
三明仙人谷不同海拔梯度下植物多样性和群落结构探究
2021-07-28 00:42冯岩
南方农业·下旬 2021年4期
冯岩

摘 要 以福建省三明市仙人谷城市森林生态系统为对象,研究不同海拔梯度下植物多样性及植物群落结构特征。结果表明,植物科分类水平上的数量和Shannon-Wiener多样性指数随海拔的增加而增加,而属、种、Simpson指数和Pielou均匀度指数都随海拔的增高而先增加后减少。海拔梯度和不同功能群水平都显著影响科属种的数量,且这2个因素对科属种的影响有交互作用,但海拔对科属种的3个多样性指数均无显著性影响。由此说明,城市森林在海拔梯度下的生物多样性保护功能表现出差异,三明市建设森林城市需要考虑人为活动、自然环境条件和本身生理生态特征对植物多样性的潜在影响,以期充分发挥可持续的森林生态服务功能。
关键词 海拔梯度;生物多样性;群落结构;森林公园
中图分类号:S718.54 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2021.12.001
生物多样性包含遗传多样性、物种多样性和生态系统多样性等[1]。生物多样性在维持整个生态系统平衡中起到了非常重要的作用[2],生物多样性的改变将直接影响整个生态系统的功能。随着经济全球化的发展、城市化的扩张及人口数量的持续增长,人类社会活动对生物多样性的影响愈来愈显著[3],如栖息地丧失与生境的破碎化[4]、资源的过度开发利用、环境污染和气候变化等,这些变化已经对物种的生存与发展造成了严重威胁[5-9]。
随着海拔梯度的变化,温湿度、光照强度和土壤养分发生改变,植物多样性也随之改变。例如,植物物种多样性随海拔的增加而呈现出先增后减的单峰模式[10],在对长江三峡库区海拔梯度下的研究也表现出相似的结果[11]。何艳华等对五鹿山植被进行研究,发现乔木层的α多样性指数都呈显著的单峰分布格局,草本层丰富度指数(Margalef)、香农-威纳指数(Shannon-Wiener)较低海拔下随海拔升高递减,随着海拔梯度升高,水热组合发生变化,地带性植被以及群落内的建群种和优势种均发生了变化[12]。此外,对长白山不同海拔梯度上牛皮杜鹃群落物种多样性进行了研究,认为海拔是影响牛皮杜鹃群落差异的主要因素,且海拔接近的群落间相似性系数较高[13]。
目前,对环境梯度下植物多样性动态的研究还不够全面,特别是城市森林在提供人类休闲娱乐生态功能的同时,对其多样性的保护功能报道较少。为此,以三明市仙人谷城市森林生态系统为对象,研究海拔梯度下植物多样性及植物群落结构特征,通过比较不同海拔下群落结构的差异性,以期为人类活动强烈背景下的城市森林植物保護及其功能的研究提供科学依据。
1 研究区域概况
试验地位于福建省三明市仙人谷国家森林公园
(E 117°38′35″,N 26°14′44″)。试验区地处沿海内陆山区,属中亚热带季风气候,兼备有大陆性和海洋性气候,表现为温暖湿润,四季分明,雨量充沛。夏季多东风炎热多雨,冬季多东北风寒冷干燥。区域内年平均雨量1 656.3 mm,3—9月是多雨季节,10月至翌年2月为少雨季节。年平均气温19.4 ℃,极端最高气温41.4 ℃,极端最低气温-5.8 ℃,日平均温度稳定通过≥10 ℃的年有效积温3 579.6 ℃,年日照时间1 637.4 h,年蒸发量
1 592.3 mm,无霜期305 d。仙人谷森林公园内主要为原始林和次生林,据调查园区有维管束植物130科、522种,森林覆盖率超95%。土壤有机碳含量平均为16.48 g·kg-1、全氮含量平均为1.73 g·kg-1,全磷含量平均为0.16 g·kg-1[14]。
2 研究方法
2.1 调查方法与试验设计
根据景区地形的情况,于2014年4月对样地进行了调查,在向阳面沿海拔400 m、500 m和600 m高程处每个海拔随机选取3块15 m×15 m的重复样地进行调查,每个样方内选择10 m×10 m亚样方进行乔木调查,
5 m×5 m的亚样方进行灌木调查,选择1 m×1 m的亚样方进行草本调查。调查并记录每个样方内高于5 cm的每种植物的名称、冠幅、株数和盖度。其中,样方内各物种按照生活型划分为一年生草本、多年生草本、一年生草质藤本、多年生草质藤本和小灌木5个生活型。
2.2 生物多样性计算
根据物种多样性的测度指数来分析植物多样性,以α多样性指数(Simpson指数和Shannon-Wiener指数)及均匀度指数(Pielou)来分析[15],公式如下:
Simpson指数=1-∑Pi2
Shannon-Wiener指数=-∑PilnPi
式中:Pi指种的个体数占群落中总个体数的比例。
Pielou指数=H/Hmax
式中:H指香农威纳指数;Hmax指最大香农维纳指数。
2.3 数据分析
采用Excel、Sigmaplot 12.0、SPSS 20.0等对数据进行了整理与分析。差异显著性检验设为P<0.05。
3 结果与分析
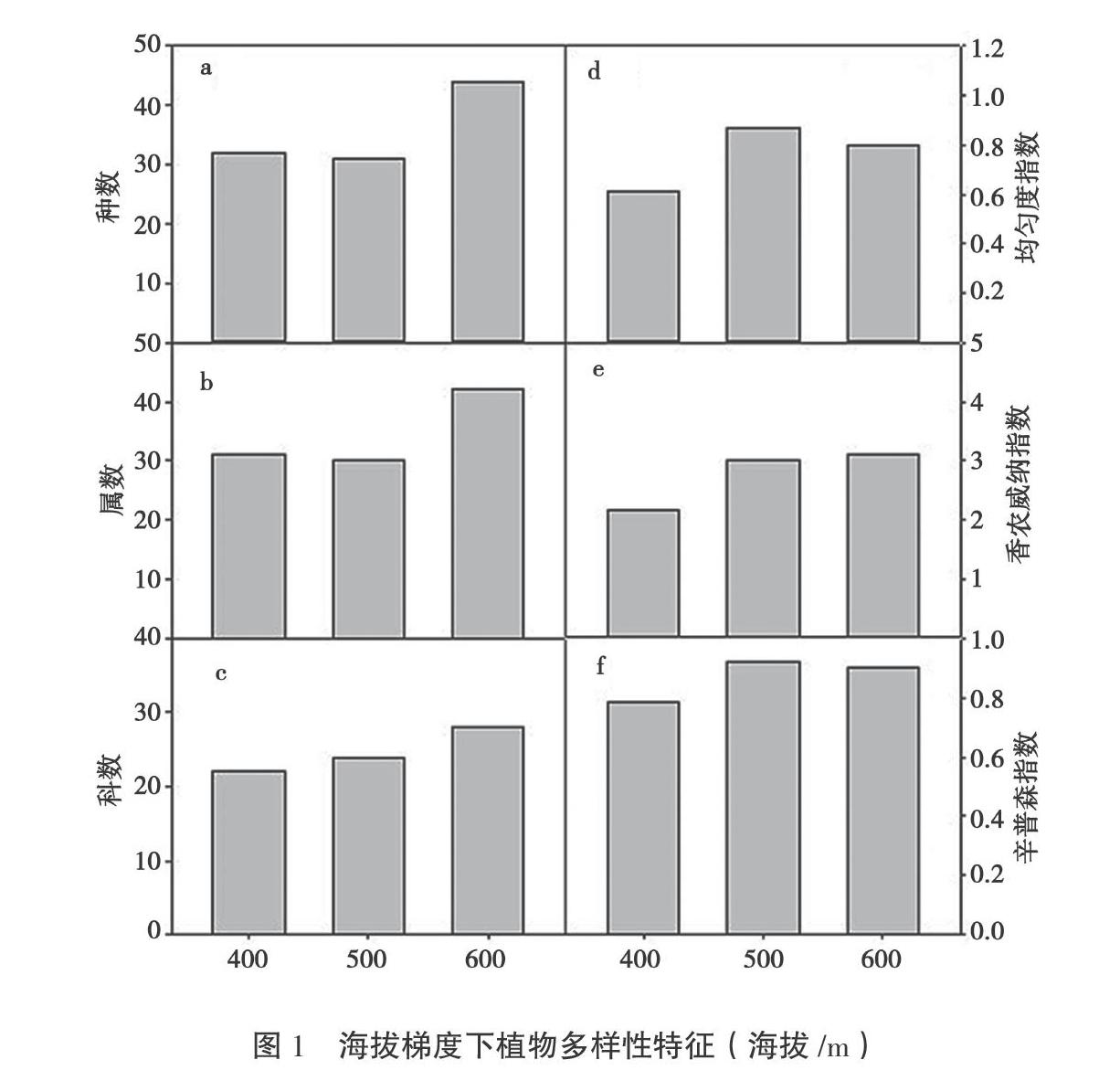
3个海拔梯度一共有80种植物,分布于46科73属。从图1可知,科的数量随海拔梯度的增加而增加,400 m和500 m处的种数,分别是32和31;600 m处较高,为
44(图1a);400 m、500 m、600 m处的属的数量为31、30、42(图1b);400 m、500 m、600 m处的科数分别为22、24、28(图1c),600 m处的科属种的数量皆为最高。
Pielou均匀度指数随样地海拔的增加而呈现出先增加后减少的趋势,分别为0.62、0.87和0.80(图1d)。Shannon-Wiener指数随海拔的升高而增加,分别为2.19、3.03和3.11(图1e)。α多样性的Simpson指数在海拔400 m为0.79,而500 m和600 m分别为0.92和0.91
(图1f)。
图2表明,海拔500 m处的乔木科属种的数量都低于400 m和600 m。灌木种和属的数量都随海拔梯度的上升而增加,科数500 m处最大,400 m低于600 m。草本的科属种的数量则是600 m>500 m≥400 m。乔木的Pielou指数随海拔梯度的增高而下降,乔木的Shannon-Wiener指数和Simpson指数500 m处的最低,400 m处最高。灌木和草本在500 m处的3个多样性指数皆高于400 m和600 m。
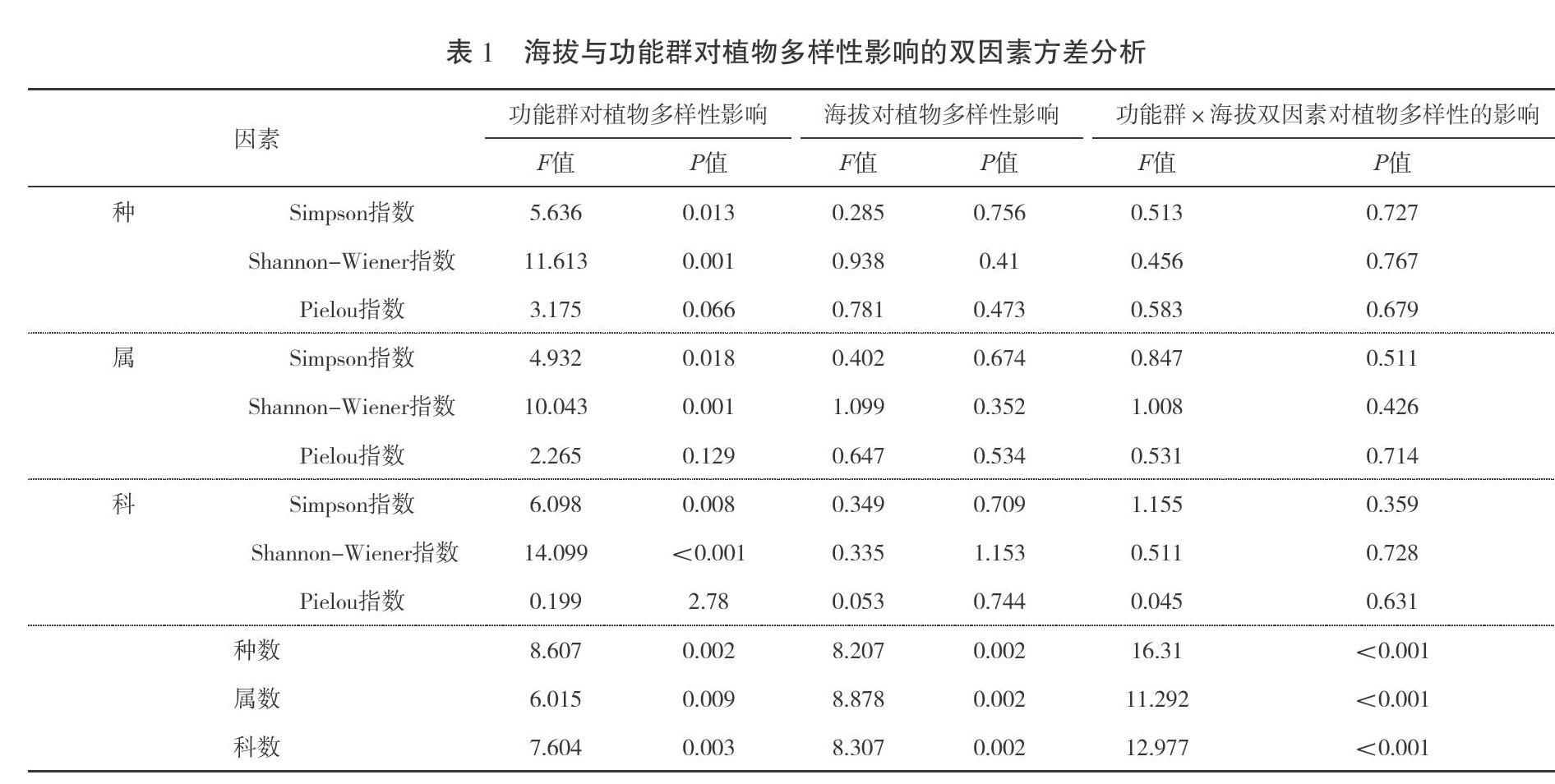
双因素统计结果见表1,功能群和海拔都显著的影响科属种的数量(P<0.01),功能群和海拔对科属种的影响有交互作用(P<0.01),对科属种的3个多样性指数的影响无交互作用。功能群对科属种的Simpson指数和Shannon-Wiener指数都有显著性影响(P<0.05),而对科属种的Pielou指数皆无显著性影响。海拔对科属种的3个多样性指数均无显著性影响。
根据表2可知,3个海拔梯度共有的植物共有21种,分布于18科。其中,山苍子(Litsea cubeba)、黄瑞木(Adinandra millettii)、虎皮楠(Daphniphyllum oldhami)、黄绒润楠(Machilus grijsii)、铁线蕨(Adiantum capillusveneris)及淡竹叶(Lophatherum gracile)六种植物在3个海拔均有发现。粗叶榕(Ficus hirta)、大葉白纸扇(Mussaenda esquiroill Levl.)、枇杷叶紫珠(Callicarpa kochiana)、二列叶柃(Eurya distichophylla)、杉木(Cunninghamia lanceolata)、南酸枣(Choerospondias axillaria)、马尾松(Pinus massoniana)及芒萁(Dicranopteris dichotoma)在400 m和600 m均有发现。狗骨柴(Tricalysia dubia)、华南紫萁(Osmunda vachellii)、乌饭树(Vaccinium bracteatum)在400 m和500 m处有发现。矩圆鼠刺(Itea oblonga)、山血丹(Ardisia punctata)、斜方复叶耳蕨(Arachniodes rhomboidea)、镰羽贯众(Cyrtomium balansae)在500 m和600 m处有发现。
此外,400 m处还有大芽南蛇藤(Celastrus gemmatus)、毛花连蕊茶(Camellia fraterna)、细枝柃(Eurya loquaiana)等15种特有植物;栲(Castanopsis fargesii)、赤楠(Syzygium buxifolium)、山矾(Syplocos sumuntia)等18种植物仅在500 m处出现;楤木(Aralia chinensis)、杜茎山(Maesa japonica)红腺悬钩子(Rubus stimulans)等26种植物仅在600 m处有发现(见表3)。
表2其中“√”代表该海拔存在此物种,表3中各植物学名为:大芽南蛇藤(Celastrus gemmatus)、毛花连蕊茶(Camellia fraterna)、细枝柃(Eurya loquaiana)、山乌桕(Sapium discolor)、尖萼红山茶(Camellia edithae)、水竹(Phyllostachys heteroclada)、木荷(Schima superba)、阴香(Cinnamomum burma-nnii)、石栎(Lithocarpus glaber)、杨梅(Myrica rubra)、半边旗(Pteris semipinnata)、小木通(Clematis armandii)、福建莲座蕨(Angiopt-eris fokiensis)、五节芒(Miscanthus floridulus)、金毛狗脊(Cibotium barometz)、栲(Castanopsis fargesii)、赤楠(Syzygium buxifolium)、山矾(Syplocos sumuntia)、秃瓣杜英(Elaeo-carpus decipiens)、裂斗锥(Castanopsis fissa)、瓜馥木(Fissistigma oldhamii)、桃叶石楠(Photinia prunifolia)、草珊瑚(Chloranthus glabra)、多花蓬莱葛(Gardneria multiflora)、刨花润楠(Machilus pauhoi)、狭叶山胡椒(Lindera angustifolia)、常山(Dichroa febrifuga)、厚叶红淡比(Cleyera pachyphylla)、大叶青冈(Cyclobalanopsis jenseniana)、花葶苔草(Carex scaposa)、木防己(Cocculus orbiculatus)、狗脊蕨(Woodwardia japonica)、刺葵(Phoenix hanceana)、钩藤(Uncaria rhynchophylla)、野木瓜(Holboellia coriacea)、栀子(Gardenia jasminoides)、多脉酸藤子(Embelia oblongifolia)、石栗(Aleurites moluccana)、楤木(Aralia chinensis)、杜茎山(Maesa japonica)、红腺悬钩子(Rubus stimulans)、三叶五加(Acanthopanax henryi)、枫香(Liquidambar formosana)、木莓(Rubus swinhoei)、灰毛泡(Rubus irenaeus)、南五味子(Kadsura longipedunculata)、朱砂根(Ardisia crenata)、福建山矾(Symplocos fukienensis)、拟赤杨(Alniphyllum fortunei)、格药柃(Eurya muricata)、络石(Trachelospermum jasminoides)、求米草(Oplismentls undulatifolius)、乌蕨(Stenoloma chusanum)、武夷山鳞毛蕨(Dryopteris wuyishanica)、土茯苓(Smilax corbularia)、海金沙(Lygodium japonicum)、鸡屎藤(Paederia scandens)、里白(Hicriopteris glauca)。
4 结论与建议
研究结果表明,科的种类及Shannon-Wiener多样性指数随海拔梯度的增加而增加,而属的种类、物种数、Simpson指数和Pielou均匀度指数都表现为在最低海拔400 m处调查样地最小。这种现象可能是因为海拔越高,人类活动对环境造成的影响就越低,同时随着海拔的增加,水热条件发生改变,捕食者的数量减少且种类也发生改变。再加上随海拔的增加,1~20 cm的土壤中的有机碳和总氮含量也发生了改变[14],这些因素共同作用于植物的生长繁殖,从而影响植物的多样性。
海拔的变化影响到不同功能群植物在科属种分类水平上的数量,且这海拔和功能群分类对科属种的影响有交互作用。海拔500 m处的乔木科属种的数量均低于400 m
和600 m。灌木种和属的数量都随海拔梯度的上升而增加,科数500 m处最大,400 m低于600 m。草本的科属种的数量则是600 m>500 m≥400 m。乔木的Pielou指数随海拔梯度的增高而下降,乔木的Shannon-Wiener指数和Simpson指数500 m处的最低,400 m处最高。灌木和草本在500 m处的3个多样性指数在皆高于400 m和600 m。功能群对科属种的Simpson指数和Shannon-Wiener指数都有显著性影响,而对科属种的Pielou指数皆无显著性影响,但海拔对科属种的3个多样性指数均无显著性影響,说明不同功能群的植物对环境的响应不同,从而影响到植物多样性和群落结构的差异[11-13]。
对于不同海拔出现的物种,研究认为在3个海拔都出现的为广布种,对于不同的海拔土壤和气候等特征都能够适应,而仅出现在一个海拔中的植物则称之为特有种,3个海拔梯度的特有种分别称之为低海拔特有种、中海拔特有种及高海拔特有种。研究发现,山苍子、黄瑞木、虎皮楠、黄绒润楠等六种植物为广布种,低海拔特有种大芽南蛇藤、毛花连蕊茶等15种植物,中海拔特有种有栲、赤楠、山矾等18种植物;高海拔特有种有楤木、杜茎山、红腺悬钩子等26种植物。其中低海拔特有种以乔木
居多。
研究发现,三明市仙人谷国家森林公园不同海拔影响植物多样性和群落结构,为在三明市打造城市绿道,增加森林城市生态福祉的同时,不仅要考虑人为活动对植物多样性的潜在影响,而且还要进一步研究造成植物分布差异的生境影响和植物本身生理生态机制,以期为生态文明建设提供科学依据。
参考文献:
[1] 钱迎倩.生物多样性研究的原理与方法[M].北京:中国科学技术出版社,1994:25.
[2] Loreau M,Naeem S,Inchausti P,et al.Biodiversity and ecosystem functioning:current knowledge and future challenges[J].Science,2001,294(5543):804.
[3] Liang Y Q,Li J W,Jing L.Impact of urbanization on plant diversity:A case study in built-up areas of Beijing[J].Forestry Studies in China,2008(03):179-188.
[4] 武晶,刘志民.生境破碎化对生物多样性的影响研究综述[J].生态学杂志,2014(07):1946-1952.
[5] 魏辅文,聂永刚,苗海霞,等.生物多样性丧失机制研究进展[J].科学通报,2014(06):430-437.
[6] 马克平,钱迎倩.生物多样性保护及其研究进展(综述)[J].应用与环境生物学报,1998,4(yy):95-99.
[7] Sala O E,Wall D H.Global biodiversity scenarios for the year 2100[J].Science,2000,287(5459):1770-1774.
[8] Chen X,Li B L.Tree diversity change in remaining primary mixed-broadleaved Korean pine forest under climate change and human activities[J].Biodiversity & Conservation,2004,13(03):563-577.
[9] 吴则焰,林文雄,陈志芳,等.中亚热带森林土壤微生物群落多样性随海拔梯度的变化[J].植物生态学报,2013(05):397-406.
[10] Changbae L,Junghwa C,Hyunje C.Elevational patterns and determinants of plant diversity in the Baekdudaegan Mountains,South Korea:Species vs.functional diversity[J].Chinese Science Bulletin,2013(31):3748-3770.
[11] 李巧燕,王襄平.长江三峡库区物种多样性的垂直分布格局:气候、几何限制、面积及地形异质性的影响[J].生物多样性,2013(02):141-152.
[12] 何艳华,闫明,张钦弟,等.五鹿山国家级自然保护区物种多样性海拔格局[J].生态学报,2013(08):2452-2462.
[13] 金慧,赵莹,赵伟,等.长白山牛皮杜鹃群落物种多样性的海拔梯度变化及相似性[J].生态学报,2015(01):125-133.
[14] 李林源.福建三明仙人谷不同海拔梯度下土壤碳氮磷含量特征研究[J].南昌工程学院学报,2014
(04):28-31.
[15] 马克平,刘玉明.生物群落多样性的测度方法 Ⅰα多样性的测度方法[J].生物多样性,1994(04):231-239.
(责任编辑:刘 昀)
猜你喜欢
故事作文·低年级(2019年8期)2019-08-20
好孩子画报(2018年12期)2018-02-13
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
环球人文地理·评论版(2016年5期)2017-01-03
农业与技术(2016年15期)2016-11-09
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
儿童时代·幸福宝宝(2009年7期)2009-11-23
