
A new species of the genus Micryletta (Anura,Microhylidae) from Hainan Island, China
2021-04-15 04:36Jian-HuanYang,NikolayA.Poyarkov
Zoological Research 2021年2期
关键词:海南
DEAR EDITOR,
We report on a new amphibian species of the genusMicrylettafrom Hainan Island, China, based on morphological and molecular analyses.The new species,Micryletta immaculatasp.nov., is diagnosed by a combination of the following morphological characters: medium-sized within genus (SVL 23.3-24.8 mm in males,n=3; 27.7-30.1 mm in females,n=2);dorsum bronze brown to reddish brown in life; dark brown spots and stripes on dorsum and flank absent; flanks largely pigmented with silver white; throat in adult males dark brown;undersides without dark patterns; outer metatarsal tubercle absent; supratympanic fold distinct; webbing between toes basal and poorly developed; tibiotarsal articulation adpressed limb reaching level of tympanum.The new species is divergent from all other congeners based on 16S rRNA gene sequences (3.0%-7.7%).Data on the natural history and male advertisement calls of the new species are provided.Following the IUCN Red List Categories and Criteria, we propose the new species to be listed as Vulnerable B1ab (iii).Frogs in the microhylid genusMicrylettaare widely distributed from northeast India, Myanmar, southern China, including Taiwan and Hainan islands, Indochina and through Peninsular Malaysia to Sumatra in Indonesia in the south (Frost, 2020;Suwannapoom et al., 2020) (Figure 1A).Currently, eight species are recognized in the genus, includingM.aishaniDas,Garg, Hamidy, Smith & Biju,M.dissimulansSuwannapoom,Nguyen, Pawangkhanant, Gorin, Chomdej, Che & Poyarkov,M.erythropoda(Tarkhnishvili),M.nigromaculataPoyarkov,Nguyen, Duong, Gorin & Yang,M.lineata(Taylor),M.inornata(Boulenger),M.steinegeri(Boulenger), andM.sumatranaMunir, Hamidy, Matsui, Kusrini & Nishikawa (Suwannapoom et al., 2020; Zug & Mulcahy, 2020).Recent phylogenetic studies have indicated thatM.inornatasensu stricto is restricted to the island of Sumatra, while the populations ofM.inornatasensu lato are widely reported from mainland Southeast Asia and contain several undescribed paraphyletic lineages with respect to other named taxa, suggesting that species diversity within the genus remains underestimated(Das et al., 2019; Munir et al., 2020; Poyarkov et al., 2018;Suwannapoom et al., 2020).
Lau & Chan (2012) reported the first record ofMicrylettaspecies from Hainan Island in China based on specimens collected in Yinggeling National Nature Reserve; they allocated this population toM.inornatalike all older accounts regardingMicrylettaspecies in the region (see Fei et al., 2009;Mathew & Sen, 2010; van Sang et al., 2009 as examples).However, we noted that specimens ofMicrylettafrom Hainan have a spotless dorsum and a relatively larger body size, and thus differ significantly fromM.inornatasensu stricto from Sumatra (Alhadi et al., 2019; Boulenger, 1890; Taylor, 1962).To clarify the taxonomic status ofMicrylettaspecies from Hainan, we examined newly collected specimens from the island incorporating morphological and molecular analyses.Results revealed that the HainanMicrylettaspecies indeed differs fromM.inornatasensu stricto and all other known congeners by a combination of morphological characters and molecular divergences, and thus represents a new species,which we describe herein.
A total of five specimens were collected during fieldwork in Hainan between 2012 and 2015.The geographic distribution ofMicrylettain Hainan is shown in Supplementary Figure S1.Specimens were fixed and preserved in 80% ethanol and deposited at the herpetology collection of Kadoorie Farm and Botanic Garden (KFBG) in Hong Kong, China.Measurements were taken to the nearest 0.1 mm with digital calipers.The descriptions of morphological characteristics followed Fei et al.(2009) and Poyarkov et al.(2018).Due to the high likelihood of undiagnosed diversity within the genus, where available, we relied on original species descriptions and topotypic materials for comparative morphological data analysis of all eight named taxa ofMicryletta(see Supplementary Methods for details).Advertisement calls were recorded in the field using aTASCAM DR-40 digital sound recorder (44.1 kHz sampling rate and 24 bit encoding), then analyzed with Raven Pro v.1.5 software (see Supplementary Methods for details).Descriptions of acoustic characteristics followed Köhler et al.(2017).
For reconstructing the phylogeny ofMicryletta, we extracted total genomic DNA and amplified and sequenced a 1 088 bp fragment of the mitochondrial 16S rRNA gene.DNA extraction and amplification protocols followed Poyarkov et al.(2018)and Suwannapoom et al.(2020) and are detailed in the Supplementary Methods.
Genealogical relationships ofMicrylettawere inferred using maximum-likelihood (ML) and Bayesian inference (BI)approaches (see Supplementary Methods).In total, 16S rRNA gene sequences for 28 Microhylinae specimens were included in the final analysis, representing all eight currently recognized nominal species ofMicrylettaand three outgroup taxa(Mysticellus franki,Uperodon systoma,Kaloula pulchra) (see Supplementary Table S1).Several undescribed lineages ofMicrylettareported in earlier phylogenetic studies (Das et al.,2019; Poyarkov et al., 2018; Suwannapoom et al., 2020) were not included in the analysis to avoid confusion and maintain simplicity.Pairwise sequence divergence (uncorrectedP-distances) was calculated using MEGA v6.0.6 (Tamura et al.,2013).
The ML and BI analyses resulted in essentially identical and generally well-resolved topologies, which only differed in two unsupported nodes (Figure 1B).Our updated phylogenetic tree agreed well with earlier phylogenies of the genusMicryletta(Poyarkov et al., 2018; Das et al., 2019;Suwannapoom et al., 2020), and suggested the following set of genealogical relationships.The first branching event inMicrylettaradiation separated a poorly supported clade(76/0.64; hereafter node support values are given as ML bootstrap (ML BS)/BI posterior probability (BPP), respectively)joiningM.sumatranafrom southern Sumatra andM.dissimulansfrom southern Thailand.Micryletta nigromaculatafrom northern Vietnam formed a sister clade with respect to all other remaining species ofMicryletta(71/0.91).Micryletta erythropodafrom southern Vietnam andM.cf.lineatafrom Peninsular Thailand formed a well-supported clade (99/1.0),with genealogical relationships with respect toM.aishanifrom northeastern India andM.inornatasensu stricto from northern Sumatra not supported (Figure 1B).The Hainan population ofMicrylettaformed a distinct clade (100/1.0), strongly supported as a sister lineage with respect toM.steinegerisensu stricto from Taiwan Island, China (96/1.0) (Figure 1B).
Genetic divergences between theMicrylettasp.from Hainan Island and all other congeners ranged fromP=3.0%(withM.steinegerisensu stricto) toP=7.7% (withM.sumatrana) (Supplementary Table S2).These values were equal to or higher than the formalP=3% threshold for the 16S rRNA gene, which is considered an indicator of species-level divergence in Anura (Vieites et al., 2009).
Thus, based on phylogenetic position, genetic divergence,stable morphological, and chromatic differences (see below),we hypothesize that the Hainan population ofMicrylettarepresents a discrete diagnosable lineage showing no evidence of reticulation with any other currently recognized congener, and as such, should be accorded species status, as formally described below.
Taxonomic account
Micryletta immaculata sp.nov.(Figure 1; Supplementary Figures S2-4; Table 1)
Chresonymy:Micryletta inornata— Lau & Chan, 2012: 54;Plate 1 (Yinggeling National Nature Reserve, Hainan, China).
Holotype:KFBG 14271, adult male from Nanlang Ranger Station, Exianling Nature Reserve, Exian Village, Dongfang City, Hainan, China (coordinates N19°01 ′02.41 ″,E109°06′27.28″, elevation ca.630 m a.s.l.), collected by J.H.Yang on 10 September 2015.
Paratypes:KFBG 14270 and 14272, two adult males,collection data identical to holotype.KFBG 14105, adult female from same locality as holotype, collected by Chao Zhang, Philip Lo, and Jay Wan on 23 July 2012.KFBG 14106,adult female from Jianfengling National Forest Park, Ledong County, Hainan, China (coordinates N18°43 ′48.20 ″,E108°50′07.64″, elevation ca.810 m a.s.l.), collected by J.H.Yang and Sheng Zheng on 28 August 2012.
Etymology:The specific epithetimmaculata(meaning“spotless”, “immaculate”) is a Latin singular feminine adjective in the nominative case and refers to the absence of distinct dark spots or stripes on the dorsum and lateral sides of the body in the new species.We propose “Hainan Paddy Frog” for the common English name and “海南小姬蛙” (Hainan Xiao Ji Wa) for the common Chinese name of the new species.
Diagnosis:The new species is assigned to the genusMicrylettabased on its possession of the following morphological characters, which are considered diagnostic for the genus: body size small; vomerine teeth absent; dorsal skin smooth and scattered with flat and small tubercles; forelimbs thin and slender; tympanum roughly distinct; subarticular tubercles on fingers and toes large and very prominent;supernumerary tubercles on palm present and distinct; three metacarpal tubercles; tips of digits round, not expanded into discs, with no peripheral grooves; distinct dermal ridges present under 3rdto 4thtoes (Dubois, 1987; Fei et al., 2009).
Micryletta immaculatasp.nov.can be distinguished from its congeners by a combination of the following characters:medium-sized within genus (SVL 23.3-24.8 mm in males,n=3; 27.7-30.1 mm in females,n=2); dorsum bronze brown to reddish brown in life; dark brown spots and stripes on dorsum and flank absent; flanks largely pigmented with silver white;throat in adult males dark brown; undersides without dark patterns; outer metatarsal tubercle absent; supratympanic fold distinct; webbing between toes basal and poorly developed;tibiotarsal articulation adpressed limb reaching level of tympanum.
Description of holotype:SVL 24.5 mm; habitus relatively slender; head small and triangular, slightly wider (HW 7.7 mm)than long (HL 7.1 mm); snout abruptly rounded in dorsal view and slightly acuminate in profile, projecting beyond margin of lower jaw; eyes moderate size, slightly protuberant, eye diameter (EL 3.0 mm) about equal to snout length (SL 3.1 mm) and interorbital distance (IOD 2.8 mm).Top of head flat, canthus rostralis rounded and distinct; loreal region weakly concave; nostril round, lateral, below canthus rostralis,closer to tip of snout than to eye; interorbital distance (IOD 2.8 mm) greater than internarial distance (IND 2.0 mm), and much greater than upper eyelid width (UEW 1.7 mm).Pineal spot absent; tympanum round and small (TMP 1.3 mm) and roughly distinct; supratympanic fold distinct and curved,extending from posterior corner of eye to arm insertion.Choanae elongated and oval-shaped, widely spaced;vomerine teeth absent; tongue roundly spatulate, with no notch at posterior tip, and free behind for about half its length.
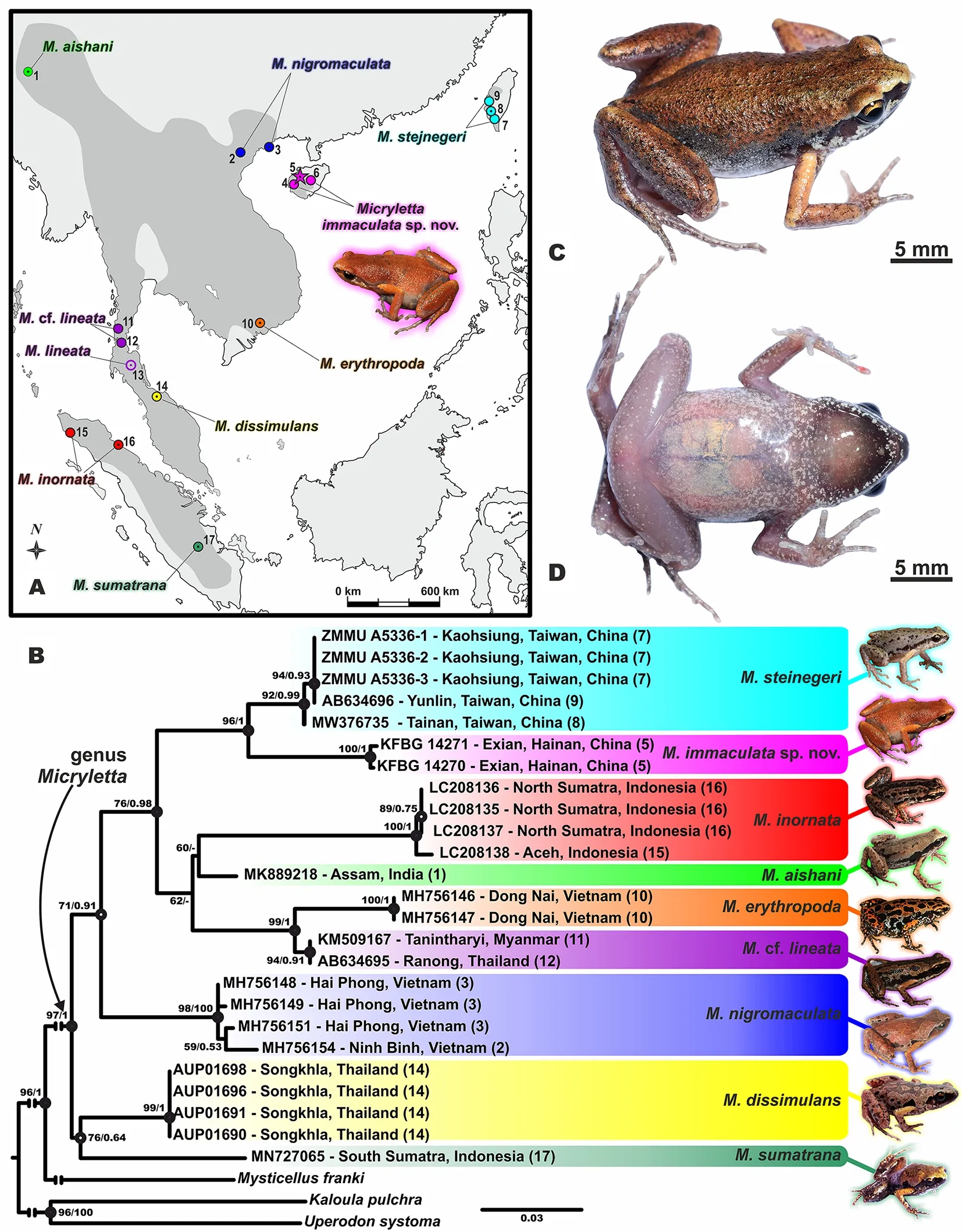
Figure 1 Phylogenetic relationships and distribution of genus Micryletta and holotype of Micryletta immaculata sp.nov.(KFBG 14271) in life

Table 1 Measurements (in mm) of type specimens of Micryletta immaculata sp.nov.
Forelimbs slender (FLL 17.1 mm), lower arm and hand length (LAHL 12.9 mm) more than a half of snout-vent length(HAL/SVL ratio 0.53).Fingers slender, free of webbing,rounded in cross-section, no lateral fringes on fingers present;first finger well-developed, second finger slightly shorter than fourth, relative finger lengths: I<II<IV<III; tips of fingers round and not dilated, about equal in width of basal phalanges;subarticular tubercles on fingers distinct, rounded and prominent, formula 1, 1, 2, 2; supernumerary tubercles on palm present and strong; three metacarpal tubercles, inner one smallest (0.5 mm in diameter) and separated from median one, median one (0.7 mm in diameter) bordering and almost directly in front of large outer one (0.9 mm in diameter); 2-3 tiny tubercles present between supernumerary and metacarpal tubercles; nuptial pad absent.
Hindlimbs slender and long (HLL 38.6 mm), mostly more than two times longer than forelimbs (HLL/FLL ratio 2.26); tibia(TL 11.1 mm) smaller than one-third of hindlimb length, heels just overlapping when thighs held at right angles to body;tibiotarsal articulation of adpressed limb reaching level of tympanum; foot (FL 12.3 mm) slightly longer than tibia.Relative toe lengths: I<II<V<III<IV; tarsus smooth, inner tarsal fold absent; tips of toes round and weakly dilated, slightly wider than those of fingers; webbing between toes basal and poorly developed, webbing formula (number of phalanges free of web): I n.a.-n.a.II 2-3¾ III 3-4½ IV 4¾-3 V; subarticular tubercles on toes similar to those on fingers but smaller,prominent, rounded, formula 1, 1, 2, 3, 2; distinct dermal ridges present under 2ndto 4thtoes; inner metatarsal tubercle oval, large, and prominent, length (IMTL 0.8 mm) shorter thanhalf of first toe length (1TOEL 2.4 mm); outer metatarsal tubercle absent.
Dorsal skin smooth above, scattered with tiny and flat tubercles on dorsum of body, flanks, and hindlimbs; subtle longitudinal median ridge present on dorsum (visible in life;nearly invisible in preservation); lateral sides of head smooth;ventral skin of body and limbs smooth; vent area smooth with few low tubercles in cloacal region.
Coloration of holotype in life:Dorsum of body and limbs bronze brown, elbow and upper arms with distinct lighter coloration; tiny tubercles on dorsum of body and hindlimbs relatively indistinct dark brown; flanks largely pigmented with silver white; dark brown spots or stripes on dorsum of body and flanks absent; dark brown cross-bars or speckling on dorsal surfaces of limbs absent.Lateral head dark brown;irregular white spots present along upper lips; lower margin of supratympanic fold black.Ventral surface transparent, with small and irregular white marbling patterns, those on chest and lateral belly relatively distinct and large, those on center of belly, throat, and ventral surfaces of limbs fewer and smaller;throat notably darker; margins of lower jaw with distinct irregular white marbling.Pupil round; iris bicolored, with upper third bronze and lower two-thirds dark brown.
Coloration of holotype in preservative:Colors greatly faded; ventral side dull white with light gray marbling on chest and lateral sides of belly; chin distinctly grayish black.
Variation and sexual dimorphism:Individuals of the type series are generally similar in appearance.Variations in size and body proportions are given in Table 1.All types showed no dark brown patterns on dorsum or limbs.Notable sexual dimorphism: females larger and thicker than males (SVL 27.7-30.1 mm in two females vs.23.3-24.8 mm in three males); chin notably darker in all three males; brighter reddish coloration on dorsum in life in females, relatively darker dorsum color in males (Supplementary Figures S2, 3).Gravid female paratype KFBG 14106 was distended with eggs(Supplementary Figure S3E, F).
Comparison:Micryletta immaculatasp.nov.differs fromM.aishaniby relatively larger body (SVL 27.7-30.1 mm in females (n=2) vs.25.6-27.3 mm in females (n=4)); snout abruptly rounded in dorsal view and slightly acuminate in lateral view (vs.snout shape nearly truncate in dorsal view and acute in lateral view); webbing between toes basal and poorly developed (vs.absent); dorsum without dark patterns(vs.with several blackish brown spots present on posterior parts of back and near groin); lateral side of body without dark patterns (vs.with prominent blackish brown streak extending from tip of snout to lower abdomen, see Supplementary Figure S4A); tibiotarsal articulation adpressed limb reaching level of tympanum (vs.reaching to armpit).
The new species can be distinguished fromM.inornatasensu stricto from Sumatra by relatively larger body (SVL 23.3-24.8 mm in males (n=3), 27.7-30.1 mm in females (n=2)vs.16.8-20.5 mm in males (n=3), 19.5 mm in female (n=1));webbing between toes basal and poorly developed (vs.absent); dorsum without dark blotches/stripes (vs.with irregular blackish brown blotches and discontinuous lateral blackish brown streak, see Supplementary Figure S4D); flanks without black spots (vs.with blackish brown spots); tibiotarsal articulation adpressed limb reaching level of tympanum (vs.reaching to eye).
Micryletta immaculatasp.nov.can be diagnosed fromM.lineataby relatively larger body (SVL 23.3-24.8 mm in males(n=3), 27.7-30.1 mm in females (n=2) vs.19.0-19.2 mm in males (n=2), 19.2-22 mm in females (n=2)); webbing between toes basal and poorly developed (vs.absent); dorsum without dark blotches/patterns (vs.with three straight continuous or broken lines, see Supplementary Figure S4E); flanks without black spots (vs.with black stripe from axilla to groin);tibiotarsal articulation adpressed limb reaching level of tympanum (vs.reaching to eye).
The new species differs fromM.dissimulansby relatively larger body (SVL 23.3-24.8 mm in males (n=3), 27.7-30.1 mm in females (n=2) vs.20.3-22.4 mm in males (n=7),24.4-26.7 mm in females (n=2)); supratympanic fold distinct(vs.indistinct and flat); webbing between toes basal and poorly developed (vs.absent); dorsum without dark patterns(vs.with merging irregular-shaped brown blotches edged in beige, see Supplementary Figure S4B); black spots on lateral sides of body absent (vs.large black spots on flanks and axillary and inguinal areas present); white spots on lips present (vs.absent).
Micryletta immaculatasp.nov.differs fromM.erythropodaby relatively smaller body (SVL 23.3-24.8 mm in males (n=3)vs.up to 30 mm in males); large and distinct dark brown spots/stripes on dorsum and flank absent (vs.present, see Supplementary Figure S4C); dark patterns on dorsal surfaces of limbs absent (vs.dorsal surfaces of limbs usually bearing dark marbled speckling); undersides without dark patterns (vs.with relatively distinct dark and light marbled speckling); outer metatarsal tubercle absent (vs.present).
The new species differs fromM.nigromaculataby relatively larger body (SVL 23.3-24.8 mm in males (n=3), 27.7-30.1 mm in females (n=2) vs.18.5-23.0 mm in males (n=18),24.2-25.9 mm in females (n=3)); webbing between toes basal and poorly developed (vs.absent); dorsum without dark blotches/patterns (vs.with dark brown irregular hourglassshaped pattern and two large dark inguinal spots); flanks without dark spots (vs.with dark patches or spots, see Supplementary Figure S4F); irregular white spots along upper lips present (vs.absent); throat in males dark brown (vs.whitish with light-gray marbling).
Micryletta immaculatasp.nov.differs fromM.sumatranaby relatively larger body (SVL 23.3-24.8 mm in males (n=3),27.7-30.1 mm in females (n=2) vs.17.4 mm in male (n=1),22.8 mm in female (n=1)); dark cross bands on tibia and tarsus absent (vs.present); black spots on lateral sides of body absent (vs.black spots on flanks and axillary and inguinal areas present); undersides without mottling patterns(vs.with dark brown and cream mottling); tibiotarsal articulation adpressed limb reaching level of tympanum (vs.reaching to eye).
Micryletta immaculatasp.nov.is most similar to its putative sister speciesM.steinegerifrom Taiwan, China, but can still be distinguished by supratympanic fold distinct (vs.weak and indistinct); dorsum bronze brown to reddish brown without dark patterns in life (vs.dorsum dark gray to violet in life with irregular dark blotches or speckles, see Supplementary Figure S4G, H); dark patterns on dorsal surfaces of limbs absent (vs.dark brown spots on dorsal surfaces of limbs present);undersides without dark patterns (vs.with grayish white and brown spots).
Natural history and distribution:Three adult males (KFBG 14270-14272) from Exianling Nature Reserve were found simultaneously calling at night in accumulated water on limestone surfaces on the forest floor (see Supplementary Figure S5A); many tadpoles were found in the water but were not collected.An adult gravid female (KFBG 14106) from Jianfengling National Forest Park was found perched on the forest floor by a vehicular road at night.
In addition to the above two localities with voucher specimens, the new species was also recorded from the following three locations on Hainan Island: (1) many males and females were found congregating in a seasonal puddle in the forest of Zhufeng in Yinggeling National Nature Reserve(elevation ca.1 470 m a.s.l.) at daytime on 31 May 2005:individuals were heard calling in nearby shrubs or found floating in water puddles; patches of eggs were observed floating on the water surface (see Supplementary Figure S5B);(2) male advertisement calls were heard and two calling males were found perched on leaves of undergrowth in a seasonal shrubby wetland (abandoned paddy field) at the forest edge of Yaxing Village in Yinggeling National Nature Reserve(coordinates N19°01 ′24.20 ″, E109°24 ′43.06 ″, elevation ca.340 m a.s.l.) at night on 29 August 2012; (3) male advertisement calls were heard from a shrubby wetland by the edge of a rubber plantation in Baoxiu Village of Jiaxi Nature Reserve (coordinates N18°49 ′40.83 ″, E109°09 ′05.09 ″,elevation ca.160 m a.s.l.), Ledong County on 28 July 2016.
The new species is currently recorded from the abovementioned five localities on Hainan Island (see Supplementary Figure S1) and inhabits a relatively wide elevational range(160 m to 1 470 m) in montane forest areas.It breeds during the rainy season from May to September, but males only appear for calling and breeding after heavy rains in accumulated water in forests and along the forest edge.Due to its perfect cryptic coloration under leaf litter and elusive habit, the new species is difficult to detect in the wild.
Advertisement call:Only a short recording (15 s) of six advertisement calls in a series was recorded from the paratype KFBG 14270, under an ambient air temperature of 28 °C (see Supplementary Figure S6).The advertisement call contained a single long note of 0.918-1.800 s duration(1.410±0.364 s,n=6), containing 63-127 pulses (98.8±27.5 pulses/call,n=6).Pulse intervals were nearly uniform from the beginning to the end of the call, with a pulse rate of 69.8±2.66(n=6) pulses/s.Calls were repeated in a series at a rate of ca.0.41 calls/s, with an irregular call interval duration of 0.680-1.516 s (1.027±0.307 s,n=5).The dominant frequency of calls ranged from 4.57 to 4.91 kHz (4.81±0.127,n=6).
Conservation status:In Hainan, outside of protected areas,natural lowland habitats are seriously affected by the expansion of small-scale rubber and orchard plantations (e.g.,banana andArecapalm), along with associated agricultural pollution from herbicides and pesticides.During field work, we observed habitat loss and degradation in natural landscapes adjoining the Exianling and Jiaxi nature reserves, where the new species has been recorded.
Following the IUCN Red List Categories and Criteria (IUCN,2012), we proposeMicryletta immaculatasp.nov.be listed as Vulnerable B1ab (iii) as the extent of its occurrence is <20 000 km2, with <10 known locations, and its habitat continues to decline in both extent and quality, as mentioned above.Therefore, the strengthening of habitat protection and improvement of management are required in lowland forest areas on Hainan Island to conserve this island-endemic amphibian and other associated species.
NOMENCLATURAL ACTS REGISTRATION
The electronic version of this article in portable document format represents a published work according to the International Commission on Zoological Nomenclature (ICZN),and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone (see Articles 8.5-8.6 of the Code).This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN.The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/.
Publication LSID:
urn:lsid:zoobank.org:pub:88CDB1FD-F12B-45CB-B72B-37A469717E91
Micryletta immaculataLSID:
urn:lsid:zoobank.org:act:4E57599F-3647-4D7F-A086-99818FBD0214
SCIENTIFIC FIELD SURVEY PERMISSION INFORMATION
Permission for field surveys in Hainan was granted by the Hainan Provincial Department of Forestry.
SUPPLEMENTARY DATA
Supplementary data to this article can be found online.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS" CONTRIBUTIONS
Both authors designed the study, performed the field work and data analysis, wrote the manuscript, and read and approved the final version of the manuscript.
ACKNOWLEDGMENTS
This study was supported by the Kadoorie Farm and Botanic Garden of Hong Kong.We thank the Hainan Provincial Department of Forestry for permission to conduct fieldwork.We are also grateful to Sheng Zheng, the forestry rangers of Exianling Nature Reserve, Yinggeling National Nature Reserve, and Jiaxi Nature Reserve, as well as colleagues from KFBG for their great help during field surveys; and Bosco P.L.Chan and Jay Wan for providing valuable field records and photos of the new species in Hainan.We also thank Chung-Wei You and P.Pawangkhanant for providing photos ofM.steinegeriandM.dissimulans, respectively.
猜你喜欢
基层中医药(2021年6期)2021-11-02
新教育(2018年8期)2018-08-29
小学生作文(中高年级适用)(2018年4期)2018-05-14
南方周末(2018-05-03)2018-05-03
音乐天地(音乐创作版)(2017年10期)2018-01-25
中国公路(2017年19期)2018-01-23
现代园艺(2017年22期)2018-01-19
中国篆刻(2017年7期)2017-09-05
下一代英才(酷炫少年)(2016年4期)2016-11-24

- Zoological Research的其它文章
- Flexible breeding performance under unstable climatic conditions in a tropical passerine in Southwest China
- Comparative transcriptomics highlights convergent evolution of energy metabolic pathways in group-living spiders
- Long-term trends in a forest ungulate community: park establishment increases numbers, but poaching is a constant threat
- Molecular phylogeny of the genus Muntiacus with special emphasis on the phylogenetic position of Muntiacus gongshanensis
- Parasitism is always costly to the host
- Yunnanilus chuanheensis, a new loach species(Cypriniformes: Nemacheilidae) from the upper Lixianjiang River in Yunnan, China