
野生老芒麦苗期耐旱性品种筛选及鉴定
2021-06-18 04:02王传旗刘文辉张永超周青平
草业科学 2021年5期
王传旗,刘文辉,张永超,周青平
(青海大学畜牧兽医科学院,青海西宁810016)
干旱是一个长期存在的全球性问题,受到各国科研工作者的极大关注[1]。青藏高原平均海拔4 000 m以上,被称为地球“第三极”,是我国草地的主要分布区,也是全世界所关注的生态环境脆弱的地区之一[2-3]。青藏高原大部分地区处于半干旱气候条件下,由于区域性、季节性、年际间及作物生长期内的降水变幅较大,植物常因周期性或难以预测性干旱而大面积减产。为从根本上减轻或避免干旱灾害,首要解决的问题是培育出具有高耐旱性的植物品种。因此,研究植物干旱适应机理,积极探索和选育耐旱植物新品种具有重要的科学价值和现实意义。
老芒麦(Elymus sibiricus)属禾本科披碱草属(Elymus),为多年生草本植物。老芒麦兼具经济和生态价值,不仅草茎柔软、富含粗蛋白,还具有抗旱、耐寒、抗风沙、耐盐碱等优良特性;此外,老芒麦还能够显著改善表层土壤理化性质,对我国贫瘠荒地土壤的培育具有重要的应用价值[4-6]。目前,关于老芒麦抗旱方面的研究主要集中在萌发期和苗期。老芒麦种子萌发期干旱适应机理研究中,研究者多采用的是高分子渗透剂聚乙二醇(polyethylene glycol,PEG-6000)配制成不同浓度的溶液模拟干旱胁迫环境。白史且等[7]对采自川西北高原的9份野生老芒麦种子进行发芽率(germination percentage,GP)、发芽势(germination vigor,GV)、萌发指数(germination index,GI)等指标的测量,认为萌发指数可作为老芒麦抗旱性鉴定的初级指标。王传旗等[8]研究表明,15%PEG-6000能够显著抑制老芒麦种子的发芽率、发芽势、发芽指数和活力指数(vigor index,VI)。老芒麦苗期是其干旱适应机理研究的重要生育时期,研究者多采用盆栽控水法且在老芒麦3~5叶期开始进行干旱胁迫。研究表明,干旱胁迫下,植物通常会主动积累各种渗透调节物质以降低水势,以便于从干旱土壤中吸收水分[9]。杨满业等[10]研究表明,老芒麦叶片相对含水量和叶绿素含量下降幅度与干旱胁迫时间呈正相关关系,而丙二醛(malondialdehyde,MDA)含量、相对电导率(relative conductivity,REC)、过氧化物酶(peroxidase,POD)、超氧化物歧化酶(superoxide Dismutase,SOD)活性、脯氨酸(proline,Pro)和可溶性糖(soluble sugar,SS)含量增加幅度与干旱胁迫时间呈正相关关系。张卫红等[11]研究表明,随干旱胁迫时间的增加,老芒麦幼苗株高(plant height,PH)、植株含水量(plant water content,PWC)呈下降趋势。目前文献中尽管对老芒麦耐旱性研究较多,但系统全面评价老芒麦苗期耐旱性的研究相对较少,尤其是缺乏针对青藏高原区域野生老芒麦耐旱性种质资源筛选及鉴定的工作。本研究正是从这方面出发,以青藏高原的优势乡土植物老芒麦为试验材料,采用盆栽控水法,对供试老芒麦种质资源苗期耐旱性进行筛选与鉴定。然而,鉴于目前国内外研究者对于植物的耐旱性鉴定方法及抗旱指标的研究工作仍处于探索阶段。因此,本研究试图用多个指标来综合反映老芒麦苗期的耐旱能力,以期筛选出耐旱性较强的老芒麦材料,为老芒麦耐旱性新品种的选育和野生老芒麦种质资源的开发利用提供科学依据。
1 材料与方法
1.1 试验材料
19种野生老芒麦分别在2015年和2016年采集于青海省境内,风干后于4℃下保存。同时,试验选用青藏高原区域在栽培草地建植、实施草地补播改良和草原生态恢复中被广泛利用的青牧1号老芒麦和川草2号两种老芒麦品种作为参比材料。试验材料来源如表1所列。
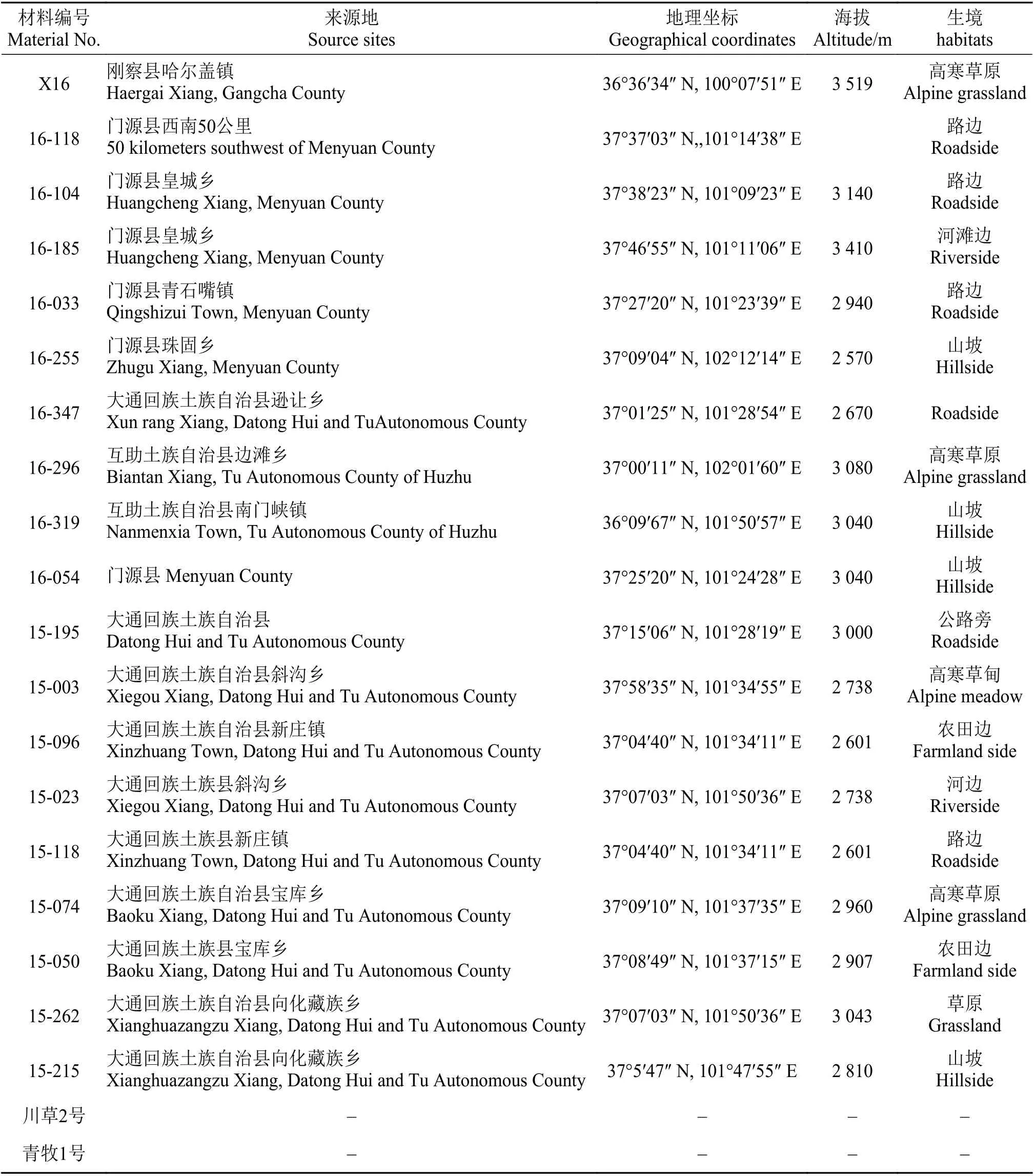
表1 老芒麦材料编号及来源状况Table 1 Material number and source status of wild and cultivated Elymus sibiricus
1.2 试验地点和时间
试验在青海大学畜牧兽医科学院青藏高原优良牧草种质资源利用重点实验室智能玻璃温室内进行。2020年6月30日播种,8月18日开展干旱胁迫试验,老芒麦苗期干旱胁迫试验共历时50 d。
1.3 试验方法
随机挑选大小均匀的老芒麦种子,用70%的酒精消毒5 min。老芒麦育苗在玻璃温室的栽培池上进行,室内温度18~25℃,相对湿度45%,光照强度1 000~1 300 lx。
老芒麦种子撒播在口径30 cm、高19 cm的花盆中,并覆土0~0.5 cm厚,每盆播量为200粒。培养基质质量由营养土、蛭石和珍珠岩按16꞉3꞉1的比例混合而成,每盆3 kg,每盆最大持水量为27.46%。当老芒麦苗龄为二叶期时进行间苗,保证每盆有100株健壮幼苗。待苗龄为四叶期时开始进行中度干旱胁迫处理(土壤相对含水量为田间持水量的45%~50%)[12-13]。共21份材料,每份材料种植8盆(4盆对照组,4盆处理组),共168盆。干旱胁迫期间,对照组每隔1 d浇灌一次;处理组每隔3 d浇灌一次,浇灌量为每盆称重的失水量,每盆约50 g,且浇灌时间均在当日的18:00− 20:00。
综合性的抗旱生理指标由株高、相对生长速率、相对成活率、叶面积、根系干物质积累率、叶绿素和相对电导率组成。其中,株高、叶面积、相对存活率和地下生物量从干旱胁迫开始起每5 d测量一次,叶绿素和相对电导率每10 d测量一次。干旱胁迫处理20 d后,老芒麦相对存活率按每10 d 统计一次。
1.4 测定指标与方法
株高:从处理组中随机选取30株成苗定株,用直尺近地面测量老芒麦绝对株高,取平均值。
相对生长速率(relative growth rate,RGR):处理组株高增长速率/对照组株高增长速率。
叶面积(leaf area,LA):从处理组中随机选取10株成苗,用叶面积仪测量叶面积,测量的单叶为植株顶端向下数第3片叶子,取平均值。
相对成活率(relative survival percentage,RSP):干旱处理下存活株数/对照组处理下存活株数×100%[14]。
地下生物量:从处理组中随机选取5株健壮成苗,冲根后用快速滤纸吸干根系表面水分称重,得其鲜重;然后在105℃烘箱内杀青15 min,再在75℃下烘至恒重,得其干重[15]。
干物质积累率(dry matter accumulation percentage,DMAP):地下部分干重/地下部分鲜重×100%。
叶绿素(chlorophyll,Chl)含量采用丙酮浸提法测定[16],相对电导率采用电导法测定[17],二者的测定材料均为老芒麦地上茎叶部分。
1.5 耐旱性综合评价
试验利用模糊数学中的隶属函数法对21份老芒麦材料苗期的耐旱性进行综合评价。隶属函数公式:U(Xi)=(Xi−Xmin)/(Xmax−Xmin)。式中:U(Xi)为隶属函数值,Xi为处理水平某指标测定均值,Xmin和Xmax为某一指标的最小值和最大值。根据隶属函数公式求得试验材料指标函数值,然后将每份材料指标的函数值相加求平均值,得到每份材料的平均隶属函数值。函数平均值划分标准:0.6~0.7属强抗,定为Ⅰ级;0.4~0.6属较抗,定为Ⅱ级;0.3~0.4属弱抗,定为Ⅲ级;0.3以下属不抗,定为Ⅳ级[18]。
1.6 数据分析
利用SPSS 19.0统计分析软件对所测数据进行分析。采用Tukey’s HSD检验,对老芒麦材料间和处理间的差异性进行单因素方差分析和Duncan多重比较分析。所有数据在分析前要满足正态分布和方差齐性。
2 结果与分析
2.1 干旱胁迫下苗期老芒麦形态特征
供试的21份老芒麦材料处理间的株高及其相对生长速率存在差异(表2)。随着干旱胁迫天数的增加,老芒麦材料株高增长减缓,处理组老芒麦株高均低于对照组。供试的21份老芒麦材料中,门源县的16-118老芒麦和大通县的16-347、15-195、15-262老芒麦及参比材料川草2号老芒麦的相对生长速率较大,其值分别为87.50%、86.84%、97.76%、88.89%和89.47%;大通县的15-096、15-215老芒麦和门源县的16-185老芒麦相对生长速率较小,其值分别为48.48%、43.75%和47.50%。
随着干旱胁迫天数的增加,处理组老芒麦叶片面积呈上升趋势,但处理组老芒麦单叶面积均低于对照组(表3)。干旱胁迫第5天,与对照相比,门源县的16-033老芒麦单叶面积显著下降(P<0.05);干旱胁迫第10天,与对照相比,门源县的16-054老芒麦单叶面积显著下降(P<0.05);干旱胁迫第15天,与对照相比,刚察县的X16老芒麦和川草2号老芒麦单叶面积显著下降(P<0.05);干旱胁迫第20天,与对照相比,互助县的16-319老芒麦和大通县的15-023老芒麦单叶面积显著下降(P<0.05)。干旱胁迫下,青牧1号老芒麦单叶面积均低于对照组但无显著差异(P> 0.05)。供试的21份老芒麦材料中,大通县的15-215老芒麦单叶面积增幅最大(为对照组的97.95%),门源县的16-054老芒麦单叶面积增幅最小(为对照的50.37%)。

表2 干旱胁迫时间对老芒麦株高的影响Table2 Effectsof drought stresstime on plant height of Elymus sibiricus cm

续表2Table 2(Continued)cm
随着干旱胁迫天数的增加,供试的21份老芒麦材料根系干物质积累率逐渐增加(表4)。干旱胁迫第5天,刚察县的X16老芒麦干物质积累率较对照组显著增加(P<0.05),互助县的16-296、16-319老芒麦干物质积累率较对照组显著增加,门源县的16-118、16-104、16-054老芒麦干物质积累率较对照组均显著增加,大通县的16-347、15-195、15-118、15-023、15-050老芒麦干物质积累率较对照组均显著增加,参比材料青牧1号老芒麦和川草2号老芒麦干物质积累率较对照组显著增加。干旱胁迫第10天,与未胁迫相比,门源县的16-033、16-185、16-255老芒麦干物质积累率均显著增加,大通县的15-003、15-096老芒麦干物质积累率均显著增加。干旱胁迫第15天,与无胁迫相比,大通县的15-215、15-262、15-074老芒麦干物质积累率均显著增加。干旱胁迫下,供试的21份老芒麦材料中,门源县的16-255老芒麦和大通县的15-118老芒麦根系干物质积累率较大(为0.60和0.57),且分别是其对照组的142.86%和146.15%;门源县的16-033老芒麦根系干物质积累率最小(为0.39),是其对照组的95.12%。
随着干旱胁迫天数的增加,供试的21份老芒麦材料相对存活率均出现了不同程度的下降现象(表5)。干旱胁迫第15天,与对照组相比,刚察县的X16老芒麦、门源县的16-033老芒麦和互助县的16-319老芒麦材料相对存活率显著下降(P<0.05)。干旱胁迫第20天,与对照相比,门源县的16-054老芒麦和大通县的15-023、15-050、15-215老芒麦材料相对存活率显著下降。干旱胁迫第30天,与对照组相比,门源县的16-118、16-104、16-185、16-255老芒麦材料相对存活率显著下降,大通县的16-347、15-195、15-003、15-096、15-118、15-074、15-262老芒麦材料相对存活率显著下降,互助县的16-296老芒麦相对存活率显著下降,参比材料川草2号老芒麦和青牧1号老芒麦材料相对存活率显著下降(P<0.05),而刚察县的X16老芒麦、门源县的16-033老芒麦和大通县的15-215老芒麦相对存活率下降极显著(P< 0.01)。干旱胁迫第40天,与对照相比,互助县的16-319老芒麦、门源县的16-054老芒麦和大通县的15-003、15-023、15-050老芒麦及参比材料川草2号老芒麦相对存活率极显著下降(P<0.01)。干旱胁迫第50天,与对照组相比,门源县的16-118、16-104、16-185、16-255老芒麦相对存活率极显著下降,大通县的16-347、15-195、15-074、15-096、15-118、15-262老芒麦相对存活率极显著下降,互助县的16-296老芒麦和参比材料青牧1号老芒麦相对存活率极显著下降。供试的21份老芒麦材料中,大通县的15-195老芒麦相对成活率较高,即使干旱胁迫至第30天,其相对成活率仍高达66%,而刚察县的X16老芒麦和门源县的16-033老芒麦相对成活率较低,干旱胁迫至第30天,其相对成活率不足40%。

表3 干旱胁迫时间对老芒麦单叶面积的影响Table 3 Effectsof drought stresstime on single leaf area of Elymus sibiricus mm2

续表3Table 3(Continued)mm2
2.2 干旱胁迫下苗期老芒麦生理特征
充足供水条件下,老芒麦材料叶绿素含量均随着生长天数的增加而上升;干旱胁迫下,老芒麦材料叶绿素含量随着干旱胁迫天数的增加而下降(表6)。干旱胁迫第10天,与对照组相比,门源县的16-118老芒麦和大通县的15-195老芒麦叶绿素含量下降不显著(P>0.05),其他老芒麦材料下降显著(P<0.05);干旱胁迫第20天,与对照组相比,供试的21份老芒麦材料叶绿素含量均下降显著。干旱胁迫第20天,与干旱胁迫第10天相比,门源县皇城乡的16-104、门源县青石嘴镇的16-033、门源县的16-319和互助县南门峡镇的16-319老芒麦叶绿素含量显著下降。干旱胁迫第10天和第20天,供试的21份老芒麦材料中,互助县的16-319老芒麦叶绿素含量最低,分别为2.27和1.34 mg·L−1。
干旱胁迫下,供试的21份老芒麦材料相对电导率值随着生长天数的增加而上升(表6)。干旱胁迫第10天,与对照组相比,供试的21份老芒麦材料相对电导率值均无显著差异。干旱胁迫第20天,与对照组相比,刚察县的X16,互助县16-319,门源县的16-104、16-054、16-033、16-255,大通县的15-003、15-023、15-118、15-050、15-262、15-215老芒麦相对电导率值均显著上升,参比材料川草2号老芒麦和青牧1号老芒麦相对电导率值均显著上升。干旱胁迫第10天和第20天,供试的21份老芒麦材料中,门源县的16-033老芒麦相对电导率值最高,分别为67.26%和81.89%。
2.3 老芒麦耐旱性综合评价
使用隶属函数法对老芒麦材料的表型和生理指标进行了综合评价(表7)。供试的21份老芒麦材料函数平均值均介于0.331 7~0.603 3。其中,大通县的15-195老 芒 麦 在0.6~0.7,16-347、15-003、15-096、15-023、15-118、15-074、15-050、15-262、15-215老芒麦在0.4~0.6;刚察县的X16老芒麦在0.4~0.6;门源县的16-118、16-104、16-185、16-255、16-054老芒麦均在0.4~0.6,16-033老芒麦在0.3~0.4;互助县的16-296老芒麦在0.4~0.6,16-319老芒麦在0.3~0.4;参比材料川草2号老芒麦和青牧1号老芒麦均在0.4~0.6。
3 讨论
3.1 干旱胁迫下苗期老芒麦形态指标的变化特点
植物是固着在地面上的有机生物,由于其无法通过行动趋利避害,因而表型可塑性就显得尤为重要。株高、叶面积、干物质积累率和相对存活率等是评价植物抗逆性强弱的重要生长量指标[19-20]。研究表明,当植物受到环境胁迫时,比如水分、光照、营养胁迫等,许多表型响应受资源限制,常表现为消极生长[21]。
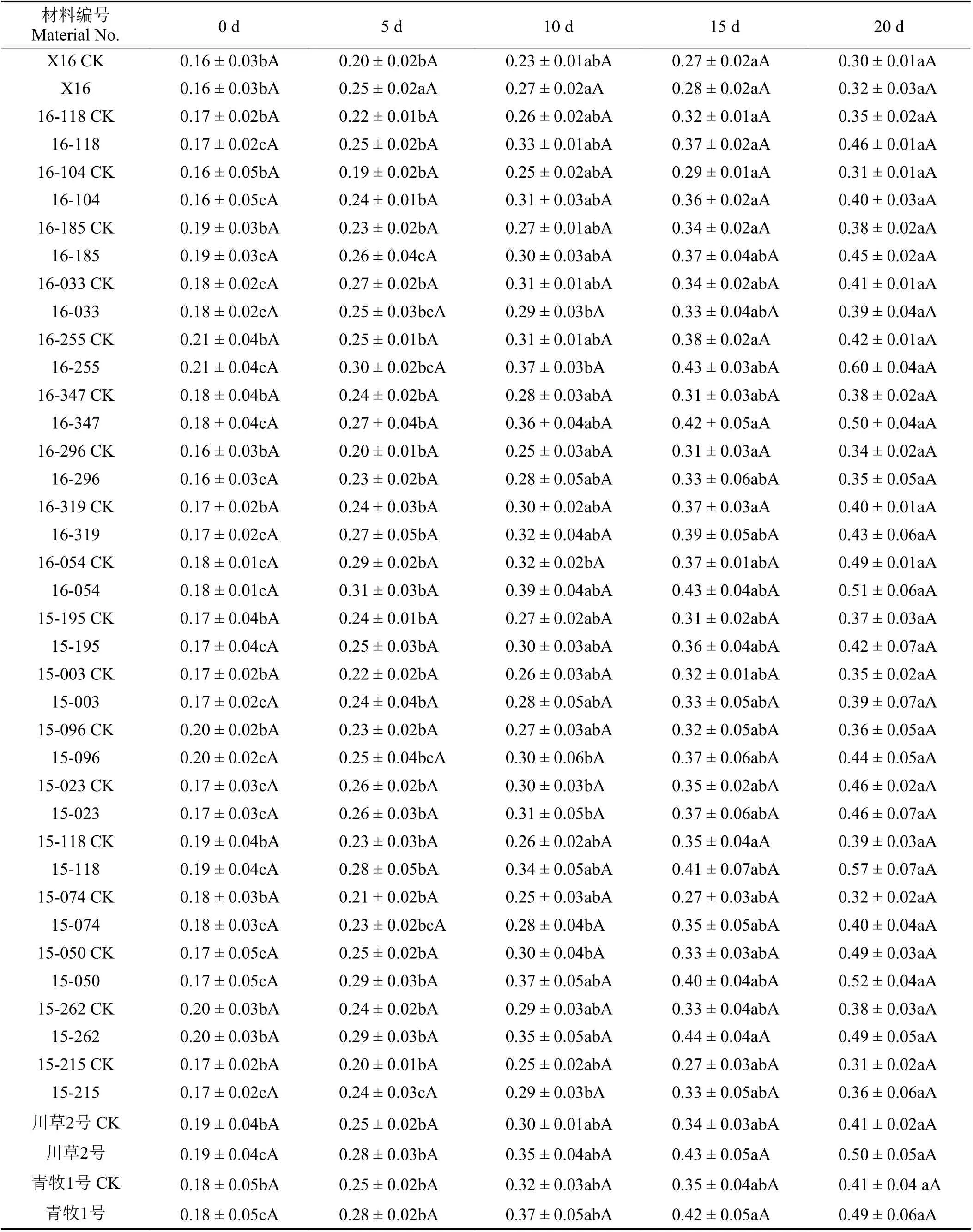
表4 干旱胁迫时间对老芒麦地下部分干物质积累率的影响Table4 Effectsof drought stresstime on dry matter accumulation percentage of underground parts of Elymus sibiricus%
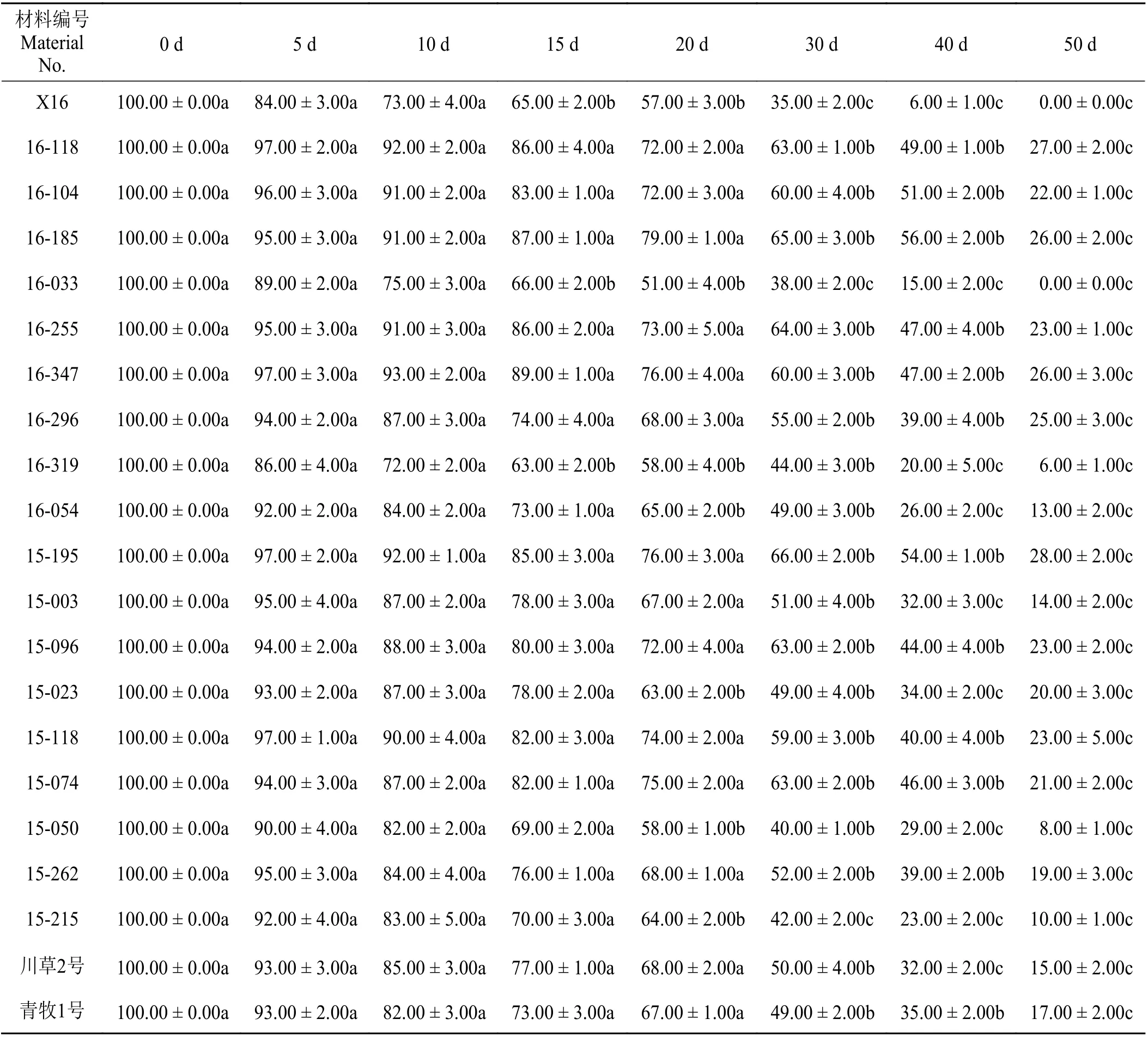
表5 干旱胁迫时间对老芒麦相对存活率的影响Table 5 Effectsof drought stresstime on relative survival percentageof Elymus sibiricus%
本研究中,供试的21份老芒麦材料在干旱处理下的株高均低于其对照组,且随着干旱胁迫天数的增加,老芒麦株高差距越发明显。表明干旱胁迫抑制了老芒麦株高的增长;同时也表明在干旱胁迫环境下,老芒麦为了汲取更多水分,通过降低株高,缩短根部水分到达叶部的距离,以促进水分的有效运输。这是老芒麦适应干旱环境的重要方式之一。
一般而言,在相同干旱胁迫下,对干旱胁迫敏感的植物,其相对生长速率较低,反之,则较高。本研究中,供试的21份老芒麦材料在干旱胁迫下的相对生长速率不尽相同。其中,门源县的16-118老芒麦和大通县的16-347、15-195、15-262老芒麦及参比材料中的川草2号老芒麦相对生长速率较大,而门源县的16-185老芒麦和大通县的15-096、15-215老芒麦相对生长速率较小,表明供试的老芒麦材料对干旱胁迫的敏感程度不同,即老芒麦材料间的耐旱性存在差异。这与刘小慧等[22]研究结果相类似。

表6 干旱胁迫时间对老芒麦生理指标的影响Table 6 Effects of drought stress time on physiological indexesof Elymus sibiricus

表7 干旱胁迫下21份老芒麦材料苗期耐旱隶属函数值及排名Table 7 Subordinate function value and ranking of 21 Elymus sibiricus materialsunder drought stressat the seedling stage
研究表明,随着干旱胁迫程度的增加,植物叶面积逐渐降低[23-24]。本研究中,干旱胁迫下,供试的21份老芒麦材料单叶面积均低于对照组,表明老芒麦叶面积增长受到了干旱胁迫的抑制。同时也说明在干旱胁迫环境下,老芒麦为防止水分过度蒸发,而表现为叶面积降低或叶片增长减缓。这与谢瑞娟等[25]研究结果基本一致。
张翠梅等[26]研究表明,随着干旱胁迫时间的延长,陇中苜蓿(Medicago sativa)根系干重不断增大。本研究中,干旱胁迫下的老芒麦材料根系干物质积累率高于对照组。这与王传旗等[27]研究结果相类似。说明在干旱胁迫环境下,为了汲取更多水分,老芒麦倾向于将更多的生物量分配到根部,即通过生物量投资的优化配置来适应干旱环境,这也是老芒麦种群适应干旱环境的重要途径之一。
相对存活率是反映植物抗逆性强弱的一个较为直观的生长量指标。相同的干旱胁迫环境下,植物相对存活率越高,其耐旱性越强。本研究中,老芒麦相对存活率与干旱胁迫天数呈负相关关系。干旱胁迫下,供试的21份老芒麦材料相对存活率均出现了不同程度的下降现象,且随着干旱胁迫天数的增加,老芒麦材料的相对存活率下降越发明显。这与李光莹等[28]研究结果类似。
老芒麦株高、叶片面积、根系干物质积累等形态的可塑性使得老芒麦具有更宽的生态幅和更强的耐旱性,这对于老芒麦物种的分布和老芒麦生物多样性的进化具有重要的意义。
3.2 干旱胁迫下苗期老芒麦生理指标变化特点
环境胁迫会影响植物叶绿素的合成与降解,因此可通过植物叶片叶绿素含量的变化来反映其对干旱胁迫的适应能力[29]。有研究表明,干旱胁迫条件下,牧草叶绿素含量降低,但抗旱性强的牧草叶绿素含量高于抗旱性弱的牧草叶绿素含量[30-31]。本研究也证实了这一点,供试的21份老芒麦材料叶绿素含量均随着干旱胁迫天数的增加而下降,且不同老芒麦材料的叶绿素含量下降幅度不同。其原因可能是干旱引起植物体内活性氧的积累,造成细胞膜损伤,从而加速了叶绿素的分解[32]。
相对电导率值是鉴定植物细胞膜稳定性的重要生理指标,同时也是反映植物受伤害程度的另一个侧面。本研究中,供试的21份老芒麦材料相对电导率值均随着干旱胁迫天数的增加而上升,但不同老芒麦材料的相对电导率值上升幅度不同且差距较大。表明干旱已对老芒麦叶片细胞膜造成伤害。这可能是由于干旱破坏了老芒麦体内抗氧化酶活性与活性氧类之间的动态平衡,造成细胞膜损伤,使膜中脂类物质大量外渗所导致。
3.3 苗期老芒麦的耐旱性鉴定
植物的耐旱性是一个复杂的综合性状,在特定地区的植物耐旱性往往是几个机制共同作用的结果[33]。因此,对于植物的耐旱性研究可以通过多种指标进行鉴定与评价。本研究中,鉴于老芒麦种质资源的丰富性和生态类型的复杂性,使用综合指标来反映老芒麦材料的耐旱性,并运用模糊数学中的隶属函数法对老芒麦耐旱性指标进行综合评价。这样可避免淘汰一些由于单一指标反映不出来的耐旱材料,或将不耐旱的材料误认为是耐旱的,最终使筛选出的耐旱性老芒麦材料较为可靠。
4 结论
本研究采用隶属函数法对供试的21份老芒麦材料苗期的耐旱性进行了综合评价。其中,大通县的15-195老芒麦属Ⅰ级强抗,16-347、15-003、15-096、15-023、15-118、15-074、15-050、15-262、15-215老芒麦材料属Ⅱ级较抗;互助县的16-296老芒麦属Ⅱ级较抗,16-319老芒麦属Ⅲ级弱抗;刚察县的X16老芒麦属Ⅱ级较抗;门源县的16-118、16-104、16-185、16-255、16-054老芒麦材料属Ⅱ级较抗,16-033老芒麦属Ⅲ级弱抗。参比材料中,青牧1号老芒麦和川草2号老芒麦同属于Ⅱ级较抗品种,但川草2号老芒麦耐旱性相对较强。供试的21份老芒麦材料中,大通县的15-195老芒麦在土壤水分亏缺条件下表现出了较强的耐旱能力,该野生草可作为老芒麦抗旱育种的重要参考草种。
本研究从生态生理角度仅对供试的21份老芒麦材料苗期的耐旱性进行了研究,不足以全面揭示其在青藏高原区域半干旱条件下的适应性。鉴于植物抗旱性的复杂性和提高旱区草牧业生产的紧迫性,建议今后应对老芒麦的耐旱性进行长期系统的综合研究,以期选育出高耐旱性的老芒麦新品种。
猜你喜欢
草业学报(2022年6期)2022-06-11
现代畜牧科技(2021年11期)2021-12-21
中国生态农业学报(中英文)(2021年8期)2021-07-28
三农资讯半月报(2020年3期)2020-03-09
校园足球(2019年5期)2019-09-10
中国畜禽种业(2019年7期)2019-01-06
种业导刊(2018年9期)2018-01-16
科学与财富(2017年17期)2017-06-16
湖北农业科学(2017年1期)2017-03-09
中国草食动物科学(2016年1期)2016-01-27
