, 其中来自F2:3群体的qBE-L4-1 解释的表型变异率最大,其加性效应值为-0.148 7。 4 个QTLs 中, 只 有F2群 体 的qBE-A02-1 的增效基因来自亲本中棉所679,其余来自农垦5 号。
3 讨论
3.1 QTL 位点的稳定性
对比前人研究,在李超[20]等陆海杂交群体的纤维品质定位中,2 个与本研究共有的标记NAU5046 与NAU5128 分别被定位到D04 与D08 染 色 体 上;Wang 等[21]利 用 陆 地 棉RIL 群 体在标记NAU3127 和DPL0062 间鉴定到了1 个纤维长度QTL 和1 个马克隆值QTL。 本研究也在这2 个标记间鉴定到了这2 个纤维品质性状相关QTLs,且纤维长度QTL 在F2和F2:3群体中均能检测到,再次验证了NAU3127 和BNL3171间QTLs 的可靠性。其余QTLs 未见前人报道。不同研究中定位到的QTLs 存在难以相互借鉴的现象。 一方面在于目前已经开发的分子标记种类繁多且数量巨大,单一研究中用到的分子标记数量有限;其二是不同亲本、不同环境对QTL 定位影响较大;其三在于不同研究中连锁图谱覆盖基因组的比例和位置不同, 由此鉴定出QTLs 区段也不同。
3.2 QTL 成簇分布
多个不同性状或者不同世代的同一性状定位到同一个标记区间内,说明可能存在基因连锁或者一因多效的现象。 这种现象在不同作物、不同群体、不同环境中均有存在。杨继龙等[22]在棉花早熟和纤维品质的研究中检测到5 个与标记CGR6764a 连锁的QTLs,这些QTLs 与4 个性状相关, 另外在连锁群LG2 的NAU5189 和NAU3377b 之间检测到4 个控制不同性状的QTLs;李帅阳等[23]在纤维品质研究中发现7 个QTLs 聚集在LG17 上32 cM 内,6 个QTLs 聚集在LG52 上23 cM 内。本研究也发现了QTL 成簇分布的现象, 尤其是在F2群体定位到的9 个QTLs, 有7 个 成 簇 存 在, 其 中qFL-A02-1、qBT-A02-1 和qBE-A02-1 聚集在标记BNL3545、NAU3903 之间,qUI-D09-1 和qBT-D09-1 聚集在标记CGR6692、NAU3915 之间,qFL-D11-1 和qBTD11-1 聚集在标记DPL0062、HAU0423 之间。
3.3 多态性标记的数量
本试验从6 688 对SSR 引物中得到了149对多态性标记,多态率为2.23%,低于陆地棉种内一般为4.13%~7.9%的多态性水平[24-25]。 锚定在连锁图谱上的多态性标记也偏少, 为119 个,这可能与筛选多态性标记时采取了严苛的质量控制有关,淘汰了大量清晰度较低、基因型缺失较多的标记。 另外,在筛选到的149 对多态性标记中, 删除了11 个群体基因型相同的标记和8 个偏分离标记。 群体基因型信息完全相同的标记可能受到连锁累赘的影响,导致区段内没有交换单株存在。 连锁图谱构建中经常出现偏分离标记,偏分离影响标记的重组值及排列顺序[26-28],进而影响QTL 定位的结果。 本试验为规避这一影响构图前去除偏分离标记。
3.4 标记在染色体上的分布
在本试验中有3 个连锁群确定了染色体编号,但难以区分是在哪个亚组,还有2 个连锁群未能锚定到染色体。 原因在于这些连锁群中的SSR 标记在参考基因组序列的2 个亚组间均有发现, 甚至有些标记不仅出现在不同亚组间,还出现在了不同染色体上,因此,辨识这些标记的染色体位置便变得非常困难。 这也再次说明了A、D 亚组间的部分同源性, 不同染色体间的DNA 序列也存在一定的相似性。 这种相似性增加了区分标记位置的难度,也给将连锁图谱与物理图谱相结合的研究方法带来了困难。 因此,如何能在保证拥有足够多态性标记的基础上,使用专一性更强的标记,将是解决连锁图谱与物理图谱难以建立联系的关键。
4 结论
本研究利用F2与F2:3群体定位纤维品质形状QTL,从6 688 对SSR 引物中得到了149 对多态性标记,通过对标记的进一步筛选,构建了包含28 个连锁群、119 个标记、 总长为1 173.5 cM的遗传连锁图谱。 根据连锁图谱进行QTL 定位,共得到20 个QTLs, 其中在F2群体中得到9 个QTLs, 在F2:3群体中得到11 个QTLs。 其中,qFL-D11-1 在2 个世代中均能检测到。 发现存在多个QTLs 聚集在同一标记区间的现象, 说明控制不同性状的QTL 可能成簇存在。 利用多世代群体进行QTL 定位有助于发现稳定可靠的QTL位点。 该研究将为以后挖掘纤维品质性状相关基因及分子标记辅助育种奠定基础。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
中国现代医生(2022年21期)2022-08-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年2期)2021-03-29
天津医科大学学报(2021年1期)2021-01-26
现代装饰(2020年7期)2020-07-27
三农资讯半月报(2020年2期)2020-03-09
汽车观察(2019年2期)2019-03-15
