
蕨类植物染色体数目研究的好材料
2021-03-24 18:30王紫娟向建英成晓
广西植物 2021年1期
王紫娟 向建英 成晓
摘 要: 染色体数目分析是现代系统与进化植物学研究十分重要和不可缺少的环节。在蕨类植物中,使用根尖或孢子母细胞作为实验材料的经典方法在取样时往往存在困难。该文以3种桫椤科桫椤属植物为例,探寻适合蕨类植物染色体数目研究的材料。由于取样时没有时间、季节和数量上的限制,我们推荐配子体作为蕨类植物染色体数目研究的实验材料。
关键词: 桫欏属, 染色体计数, 桫椤科, 配子体, 实验材料
Abstract: Analysis of chromosome numbers is a very important and indispensable step in modern systematic and evolutionary botany research. In ferns, the classical methods using root tips or sporocytes as materials often have difficulties when sampling. In this paper, three species of Alsophila (Cyatheaceae) were taken as examples to explore the materials suitable for the study of chromosome numbers of ferns. As there are no time, season and quantitative restrictions in sampling, gametophytes were recommended as experimental materials for the study of chromosome numbers of ferns.
Key words: Alsophila, chromosome count, Cyatheaceae, gametophyte, experimental material
染色体数目分析是现代系统与进化植物学研究十分重要和不可缺少的环节。对于那些对分类、起源和进化感兴趣的植物学家来说,染色体数目分析是研究任何一类植物的一个基本环节(Guerra,2008)。植物体内的染色体数目能提供物种种内和种间遗传分化的信息,这有助于我们在各个分类等级上理解植物的系统发育关系(Windham & Yatskievych,2003;Glick & Mayrose,2014)。认识到染色体数目数据对植物系统进化研究的重要性,研究人员已经做了很多努力来尽可能多地确定物种和种群的染色体数目。
有花植物的染色体研究可以追溯到19世纪,而蕨类植物的染色体研究直到20世纪才开始,并且在最初的20年内只有不到20种蕨类的染色体数目被报道过,这一方面由于蕨类植物的染色体数目普遍高,并且染色体小,用传统的石蜡切片法很难进行准确的染色体计数(吴兆洪,1984;王任翔等,2007);另一方面由于蕨类植物的根尖细胞含有坚硬的黏性物质,采用有花植物根尖水解的方法效果不理想,很难制作压制均匀、染色体清晰的装片。因此,直到1950年Manton的书Problems of Cytology and Evolution in the Pteridophyta(《蕨类植物的细胞学和进化问题》)出版后,蕨类植物的染色体准确计数才得以实现。Manton采用孢子母细胞醋酸洋红压片技术来解决蕨类植物的染色体计数问题。蕨类植物的染色体研究虽然起步较晚,但发展速度却比显花植物快得多。在Manton的书出版之后的20年内,有2 000多种蕨类植物的染色体数目被报道出来,占世界总蕨类植物的20%(Walker,1973)。在20世纪80年代到90年代初,蕨类植物的染色体研究达到了高峰(王任翔等,2007),此后,相关的研究论文急剧减少。一方面,可能因为分子生物技术的快速发展,转移了科研人员的研究兴趣;另一方面,Manton采用的孢子母细胞压片法也有一定的局限性,比如难以获取合适的孢子母细胞。邵文(2013)建议使用胚胎或幼孢子体进行蕨类植物的染色体研究,但从孢子到胚胎和幼孢子体的形成需要较长时间,会影响研究进展。因此,有必要为蕨类植物染色体数目研究寻找一种更方便、更有效的实验材料。
本研究以孢子来源的配子体为材料,以3种桫椤科(Cyatheaceae)桫椤属(Alsophila)蕨类为例,探讨配子体在蕨类植物染色体计数研究中的优势。
1 材料与方法
用于染色体计数的实验材料是用孢子培养出来的配子体。孢子可以从保存好的标本上收集或从活植物体上采集。
用于本研究的蕨类为黑桫椤(A. podophylla)、大叶黑桫椤(A. gigantea)以及滇南桫椤(A. austro-yunnanensis)。2015年,在云南南部的西双版纳热带植物园、小腊公路以及河口至屏边的公路旁分别收集了它们的孢子(表1),在实验室里将其播种于腐殖土中,进行配子体培养,当发育成幼原叶体或心型原叶体具有良好的分生组织后(1~3周),用0.002 mol·L-1的8-羟基喹啉溶液预处理3~5 h。首先用Farmers固定液(95%乙醇∶冰醋酸=3∶1)在4 ℃固定0.5 h以上,依次用95%、70%和50%酒精以及蒸馏水洗脱,然后用1 N盐酸在60 ℃的水浴锅中解离6~10 min,最后在1%醋酸洋红中对配子体材料进行压片。
配子体有丝分裂细胞经过预处理、固定、解离、染色和压片后,用奥林巴斯BX51显微镜观察拍照。
2 结果与分析
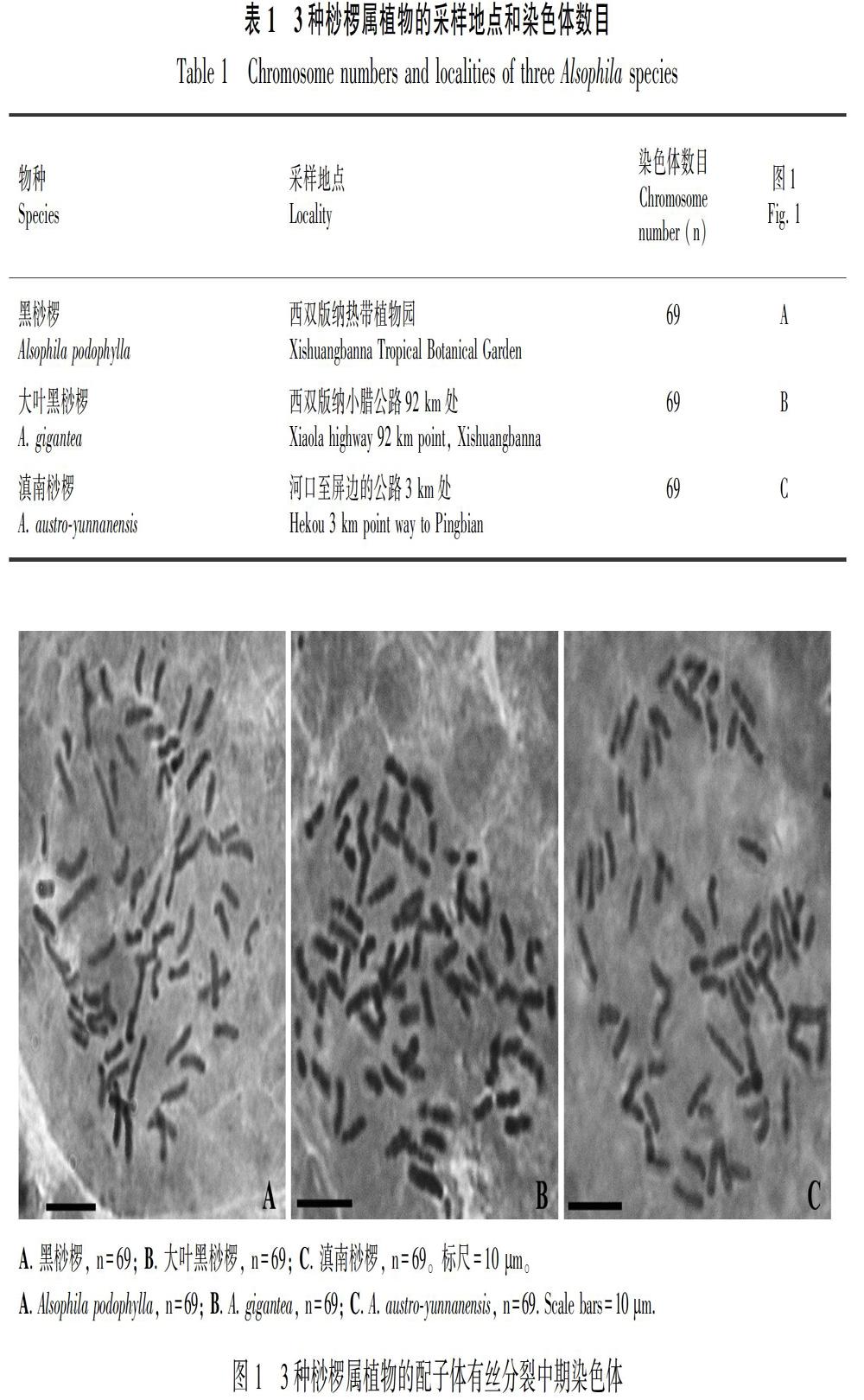
染色体观察结果汇总于表1。这三种蕨类的染色体都很分散,易于观察和计数,n=69(图1)。
黑桫椤在中国的香港、台湾、贵州、云南、福建、广东、广西以及海南都有分布,另外也分布于日本、越南、老挝、泰国以及柬埔寨等国家(张宪春,2004)。早在1965年,Roy & Holttum就对广东鼎湖山的黑桫椤材料进行了染色体数目报道;在1989年,Nakato对分布于日本的黑桫椤进行了染色体研究,均以根尖为材料,发现黑桫椤为二倍体,报道其染色体数目分别为2n=c.140和2n=138。本研究中,以配子体为实验材料,观察到的结果与他们报道的一致,黑桫椤配子体的染色体数目为n=69(图1: A),黑桫椤为二倍体物种。
大叶黑桫椤分布于中国的云南、广东、广西以及海南,在日本的南部、马来半岛、老挝等东南亚国家以及印度等南亚国家也有分布 (张宪春,2004)。早在1954年,Manton & Sledge采用孢子母细胞醋酸洋红压片法对分布于南亚锡兰(现更名为斯里兰卡)的大叶黑桫椤进行了染色体数目研究,发现n=69;在1962年,Abraham et al. 以根尖为材料,对分布于印度南部的大叶黑桫椤进行了染色体分析,得到2n=138;在1999年,Kato & Nakato同样以根尖为实验材料,对采自中国海南的材料进行了染色体报道,2n=138。本研究以配子体为材料,观察到n=69(图1: B),与他们报道的结果一致,大叶黑桫椤是一个有性二倍体物种。
滇南桫椤为中国特有种,主要分布在云南的屏边和麻栗坡(张宪春,2004)。本研究材料采自屏边,此次为首次对其染色体数目进行报道。我们观察到滇南桫椤配子体的染色体数目为n=69(图1: C),它也是一个有性二倍体种。
3 讨论与结论
以孢子来源的配子体为材料,观察到黑桫椤和大叶黑桫椤的染色体数目与以往报道的采用根尖或孢子母细胞为材料的染色体结果一致,但利用配子体为实验材料的优势却是显而易见的。
对蕨类植物进行染色体研究时,通常采用根尖或孢子母细胞为材料,但根尖或孢子母细胞的生长活跃期非常有限,这就限制了取样时间,取样的困难阻碍了很多植物的细胞学研究。以孢子培养来源的配子体为材料,取样时间不限,只要给予适当的人工条件,孢子一年四季均可培养,配子体取样不受具体时间、季节影响。孢子可以从标本馆标本或活植物体上收集。如果储存得当,蕨类植物的孢子可以存活数十年(Windham & Haufler,1986)。此外,孢子培养配子体的技术非常简单,可为研究人员提供足够多的样本材料进行细胞学研究,特别是对于那些难以判断根尖生长活跃状态或难以把握孢子母细胞产生精确时间的没有经验的人来说。通过孢子培养产生的配子体在一年中任何时候都可获得,打破了季节和时间的限制,减少了野外采样的工作量。这种方法在处理难于获得的蕨类植物时特别有利,例如桫椤科(Cyatheaceae)和碎米蕨类(Cheilanthoid ferns)植物。桫椤科植物是一类特殊的蕨类植物,具有树状根茎,1975年被列入“濒危物种国际贸易公约”(CITES),是中国二级重点保护植物。野外直接挖取根尖会使这些物种在野外的生存受到极大的威胁。桫椤科是一个泛热带大科,全球分布着约500种现存物种(Tryon & Gastony,1975);然而,到目前为止,被计算染色体的物种还不到40种。碎米蕨类通常分布于偏僻的地方很难成功移栽,孢子囊群小,并由毛、鳞片和反折的叶边覆盖,很难获得适宜的根尖材料或孢子母细胞;在Windham & Yatskievych(2003)以孢子萌发的配子体为实验材料进行细胞学研究之前,只有不到20%的碎米蕨类染色体被报道出来。况且,蕨类植物根尖含有难以水解的黏性物质,同时蕨类植物的染色体小、数目多,很难获得染色体完全分散的装片,而蕨类植物的配子体只有一层细胞构成,易于压制,染色体数量是根尖细胞的一半,因而更容易被观察和统计。
Manton(1950)在她的书Problems of Cytology and Evolution in the Pteridophyta(《蕨类植物的细胞学和进化问题》)中提到了这种方法,但除了Roy & Manton(1965)、Gastony & Haufler(1976)、Windham & Haufler(1986)、Windham & Yatskievych(2003)、Dong(2011)外,很少有研究者利用配子体进行蕨类植物的细胞学研究。大多研究仍然使用根尖或孢子母细胞作为蕨类细胞学研究的实验材料(卢金梅等,2006;Hsieh et al.,2008;Wang et al.,2011;王任翔等,2013)。
基于本文介绍的3种蕨类植物所取得的积极结果,以及所提到的众多优点,我们强烈推荐将配子体作为蕨類植物的细胞学研究材料。采用孢子萌发的配子体为实验材料,可以弥补根尖或孢子母细胞取样的不足。
参考文献:
ABRAHAM A, NINAN CA, MATHEW PM, 1962. Studies on the cytology and phylogeny of the pteridophytes. Ⅶ. Observations on one hundred species of South Indian ferns [J]. J Ind Bot Soc, 41(3): 339-421.
DONG SY, 2011.Taxonomic studies on Asplenium sect. Thamnopteris (Aspleniaceae) I: Cytological observations [J]. Am Fern J, 101(3): 156-171.
GASTONY G, HAUFLER CH, 1976. Chromosome numbers and apomixes in the fern genus Bommeria (Gymnogrammaceae) [J]. Biotropica, 8(1): 1-11.
GLICK L, MAYROSE I, 2014. ChromEvol: Assessing the pattern of chromosome number evolution and the inference of polyploidy along a phylogeny [J]. Mol Biol Evol, 31(7): 1914-1922.
GUERRA M, 2008. Chromosome numbers in plant cytotaxonomy: Concepts and implications [J]. Cytogenet Genom Res, 120(3-4): 339-350.
HSIEH TH, CHANG YH, CHIOU WL, et al., 2008. Chromosome numbers of five species of the Marattiaceae in Taiwan [J]. Taiwan J For Sci, 23(4): 377-381.
KATO M, NAKATO N, 1999. A cytotaxonomic study of Hainan(S. China) pteridophytes, with notes on polyploidy and apogamy of Chinese species [M]// ZHANG XC, SHING KH. Ching Memorial Volume. Beijing: China Forestry Publishing House: 1-19.
LU JM, LI DZ, WU D, 2006. Chromosome numbers of four genera in the Dryopteridaceae [J]. Acta Phytotax Sin, 44(5): 516-522. [卢金梅, 李德铢, 吴丁, 2006. 鳞毛蕨科四个属的染色体数目(英文) [J]. 植物分类学报, 44(5): 516-522.]
MANTON I, 1950. Problems of cytology and evolution in the pteridopyhta [M]. Cambridge: Cambridge University Press.
MANTON I, SLEDGE WA, 1954. Observations on the cytology and taxonomy of the pteridophyte flora of Ceylon [J]. Philos Trans Roy Soc London B, 238(654): 127-185.
NAKATO N, 1989. Cytological studies on the genus Cyathea in Japan [J]. J Jpn Bot, 64(5): 142-147.
ROY SK, HOLTTUM RE, 1965. Cytological observations on ferns from Southern China [J]. Am Fern J, 55(4): 154-158.
ROY SK, MANTON I, 1965. A new base number in the genus Lygodium [J]. New Phytol, 64(2):286-292.
SHAO W, 2013. Cytological methods in fern by using embryo and young sporophyte [J]. Acta Bot Boreal-Occident Sin, 33(2):407-410. [邵文, 2013. 利用胚胎和幼孢子体进行蕨类细胞学研究 [J]. 西北植物学报, 33(2):407-410.]
TRYON RM, GASTONY GJ, 1975. The biogeography of endemism in the Cyatheaceae [J]. Fern Gazette, 11(2/3): 73-79.
WALKER TG, 1973. Evidence from cytology in the classification of ferns [J]. Bot J Linn Soc, 67(Suppl. 1): 91-110.
WANG RX, LU SG, DENG XC, 2007. Cytotaxonomic studies of the Chinese pteridophytes: A review [J]. Acta Phytotax Sin, 45(1): 98-111. [王任翔, 陆树刚, 邓晰朝, 2007. 中国蕨类植物细胞分类学研究概况 [J]. 植物分类学报, 45(1): 98-111.]
WANG RX, SHAO W, LU SG, et al., 2011. Cytotaxonomic study of 12 species in the Polypodiaceae from Southern China [J]. Am Fern J, 101(4): 307-316.
WANG RX, LIU L, SHAO W, et al., 2013. Cytotaxonomic study on two species of the Arthropteris from Guangxi, China [J]. Guihaia, 33(6): 723-726. [王任翔, 劉灵, 邵文, 等, 2013. 广西爬树蕨属两种植物的细胞分类学研究 [J]. 广西植物, 33(6): 723-726.]
WINDHAM MD, HAUFLER CH, 1986. Biosystematic uses of fern gametophytes derived from herbarium specimens [J]. Am Fern J, 76(3):114-128.
WINDHAM MD, YATSKIEVYCH G, 2003. Chromosome studies of Cheilanthoid ferns(Pteridaceae: Cheilanthoideae) from the western United States and Mexico [J]. Am J Bot, 90(12): 1788-1800.
WU SH, 1984. A survey of cytology in the pteridophyta [J]. Guihaia, 4(1): 1-8. [吴兆洪, 1984. 蕨类植物细胞学概说 [J]. 广西植物, 4(1): 1-8.]
ZHANG XC, 2004. Cyatheaceae [M]//Flora Reipublicae Popularis Sinicae. Beijing: Science Press, 6(3): 249-274. [ 张宪春, 2004. 桫椤科 [M]//中国植物志. 北京: 科学出版社, 6(3): 249-274.]
(责任编辑 李 莉)
