
胡杨叶片的胞间CO 2浓度及气孔和非气孔限制的探究
2021-08-04 03:56武胜利管文轲史智欣何宇翔岳永江
湖北农业科学 2021年13期
林 兵,武胜利,管文轲,宋 川,史智欣,何宇翔,,岳永江,
(1.新疆师范大学地理科学与旅游学院,乌鲁木齐 830054;2.新疆林业科学院,乌鲁木齐 830018)
植物在水分胁迫下,光合作用持续减弱的主要原因为两方面[1-3]:一是气孔导度Gs的减小,导致植物气孔内的CO2减少,并致使植物无法维持高强度的光合作用,这个过程被称为植物光合作用的气孔限制,以气孔限制值Ls来表示;此外,因为植物叶片温度的不断升高,导致出现叶绿体活性以及Rubisoo活性降低、RuBP羧化酶再生能力降低等情况[4],并最终导致叶片的光合作用能力减弱,这个过程称之为光合作用的非气孔限制。胞间CO2作为植物进行光合作用的中介[5],它会受到大气CO2浓度的制约,同时还受到植物叶片气孔导度和叶片光合消耗的影响和制约,因此对植物叶片胞间CO2浓度变化规律的分析是探究植物气孔限制和非气孔限制的基础。
荒漠河岸胡杨(PopuluseuphraticaOliv.)属杨柳科胡杨亚属木本植物,是塔里木河荒漠河岸林中最常见和典型的耐干旱、耐盐碱的重要林木树种之一[5,6]。在干旱的荒漠地区,胡杨有着极强的抗逆能力,其对于维持荒漠地区的生态环境起着关键的作用[7]。目前国内外对胡杨进行了一些研究和报道,张霞等[8]、史军辉等[9]分析了胡杨的耐盐性;武逢平等[10]、周燕等[11]研究了胡杨的根蘖繁殖特性和胡杨雌花离体培养中的不定根诱导和植株再生体系的建立;吐热尼古丽·阿木提等[12]对胡杨生态系统的脆弱性进行了研究;黄文娟等[13]分析了胡杨异形叶结构型性状及其与胸径的关系;吴桂林等[14]、王海珍等[15]、王勇等[16]研究了胡杨的水分利用特性;王希义等[17]研究了胡杨的生物量和种群生存特征;钟小莉等[18]、王海珍等[19]研究了干旱胁迫下氮素对胡杨幼苗生长及光合的影响和胡杨异形叶光合系统Ⅱ叶绿素荧光特性;高生峰等[20]对极端干旱区淹灌对胡杨长势的影响进行了研究。但截至目前,对胡杨叶片的胞间CO2浓度,以及气孔限制与非气孔限制因子的研究不多。本文在分析胞间CO2浓度的季节变化及其影响因素基础上,探究了胡杨叶片的气孔限制情况,以及非气孔限制因子的变动周期规律,研究结果有助于进一步探索胡杨在水分亏缺条件下叶片的气孔调节规律,为未来对塔里木河下游胡杨林的保护、恢复以及更新提供理论依据。
1 研究区概况
尉犁县地处新疆维吾尔自治区的中部区域,为巴音郭楞蒙古自治州所管辖。尉犁县系暖温带大陆性干旱荒漠气候,温度冷热差异大,年月变化差距大,冬天干旱且寒冷,夏天酷热。整年热量丰富,空气干燥,蒸发量大,降水稀少,年际变化差距大,光照充足。全年平均气温10.5℃,最冷月(1月)平均气温-11.2℃,极端最低气温-22.6℃;最热月(7月)平均气温25.6℃,极端最高气温38.3℃;气温年较差为36.8℃。全年平均降水量仅有50.7 mm,但是年平均潜在蒸发量却多达2 730.3 mm。尉犁县的主要风向为北偏东,以东北风为主要风向,其次风向为南偏西,年均风速2.3 m/s,最大风速可达24 m/s(10级)。每年春夏季节8级以上大风年均为15 d,风沙日数23.1 d,浮尘天数24.2 d。境内主要天然植物有胡杨、灰胡杨(PopuluspruinosaSchrenk.)、柽柳(Tamarixspp.)、胀果甘草(Glycyrrhiza inflateBat.)和罗布麻(Apocynum venetumLinn.)等。
2 材料与方法
2.1 研究材料
试验于2018年6—8月在塔里木河中下游尉犁县境内的天然胡杨林进行,选取具有代表性的3株长势健康、树干通直、树皮光滑、无病虫害的胡杨树木作为待测样株。同时对3株胡杨标记编号,树样的基本生物学参数见表1。
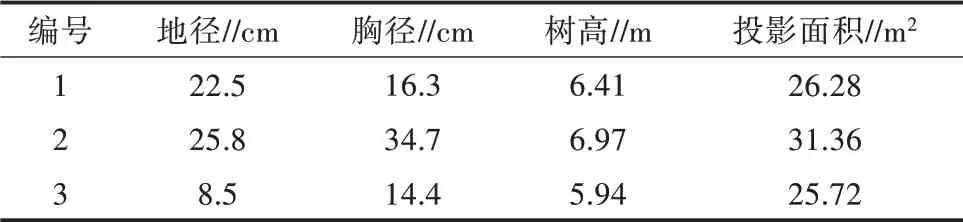
表1 胡杨试验树样的生物学参数
2.2 测定项目和方法
光合生理参数的测定:2018年6—8月在胡杨速生期于尉犁县塔里木河中下游展开,选取晴朗无云日进行,每月1次,每次3 d,取外围向阳新树梢上倒3、4位成熟叶片,于6:00—22:00每2 h测定1次,每次选取3~4片功能叶作为重复。
测定方法:使用LI-6400便携式光合作用测定系统,测定胡杨叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等指标及有效光合辐射(PAR)、大气CO2浓度(Ca)、气温(Tair)、叶温(Tleaf)、空气相对湿度(RH)、叶片内外水汽压差(VPDL)等微气象参数。
2.3 气孔限制值的计算
国内外学者从不同的角度提出了多种计算方法。目前最常用的计算气孔限制值(Ls)的方法如下公式[21]:

式中,Ca为大气CO2浓度,单位mmol/mol;Ci为胞间CO2浓度,单位mmol/mol。
2.4 数据分析
通过Excel软件数据进行筛选。使用SPSS软件进行数据的整理、统计以及分析。
3 结果与分析
3.1 胞间CO 2浓度和影响因子变动规律
在胡杨的生长季6—8月,选择晴朗无云的晴天作为实验数据测量日,以分析胡杨叶片胞间CO2浓度的日变化规律和影响因素的同步变化规律。从胞间CO2浓度Ci的变化(图1a)可以看出,在生长季内胡杨叶片的胞间CO2浓度均呈“V”字型的变化趋势,最低值均出现在14:00,这与徐俊增等[22]、翁晓燕等[23]的研究结果基本一致。同时,生长季内变化趋势幅度为6月>7月>8月,各月差异十分明显。

图1 速生期胡杨胞间CO2浓度(a)、大气CO2浓度(b)、净光合速率(c)、气孔导度(d)和蒸腾速率(e)的日变化
胡杨叶片胞间CO2浓度变动周期规律受Gs、Ca、Pn和Tr等因素的共同影响和制约。如图1,表明叶片Ci变动周期规律主要受Ca的影响。当Ca出现“双低”值时,胞间CO2浓度则也会紧跟其后,并由此发生相应的变化。6月胞间CO2浓度在14:00和16:00和8月出现低值就是受大气CO2浓度的直接影响。
由于Gs、Pn和Tr随外部光强的变化而随即产生相应变化,气孔缓慢打开的上午与缓慢合上的下午呈现了显著差异的规律。上午,气孔呈现出缓慢打开的趋势,气孔导度、蒸腾速率和净光合速率逐步递增。但同时胞间CO2浓度表现为逐步递减,其原因一是大气CO2浓度正在逐步递减,另一方面是叶片的净光合速率和蒸腾速率的升高进而致使胡杨叶片胞间CO2浓度的消耗大大加剧,Ca随之逐步递减,当Pn处于峰值时,Ca则降至最低值。午后再随着太阳辐射的减弱,最终致使胡杨叶片的净光合速率、蒸腾速率急剧减小,叶片的光合消耗减弱,同时大气CO2浓度则逐步升高,胞间CO2浓度因此呈现增加的趋势。
由此可知,胡杨叶片胞间CO2浓度和大气CO2浓度的律动周期大体一致,与净光合速率、蒸腾速率则相反。这与胞间CO2既是作为中介受到大气CO2的补给,同时也受叶片光合及蒸腾消耗的影响的结论一致,与徐俊增等[22]、郭文霞等[24]的研究相符。而胡杨叶片Ca与Gs的律动规律为相反趋势,呈负相关,原因为高气孔导度相应会产生更高的净光合速率和蒸腾速率。因此在这样高气孔导度、高净光合速率和高蒸腾速率的时段,叶片光合和蒸腾的消耗远多于经过气孔进入胡杨叶片胞间的CO2。
3.2 胡杨胞间CO2浓度与光合速率的关系
从生长季各个测量日10:00—12:00(图2a)和14:00—16:00(图2b)的胞间CO2浓度与净光合速率的规律来看,在不同时段,净光合速率与胞间CO2浓度的联系呈现不同。10:00—12:00时,净光合效率与胞间CO2浓度表现为正线性关系,且不显著。而14:00—16:00由于净光合速率的升高,胞间CO2浓度随即表现出一定的下降,和徐俊增等[22]、郭文霞等[24]的研究相符。这也说明光合作用对细胞CO2的耗损与气孔对CO2的导度联合决定了胞间的CO2浓度。14:00—16:00由于净光合速率升高而胞间CO2浓度减小,说明这一时段CO2进入胞间的速率明显小于强烈光合作用对CO2的耗损,因此显现出部分的气孔限制。
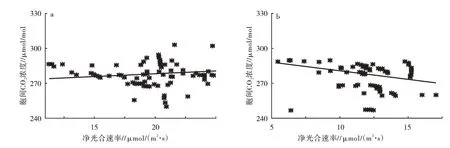
图2 胡杨10:00—12:00(a)和14:00—16:00(b)叶片胞间CO2浓度与净光合速率的关系
4 气孔限制与非气孔限制的讨论与探究
植物叶片的光合作用是受到气孔和非气孔限制因子的联合影响和制约[25-27]。
4.1 气孔限制值
气孔限制值Ls是解释和反映因为气孔导度的减小进而致使的对净光合速率的制约大小[28-30]。在生长季内,分析胡杨叶片Ls的变动周期规律,可知Ls总体上为先升高后减小的趋势,具体表现为早上和晚上低、中午高。但各月具体变化规律有一定的差异。6月从6:00开始一直上升,10:00—12:00期间增速减缓,然后一直增加至16:00。造成这种结果,主要是因为大气CO2浓度的直接影响,使得气孔限制值一直居高不下。7—8月的变化趋势基本相同,先升高随后轻微减小,而后上升最后快速下降。气孔限制值Ls主要是随净光合速率的升高而增大。这是因为气孔限制值是一个相对概念,主要是由气孔对CO2导度和光合作用的需求来联合决定的。在早上和晚上,叶片的气孔导度较低,但此刻光合作用所要耗损的CO2也低,故在这两个时段表现为较低的气孔限制状况。另一方面,胡杨叶片在中午的气孔导度较高,但光合作用所耗损的CO2更多,因此表现出较高的气孔限制(图3)。

图3 胡杨气孔限制值Ls的日变化
4.2 非气孔限制指标
非气孔限制是反映非气孔限制因子的重要指标,非气孔限制因子包括叶温过高导致叶绿体活性与Rubisoo活性降低、RuBP羧化酶再生能力降低等[31]。许大全[1]在相关研究中提到:断定叶片光合作用减弱是受气孔限制还是非气孔限制的可靠判据是Ci和Ls的变化规律方向,即Ci降低和Ls升高表明受气孔导度降低的影响;而Ci增大和Ls降低则表明是受非气孔因子的限制和影响。
从图4可以看出,胡杨生长季内各月下午时段的气孔限制值Ls呈下降趋势,而胞间CO2浓度则呈上升趋势,说明此时间段主要为非气孔限制因素限制了胡杨的光合作用。同时,7月和8月10:00—14:00和12:00—16:00,净光合速率开始下降,出现了以非气孔限制因素为主造成了光合速率下降,随后出现了气孔限制因素造成光合速率下降。这表明在一个时段造成胡杨净光合速率的降低是以非气孔限制或气孔限制因素为主,但不能排除气孔限制或非气孔限制因素的共同作用。

图4 胡杨的非气孔限制指标
而在Ramanjulu等[32,33]的研究中,则用胞间CO2浓度与气孔导度Gs的比值Ci/Gs来表示非气孔因子的制约状态。从生长季内各月胡杨叶片Ci/Gs的变动制值变化方向所分析出来的结果相一致。周期规律看出:Ci/Gs与气孔限制值Ls的变动周期规律相反,早上和晚上高、中午低。其因为早上和晚上的太阳辐射不高,而与光合作用有密切关系的酶的活性不能充分发挥,呈现出较强烈的非气孔限制影响。在生长季内的各月,10:00—18:00胡杨叶片的非气孔限制变化皆表现出比较平缓的变动规律。7月和8月胡杨净光合速率下降的10:00—14:00和12:00—16:00,Ci/Gs值均呈现上升趋势并出现了一个高峰,这也与前文中以胞间CO2浓度和气孔限
5 小结
本文分析了生长季内胡杨叶片胞间CO2浓度的变化规律及其影响因素,并分析探究了胡杨气孔限制和非气孔限制的情况,结果显示,生长季内胡杨叶片的胞间CO2浓度变化呈“V”型的变化趋势,最低值均出现在14:00。胡杨胞间CO2浓度随影响因素包括大气CO2浓度、气孔导度、净光合速率和蒸腾速率的变化而同步变化:与大气CO2浓度的变化规律基本保持一致;与气孔导度、净光合速率和蒸腾速率的变化规律相反,呈负相关。联合使用气孔限制值和非气孔限制这两个判断指标,能够进一步了解导致胡杨的净光合速率下降的气孔限制和非气孔限制状况和原因。但仍然存在一些不足,未能全面参考植物的其他生理生态数据,例如植物的水分利用效率、叶水势等指标。关于这点,今后可进行更深入的研究。
猜你喜欢
生态学报(2024年1期)2024-01-25
东北林业大学学报(2022年4期)2022-05-16
新疆农业科技(2021年2期)2021-07-03
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
生物技术通报(2014年10期)2014-03-21
赤峰学院学报·自然科学版(2012年19期)2012-10-14
