
食品鲜味感知研究进展
2021-02-05 05:53朱忆雯张宁龙
中国食品学报 2021年1期
朱忆雯,张宁龙,姜 水,刘 静,刘 源
(上海交通大学农业与生物学院食品科学与工程系 上海200240)
1 食品感知概述
1.1 食品感知
感知是感觉(Sensation)和知觉(Perception)的整合。感觉是知觉的基础,是人脑对事物个别属性的认识。例如:人能嗅闻到烤肉的香气,看到橘子是黄色的,尝到糖果是甜的,触摸到毯子是柔软的等。当通过感官获得这些外界信息后,大脑对其进行加工与解析,产生对事物的整体认识,进而了解其意义,该过程便是知觉[1]。
食品感知(Food sensory perception)是人类通过视觉、嗅觉、味觉、触觉和听觉等途径感觉、加工、认识和理解食品刺激的动态过程。所感知的信息包括食品的外观(颜色、光泽等)、质地、风味(滋味、气味、口感、三叉神经感)、温度和声音等。通常来说,食品鲜味感知主要是通过口腔加工中释放的鲜味物质与味觉受体结合产生信号,进而大脑整合信息,形成特定的鲜味认知。
1.2 食品感知的重要性
1.2.1 满足生理和心理需求 饥饿和食欲是驱使人类寻找和感知食品的重要生理和心理因素,决定了对食品的选择、接受和摄入等行为。“选择”体现在人类倾向于摄取对人体有益的食物,避免有害物质;“接受”和“摄入”体现在人类将食品的感知结果与个体本身的期望、喜好和需求相衡量,进而决定食品的接受程度、摄入与否及摄入量等[2-3]。比如人类倾向于选择鲜味和甜味的食物,因为鲜味一般意味着食物富含蛋白质或氨基酸[4],而甜味则一般由糖类引起,蕴含生命活动所必需的能量。与之相对,苦味是“负面”滋味,因为食品中的苦味一般意味着食品含有有害物质或发生腐败变质。因此,对于甜味和鲜味的感知能够帮助并引导人类选择更富营养和能量的食品,以维持健康和生命活动;苦味感知能够提醒人类食品中可能存在有害物质,从而避免该类食品的摄入。此外,食品感知还可以诱导唾液、胃酸和胰岛素分泌以及增加食欲等,从而促使身体为摄取食物做好准备[5]。
1.2.2 食品感知的失衡 生理与心理的动态平衡是维持人体健康的基础。个体生理与心理的失衡会导致食品感知的失衡,进而可能诱发暴食或神经性厌食等疾病。神经性厌食症是由于长期限制正常进食或采取暴饮暴食后催吐等方式限制能量摄入,而引起的饮食失控症[6]。行为学研究表明,神经性厌食症患者对食品的关注度较正常受试者更低,且在摄入食品前、后更易产生焦虑情绪和饥饿感,也更易出现过度运动和饮食节制等行为[7-9]。脑科学研究表明,神经性厌食与暴饮暴食可能是大脑对饥饿信号产生错误理解所致。如果大脑无法准确识别饥饿信号,则会导致进食受限,体重减轻,进一步诱发神经性厌食;而如果大脑对饥饿信号产生了“夸大”理解,则容易导致暴饮暴食[10-11]。
1.3 食品感知的影响因素
个体敏感程度、所处状态、既定习惯和文化观念等属于感知主体的影响因素,而食品的状态和性质等属于感知客体的特性。个体对食品的处理方式和进食环境等也是影响食品感知的重要因素。
1.3.1 食品差异 食品本身的差异如外观、成分构成、质构以及包装等是影响食品感知的根本原因。如饼干油脂含量不同会影响其外观和质构,进而影响饼干的观感以及疏松度和适口性。研究表明容器类型(水杯、普通酒杯、鸡尾酒杯和塑料杯)会影响消费者对于不同颜色饮料(红色、绿色、黄色、蓝色、橙色、棕色和无色)的味觉联想,可通过调整容器类型促使跨文化消费者对不同颜色饮料产生相似或不同的风味期望[12];奶酪的颜色过白或过于透明时,都会对消费者的喜好产生负面影响[13]。
1.3.2 个体差异 个体差异是食品感知的另一个重要影响因素。个体差异包括先天因素(遗传因素)和后天因素(所处人文环境、社会人口学特征、个体状态及口腔加工模式等)。遗传因素即个体基因型的不同是最本质的个体差异。TAS1R1(T1R1)基因中单核苷酸多态性(Single nucleotide polymorphisms,SNPs) 如 rs41278020、34160967 和35118458 等,以及TAS1R3(T1R3)基因中SNPs如rs79148073、307377 等与鲜味感知存在潜在相关性[14],其中rs34160967(Ala372Thr)和rs307377(Arg757Cys) 对个体的鲜味敏感性具有重要作用[15-16]。然而,近期研究人员调查了734 名受试者(33 位非洲裔、43 位亚裔、561 位欧裔和97 位其它种族) 的鲜味强度感知和rs34160967 的关系,发现两者之间没有显著性差异[17]。目前对于鲜味敏感度相关基因的研究相对较少,且尚未明确不同鲜味相关SNPs 与不同人种鲜味敏感度的相关性。
人文环境导致的个体差异主要归因于生活中接触到的文化、观念、信仰差异等,进而造成摄入行为、偏好、饮食习惯等的多元化[18]。日常社会人口学特征涵盖了地域、年龄、性别、收入、教育程度、家庭情况等多方面因素。Zhu 等[19]调查发现性别、年龄、婚姻状况、教育程度、工作情况和收入等人口学特征因素显著影响中国消费者对增鲜调味品的认知程度和支付意愿,其中教育程度对自我认知影响更大,月收入对支付意愿影响更大。个体状态则包括情绪状态、生理期状态等。Weenen等[20]发现孕妇在孕早期有更高的苦味阈值,在孕中期偏好甜味,并且在孕中期和孕晚期对咸味食品有更强的食欲。Miranda 等[17]发现受试者(734人) 的饥饿程度和距上次进食后的时长与鲜味强度感知有显著正相关,而生物学因素(年龄、性别、血统和种族)则没有影响。
动态口腔加工是人类感知食品的重要过程,食品口腔加工的概念由浙江工商大学的陈建设教授首次提出[21]。口腔加工过程中大部分因素(唾液分泌量及组成、口腔温度、舌面形貌和咀嚼参数等)都会显著影响食品感知[22-24]。近年来,食品口腔加工摩擦学研究逐渐兴起,主要致力于探究食品结构与性质的变化对口腔摩擦和食品感知的影响。
1.3.3 进食环境 进食环境是影响食品感知的重要因素,包括温度、声音、光线和空气等。近期Bravo-moncayo 等[25]调查了噪音对消费者品尝咖啡体验的影响,结果表明:在噪音较强的环境中,多数消费者对咖啡的感官和享乐属性不太敏感,且购买意愿降低。进食同时捕捉到的视觉信息也影响食品感知,如Liu 等[26]发现相较于纸质材质的桌布,织物桌布会显著影响人对菜品的偏好度,且对餐厅的服务、氛围和整体体验都有较高评价。Hartley 等[27]发现将0.1%脱水菠菜作为视觉提示添加到含有味精的米饭中,可以增强受试者对其鲜味的感知。
2 食品鲜味感知的基础
2.1 物质基础
鲜味物质主要包括游离氨基酸及其盐类、核苷酸及其含类固醇的衍生物、有机酸及其盐类、有机碱类和鲜味肽类等[28-33]。鲜味物质来源广泛[29,32,34],常见的鲜味成分及其食物来源见表1。
2.2 生理学基础
从舌上鲜味受体、味觉信号传导通路到大脑味觉皮层等整个味觉系统构成了鲜味感知的生理基础[64],负责鲜味的识别、味觉信号的产生、转导和加工处理等过程[65]。根据鲜味感知的顺序将哺乳动物的味觉系统分为4 个水平:(1)舌头、味觉上皮和乳突;(2)味蕾和味觉受体细胞;(3)味觉受体;(4)传入神经和大脑味觉皮层[66]。
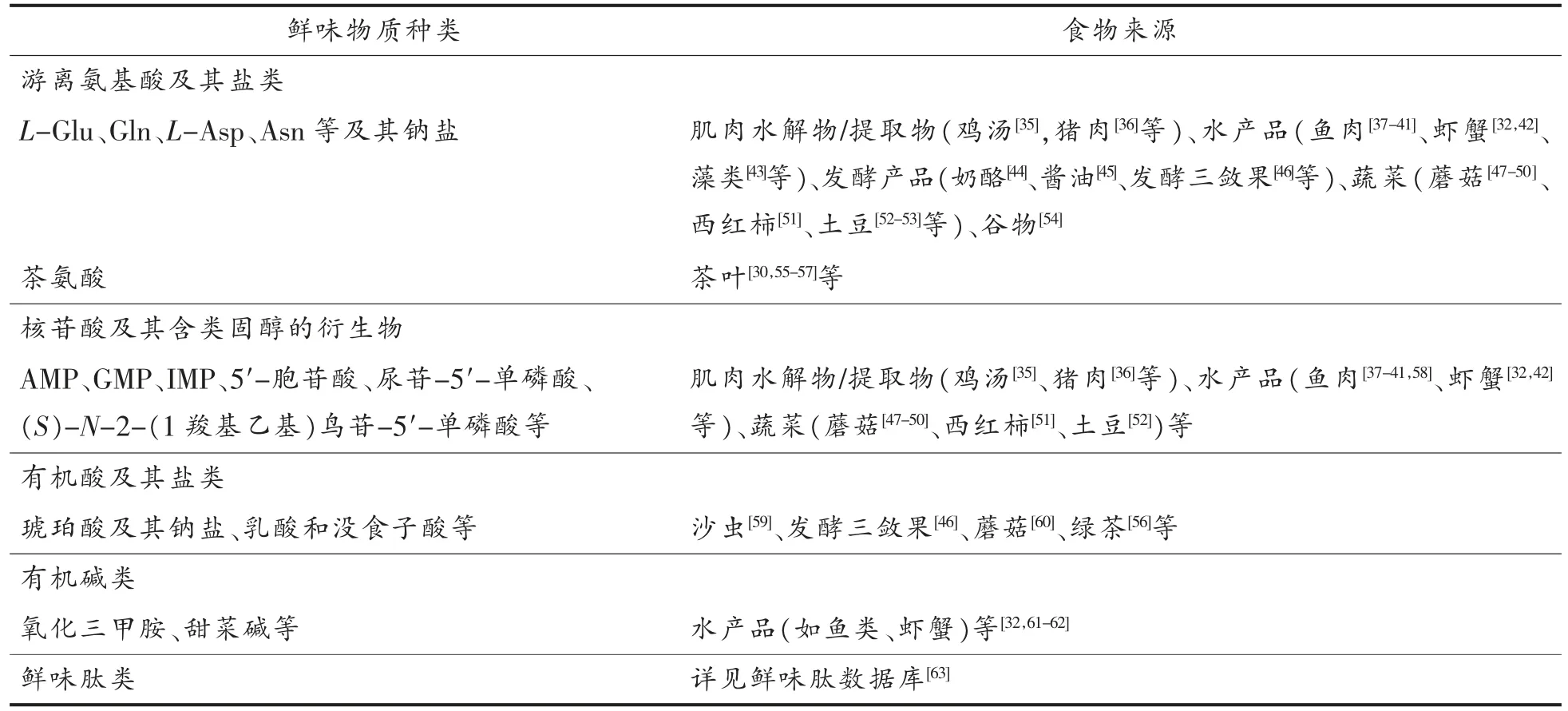
表1 食品中常见的鲜味物质Table 1 Common umami substances in food
2.2.1 舌微观结构 舌头是哺乳动物口腔中主要的味觉感受器官,其次是软腭。舌表面上皮分布着形状各异的味觉乳突[67],其上镶嵌着数量不等的味蕾。味蕾是由多个纺锤形细胞组成的细胞簇,主要存在于味觉乳突上,少部分也嵌入软腭上皮、口咽、会厌和上食道等部位[65,68],是基本的味觉感受单元。组成味蕾的细胞可根据其微观结构和功能特点分为5 种类型[68-69]:I 型(类胶质)细胞、II 型(味觉受体)细胞、III 型(突触前)细胞、IV 型(基底)细胞和V 型(边缘)细胞。其中II 型(味觉受体) 细胞占味蕾细胞的三分之一,膜上表达GPCRs,用于感受鲜味、甜味和苦味[68]。鲜味受体细胞的顶端突出部分带有微绒毛,可延伸抵达味孔,能与口腔中的物质接触并响应鲜味刺激[65]。
2.2.2 鲜味受体 目前报道了8 种鲜味受体,均为GPCRs 跨膜蛋白质[70]。1996年发现了首个鲜味受体(谷氨酸代谢型受体4,metabotropic glutamate receptors 4,taste-mGluR4)[71],2002年Nelson 等[72]发现了T1R1/T1R3 鲜味异二聚体,此后又陆续发现了brain-mGluR4[73],brain-mGluR1[74],tastemGluR1[75],GPRC6A(G-protein-coupled receptor family C,group 6,subtype A class C and group 6 subtype A receptor)[76],胞外钙敏感受体(Extracellular-calcium-sensing receptor,CaSR)[77]和类视紫红质GPCR A 类(Rhodopsin-like GPCR class A,GPR92)[78]等6 个鲜味受体。除GPR92 是A 类GPCRs 外,其它受体均为C 类GPCRs 异二聚体,具有相似结构,由胞外N-端捕蝇草结构域(Venus flytrap,VFT),连接VFT 和跨膜结构域的半胱氨酸富集区(Cysteine-rich domain,CRD),7 次跨膜结构域(7-Transmembrane domain,7-TMD)和胞内C-端结构域组成。
2.2.3 传入神经和大脑味觉皮层 舌头和软腭上的味蕾由鼓索神经(Chorda tympani,CT)、大浅岩神经(Greater superficial petrosal,GSP)和舌咽神经(Glossopharyngeal,GL)3 股外周神经支配[65]。鲜味受体细胞产生的味觉信号刺激传入神经,经外周和中枢神经信息编码、传导后进入大脑味觉皮层[79]。
3 鲜味感知过程
鲜味感知在食品入口前即已发生,随着“第一口”至口腔加工整体过程,鲜味感知涉及鲜味物质释放,鲜味受体-配体结合,味觉信号产生-传递和大脑响应等复杂的感知通路。鲜味信号在大脑中响应后会触发认知和行为反馈,引发生理和情感2 个方面的反应。
3.1 食物入口前的感知
食物的外观、散发的气味和特定食物的声音等,结合人的经验/经历,使人在品尝食物之前产生了期望,从而可能影响口腔中的鲜味体验。研究发现食品的颜色或包装容器会不同程度地影响食物入口前的鲜味强度期望和接受度。例如,酱油的褐色强度与鲜味的预期强度存在正相关[80]。Zhou等[81]研究了面条颜色、容器特征(外观和材质)与面条鲜味认知强度的关系,发现传统面条、绿色和黄色面条的鲜味认知强度更高,红色面条的鲜味预期值较低;用于展示面条的容器不影响参与者判断面条颜色与鲜味预期强度的关系;然而,容器特征与面条的颜色会产生交互作用,从而影响人们对不同颜色面条的主观评价和滋味期望。外形/颜色对鲜味感知的影响更倾向于被人的经验、记忆中某食品的固有属性左右,即认为之前接触过的、常规的食品鲜味更强;当外形/颜色等属性改变时,鲜味感知往往会减弱。
3.2 口腔加工中的感知
食物入口后开始了更复杂的鲜味动态感知过程,涉及鲜味物质释放,鼻后气味对鲜味感知的影响等方面,上述过程在口腔加工过程中反复发生或/和同时进行,产生了每种食品特有的口腔鲜味感知体验。
口腔加工过程可分为口腔准备、口腔呈递、吞咽和食管4 个阶段[82]。鲜味物质在口腔加工阶段持续释放,如红烧肉咀嚼过程中鲜味释放呈先上升后下降的趋势,且在咀嚼中后阶段鲜味感知最强[83]。鲜味物质释放的同时伴随鼻后气味物质的释放和感知,且两者发生相互作用。鼻后气味对鲜味感知有正、负两方面的影响。Bonito 鱼干的鼻后气味和馏出香味物质在含NaCl 的溶液中可诱发并增强鲜味强度[84-85];相反,基于小鼠行为学实验发现己醛能够减弱厌食小鼠对MSG 的鲜味感知[86]。此外,研究发现奶酪中的10 种关键香味组分[87],日本大豆酱中的二甲基三硫醚[87]和桂林白腐乳中的(E,E)-2,4-癸二烯醛、1-辛烯-3-醇、(E,E)-2,4-壬二烯醛、丁香酚、己酸乙酯和苯乙醛等关键香味组分[89]均可增强鲜味和鲜味后味。
从微观角度分析,口腔加工过程配体(鲜味物质)不断刺激鲜味受体,两者发生动态相互作用。鲜味受体T1R1/T1R3 为异源二聚体,T1R1-VFT为T1R1 的正构结合位点,L-Glu 密切结合在T1R1-VFT 铰链区域;5’-核苷酸,如GMP 和IMP则结合在T1R1-VFT 的开口区域,进一步稳定了T1R1 的闭合构型(活性状态,图1右上所示),由此可从分子层面解释Glu 和5’-核苷酸的鲜味协同增效机制[90-91]。另外,T1R1 的7-TMD 是鲜味化合物N-(庚-4-基)苯并[d][1,3]二氧基-5-羧基【N-(heptan-4-yl)benzo [d][1,3]dioxole-5-carboxamide】的结合位点[90]。T1R3 为T1R1/T1R3 与甜味受体T1R2/T1R3 的共有亚基,而T1R3 的7-TMD为2 种甜味相关物质2-(4-甲氧基苯氧基) 丙酸(Lactisole,甜味抑制剂)和甜蜜素的结合区域,故鲜味受体T1R1/T1R3 也能响应上述2 种物质的刺激,前者抑制T1R1/T1R3 的活性,而甜蜜素增强了鲜味受体对L-Glu 的敏感性[92]。
鲜味肽的感知机制也备受关注,研究发现T1R1-VFT 是鲜味肽的潜在结合位点[93-94]。然而,同时有研究揭示鲜味肽主要结合在T1R3-VFT(图1右下所示),当MSG 与鲜味肽共存时有助于T1R3-VFT 铰链区扩大,促使鲜味肽等大分子与T1R3-VFT 结合[70,95-96]。目前的研究尚未得出一致的结论,有待进一步研究阐明。
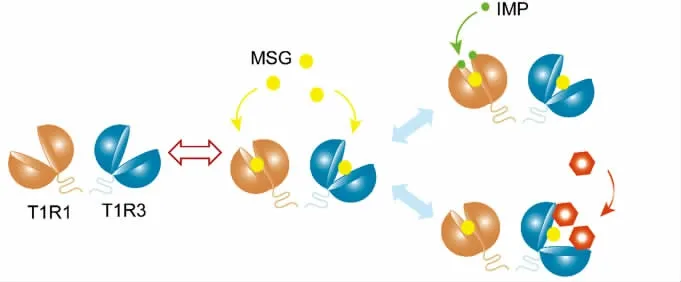
图1 MSG、IMP/GMP 和鲜味肽与T1R1/T1R3-VFT 结合时的构象变化模型图Fig.1 Model diagram of conformational changes of MSG,IMP/GMP and umami taste peptides combined with T1R1/T1R3-VFT
3.3 鲜味感知通路
3.3.1 鲜味信号的胞内通路 T1R1/T1R3 与鲜味配体结合被激活后,G 蛋白亚基解离。其中解离后的Gβ/γ与磷脂酶C-β2(Phospholipase Cβ2,PLCβ2) 相互作用,进而将磷脂酰肌醇4,5-二磷酸(Phosphatidylinositol 4,5-bisphosphate,PIP2)裂解为肌醇-1,4,5-三磷酸(Inositol 1,4,5-triphosphate,IP3) 和二酰基甘油(Diacylglycerol,DAG)。IP3 激活III 型IP3 受体(Type III IP3 receptor,IP3-R),刺激Ca2+从内质网(Endoplasmic reticulum,ER)释放到细胞质中。胞内Ca2+升高激活瞬时受体电位阳离子通道亚家族M 成员5 通道(Transient receptor potential cation channel subfamily M member 5,TRPM5),导致膜去极化,促使ATP通道Pannexin-1(Px-1)和钙稳态调节剂1(Calcium homeostasis modulator 1,CALHM1)[97]打 开并释放神经递质ATP,进而刺激传入神经纤维,实现味觉受体细胞内的信号转导(图2)[68,98]。
3.3.2 鲜味信号的传导通路 受体细胞产生的信号刺激传入神经,经外周和中枢神经信息编码、传导后进入大脑。具体而言,CT 介导鲜(强烈)、咸和甜味,GL 主要传导苦味和酸味,GPS 对甜(强烈)、鲜味敏感[99-100]。其中CT 和GSP 汇入膝状神经节,GL 汇入岩神经节,一起传至孤束核(Neural stations of the solitary tract,NST)[68]。味觉信号从受体细胞到NST 之间的传导主要由神经系统参与,微观神经角度的调控异常复杂而精细化[68]。传入神经节-神经元(Afferent ganglion neurons)从味觉细胞接收单个味觉信号的输入,它们称为专一神经元。其它传入神经节-神经元接收来自多种味觉细胞或III 型细胞信号的输入,因此是多敏感的“多面手”神经元。汇至NST 的神经信号途经丘脑腹后内侧核的细胞旁部位(Parvicellular portion of the ventroposteromedial nucleus of the thalamus,VPMpc),并终止于脑岛皮层(Insular cortex,IC)和眶额皮质(Orbitofrontal cortex,OFC),完成味觉信号的传递[28,101]。
3.4 大脑响应及反馈
3.4.1 鲜味在大脑中的活性区域 味觉信号输入到大脑皮层后,引起特定空间区域的显著响应,即大脑味觉地图(Gustotopic map)[102-104]。基于功能性磁共振成像(Functional magnetic resonance imaging,fMRI)等技术[104-105],研究表明鲜味激活的人类大脑活性响应区域主要包括初始味觉皮层(Insular/opercular cortex)和次级味觉皮层(OFC)[106-110],其次为前扣带回皮层(Rostral anterior cingulate cortex,ACC)[106,111]、海马旁回(Parahippocampal gyrus)[111]、丘脑[109]和前脑盖皮层(Frontal operculum)[109]等区域。
鲜味感知的个体差异也体现在大脑对鲜味信号响应的分布区域上。Han 等[109]采用fMRI 技术调查了对鲜味具有不同识别能力的个体的大脑鲜味响应特征,以及反复接受鲜味刺激对鲜味识别和味觉神经活动的影响。研究发现,低鲜味识别者在丘脑、海马和后脑岛中具有更强的鲜味激活响应,而高鲜味识别者在味觉初级皮层(前脑盖皮层和中央后回)和次级味觉皮层OFC 中具有更强的激活响应。同时,低鲜味识别者经过长达3 周的鲜味反复刺激后,鲜味识别能力显著提升,然而并未反映在神经激活的变化中。即注意力和联想/记忆相关的大脑结构在鲜味感知中起重要作用,而神经激活的变化较小。另外,fMRI 研究发现健康女性和神经性厌食症患者对由MSG 激活的大脑响应也明显不同[110]。与健康女性相比,厌食症患者的右脑岛对MSG 刺激的激活程度更高,行为上表现为对MSG 喜好度的降低。
3.4.2 鲜味刺激后大脑反馈 消费者摄入食物后,主要引发包括生理和情感两方面的反馈,前者如大脑活动、自主神经系统活动和面部表情等,后者如幸福感、喜欢/厌恶和中性情感反馈等[112]。研究发现餐前摄入含MSG 的肉汤会降低参与者的食欲,减少食物摄入量,尤其是对有暴食症状和超重女性而言[113-116]。近期Magerowski 等[117]结合行为学和fMRI 技术探究了上述现象的神经认知机制:在视觉模拟自助餐的行为测试中,摄入含MSG 的肉汤的参与者对于饱和脂肪的摄入量减少且对不同餐盘的注视切换率降低;fMRI 结果显示,与控制饮食决策相关的左外侧前额叶皮层的参与度增加。
4 鲜味评价
鲜味评价方法包括理化指标评价、感官评价、智能感官技术、心理物理技术和其它技术等,目前尚无统一的标准评价方法。
4.1 理化指标
MSG、鲜味氨基酸和鲜味核苷酸等含量可部分表征食品的鲜味强度。滋味活性值(Taste activity value,TAV) 为滋味化合物的含量和其阈值的比值,可用于判定单个化合物在某一体系中的滋味贡献。当TAV>1 时,表明该化合物对样品的滋味可能有重要贡献。例如,Glu 在炭烤羊腿中的TAV 最高可达6.54,与鲜味感官强度呈显著正相关[118]。同样的,研究表明Glu 对甜面酱的鲜味也有重要贡献[119]。IMP 对紫菜的滋味贡献较大[120]。
鲜味当量值(Equivalent umami concentration,EUC)由日本学者Yamaguchi[121]提出,将鲜味氨基酸和鲜味核苷酸混合溶液所呈现的鲜味强度转化为等价的谷氨酸钠(即味精)的浓度,用于量化混合溶液的鲜味强度。EUC 在水产品和食用菌等食品中得到较为广泛的应用,如中华绒螯蟹(Eriocheir sinensis)[122]、长江刀鲚(Coilia ectenes)[42]、北欧鱿鱼(Loligo forbesii)[123]、鹦鹉螺(Pleurotus geesteranus)[124]、暗纹东方鲀(Takifugu obscurus)、红鳍东方鲀(Takifugu rubripes)[125]和食用菌[126-127]等。
在食品体系中,鲜味感知归因于鲜味成分的种类与含量、食品质构等多因素的交互作用。TAV仅考虑单个成分的滋味作用,EUC 仅考虑鲜味氨基酸和鲜味核苷酸之间的协同作用[128],尚需与感官评定结合进一步做鲜味评价。
4.2 感官评价
感官评价可获得评价员或消费者对产品最直观、真实的感受。食品鲜味的感官评价涉及阈上强度感知范围(Supra-threshold intensity perception range)。个体对MSG 的觉察或识别阈值可反映个体对鲜味的敏感程度,检测方法包括单一刺激法、二点选配法(Two-alternative forced choice,2-AFC)和三点选配法等。其中,2-AFC 为目前常用的阈值测定方法[15,129]。
目前,鲜味强度定量感官评价尚未有统一的方法,通常采用标度法(五点/七点/九点/十点/十一点或线性),参比样和浓度的设置差别较大。如顾艳君等[130]采用鲜味参比样:以质量浓度1 g/100 mL 的MSG 溶液(另含0.3 g NaCl)的鲜味强度标示为1 度,谷氨酸钠为100 度。Zhang 等[53]以0.225 g/100 mL 的MSG 溶液(另含L-MSpG 和D-MSpG)作为十点标度上的5 分;Teo 等[131]以0.12,0.3,0.7 g/100 mL 的MSG 溶液分别作为100点标度上的28,43,69 分进行感官培训。
2-AFC 测试也可用于确定食品的鲜味强度。Zhu 等[132]根据Steven 心理物理函数构建了MSG浓度-鲜味感官强度方程:R=72.70S0.82(式中,R——鲜味强度值;S——MSG 的质量浓度,g/100 mL)。采用2-AFC 法,根据50%选择率原则可确定样品的相对MSG 质量浓度,结合上述方程定量其鲜味感官强度值。该方法在评价草鱼和暗纹东方鲀鱼肉及鱼汤的鲜味强度中得到应用。
4.3 智能感官分析
智能味觉感官模拟人类味觉系统,通常包括传感器阵列和模式识别系统,可以客观评价食品滋味。目前已有数款用于食品鲜味分析的商用电子舌系统,国内外也开发了部分鲜味仿生传感器。
4.3.1 商用电子舌 味觉传感器根据工作原理可分为电化学传感器、光学传感器、质量传感器和仿生传感器等[133-134]。其中,电化学传感器因具有较好的稳定性和广谱性,故常用于构建商用电子舌。常见的商用电子舌包括中国、日本、法国公司开发的产品,其类型和在鲜味评价方面的应用如表2所示。目前,电子舌用于鲜味定量评价研究相对较少,需要将电子舌与人工感官评价结果的相关性作为重要评判标准。一些研究报道在豆酱(r=0.821)[135]、绿茶(r=0.86)[136]和鲜味肽[137]等样品的鲜味强度评价中,电子舌输出数据和感官评价结果具有较好的相关性。

表2 商用电子舌的类型及其应用领域Table 2 Overview of the types and configurations of common commercial electronic tongues
4.3.2 仿生味觉传感器 仿生味觉传感器以酶、组织、细胞以及受体等作为识别鲜味物质的敏感元件,结合二级传感器检测敏感元件和配体之间的特异性反应。由于酶具有良好的特异性和催化能力,因此基于酶催化氧化还原的生物传感器被广泛应用于鲜味成分定量检测。如基于L-谷氨酸氧化酶和L-谷氨酸脱氢酶的生物传感器用于评估番茄[150-151]、大米汤[152]及调味料[153]中谷氨酸的含量,以及基于5’-核苷酸酶和黄嘌呤氧化酶的双酶电化学传感器用于肉制品中肌苷酸含量的检测[154-155]。
以组织、细胞和受体为敏感元件的仿生味觉传感器中,Huang 等[156]开发了基于T1R1 的电化学生物传感器,评价4 种鲜味配体(GMP、MSG、IMP和WSA)与鲜味受体之间的相互作用。Chen 等[157]通过构建受体传感器研究了3 种鲜味受体(T1R1/T1R3,mGluR4,mGluR1)与鲜味物质IMP 和MSG的感知差异。由于鲜味受体结构的复杂性,为简化传感器构建过程,开发了以T1R1 鲜味配体结合域VFT 为敏感元件的场效应管传感器[158]。此外,从哺乳动物中分离得到带有受体的纳米囊泡具有更好活性,也已成功用于鲜味检测[159-161]。
4.4 心理、生理指标
感官评价的结果主要基于人的主观报告,而心理、生理指标则是对人体反应的客观表征。食品鲜味感知过程涉及大脑、肌肉等多机体功能,可采用行为学、解剖学、生理学、神经科学等方法分析人对鲜味刺激的反应。
在行为学、解剖学和生理学方面,测量面部反应在参与者无法用语言表达或在观察性研究中具有实用性。面部反应可以通过面部动作编码系统(Facial action coding system,FACS) 进行测量和分析[162]。有研究发现虽然被试者对鲜味刺激没有表现出强烈的面部反应,但是通过综合分析5 种基本味觉后发现面部反应能够同时区分刺激物的性质和浓度[163]。Zhi 等[164-166]尝试运用自动面部表情分析技术表征亚洲消费者对不同浓度滋味溶液的情感特征,准确度最高可达到74.3%。肌电图(Electromyography,EMG) 技术可测量面部肌肉的电位和面部血流指标等,眼动追踪可识别被试者眼部动作,能够提供关于消费者的期望、感知、记忆、情绪和决策过程的信息,并用这些信息解释和预测其行为,可被进一步运用以探索鲜味刺激引起的面部反应中。
4.5 其它测定方法
4.5.1 动物味觉实验 啮齿类动物等哺乳动物具有与人类相似的鲜味受体,可用于探索已知滋味物质的剂量-反应关系。动物味觉实验包括简短摄取法(Brief access test)和双瓶偏好法(Two-bottle choice paradigm)等,可通过动物对实验样品的摄取次数判断其偏好程度[167],或是摇头、躁动等一些逃避反应进行综合评价[168]。Narukawa 等[169]通过小鼠行为学研究(48 h 双瓶偏好测试和条件性味觉厌恶测试)和味觉神经反应,评价了琥珀酸钠、L-茶氨酸、甜菜碱和D-MSG 等物质是否呈鲜及各物质与IMP 的协同作用。对小鼠而言,琥珀酸钠具有鲜味并且与IMP 显示出强协同作用;L-茶氨酸和D-MSG 不具有鲜味,而L-茶氨酸在IMP 存在时呈鲜,D-MSG 与IMP 的协同较弱。
4.5.2 计算化学 通过计算化学模拟与分析鲜味物质的性质、对接和计算鲜味配体与受体的结合位点和动力学参数,可为鲜味分子评价提供辅助手段。在定性方面,可通过结合区域和活性位点等判断物质是否呈鲜,如BMP 与鲜味受体的结合区域主要位于其肽序列中的负电基团[93];且当鲜味肽的活性位点位于D 或E 基团时,具有更好的呈鲜特性[170]。在定量方面,可预测鲜味物质的鲜味强度。余霞琴[171]分析了鲜味六肽与鲜味受体之间形成的复合物结构,结合能大小能够准确反映配体与受体间的结合强度,进而预测其鲜味强度。Yang等[172]通过同源模型、分子对接和MD 模拟等研究,发现浓厚感(Kokumi) 味的γ-谷氨酰胺肽与T1R3-MSG 具有相互作用,证明γ-谷氨酰胺肽可以增强MSG 的鲜味强度。
5 小结与展望
食品感知是未来食品个性化需求和智能制造的重要基础。食品鲜味感知是一个系统而复杂的过程,其影响因素繁多,涉及心理学、化学、生理学、神经科学等学科。食品鲜味感知对解释人类摄食和偏好行为等方面具有重要意义。目前,对食品鲜味感知的物质与生理基础、感知机制和评价技术等均有一些研究,然而仍存在较多未知或不明确之处。鲜味感官强度的评价方法尚需进一步标准化。
面向未来,食品鲜味感知科学可能着重于以下3 个方面:(1)食品鲜味感知的基础研究,包括新型鲜味物质的发掘、鲜味感知机制、个体差异和脑科学等相关研究;(2)食品鲜味感知精准分析技术;(3)感官交互与消费者研究。
猜你喜欢
美食(2022年2期)2022-04-19
美食(2022年4期)2022-04-16
食品安全导刊(2021年20期)2021-08-30
疯狂英语·新读写(2021年2期)2021-02-25
中国调味品(2019年2期)2019-03-18
旅游世界(2018年7期)2018-08-07
中国化妆品(2017年12期)2017-06-27
哈尔滨医药(2016年3期)2016-12-01
癌变·畸变·突变(2016年3期)2016-02-27
