
鸭坦布苏病毒甲基转移酶的真核表达及鉴定
2021-09-12 16:34韩凯凯严若峰刘青涛刘宇卓李银赵冬敏黄欣梅杨婧
江苏农业科学 2021年15期
韩凯凯 严若峰 刘青涛 刘宇卓 李银 赵冬敏 黄欣梅 杨婧


摘要:甲基转移酶(methyltransferase,MTase)是鸭坦布苏病毒(duck Tembusu virus,DTMUV)非结构蛋白5的重要功能基团,在病毒复制及致病过程中发挥重要作用。为了对鸭坦布苏病毒甲基转移酶进行真核表达,并对其生物学功能进行鉴定,以GenBank中收录的DTMUV JS804株基因序列为模板,设计针对MTase的特异性引物,提取TMUV RNA,反转录得到cDNA,以其为模板进行扩增,得到MTase的基因片段,然后亚克隆至pCMV-Flag真核表达载体中,构建重组质粒pCMV-Flag-MTase,提取去除内毒素的重组质粒,并转染至BHK-21细胞,通过间接免疫荧光观察和Western Blot鉴定该蛋白的表达。结果显示,质粒pCMV-Flag-MTase经双酶切鉴定证明构建正确,通过间接免疫荧光与Western Blot检测,重组质粒在BHK-21细胞内正常表达,表达的蛋白与天然坦布苏病毒MTase具有相同的反应原性。
关键词:鸭坦布苏病毒;甲基转移酶;真核表达
中图分类号:S852.65+7 文献标志码: A
文章编号:1002-1302(2021)15-0147-03
收稿日期:2020-12-16
基金项目:国家自然科学基金(编号:31502101);江苏省农业科技自主创新资金[编号:CX(20)3093]。
作者简介:韩凯凯(1983—),男,河南新乡人,博士,副研究员,主要从事水禽疫病防控相关研究。E-mal:hankk0917@126.com。
坦布苏病毒属于黄病毒科黄病毒属蚊媒病毒类的恩塔亚病毒群,是一种具有包膜的单股正链RNA病毒,通过节肢动物的叮咬进行传播而导致宿主发病,主要引起20日龄以内的雏鸭出现严重的神经症状,死淘率增高及产蛋鸭的产蛋大幅下降[1]。坦布苏病毒基因组是单股正链RNA,含有一个大型开放阅读框,可编码7种非结构蛋白(NS1、NS2A、NS2B、NS3、NS4A、NS4B、NS5)和3种结构蛋白(C、PM和E)[2]。NS5在这几种蛋白中分子量是最大的,大约100 ku,整个NS5蛋白具有2个功能域,分别是N端的甲基转移酶结构域和C端的RNA依赖的聚合酶结构域。NS5蛋白甲基转移酶功能基团同时具有N-7和2′-O这2种甲基化功能,可以催化病毒遗传物质RNA的5′端形成一型的RNA帽子[3],这一帽子结构对病毒具有重要作用,可以防止病毒遗传物质被宿主细胞内的核酸外切酶降解,辅助病毒完成免疫逃逸。
目前,在黄病毒属其他病毒,如登革病毒、西尼罗河病毒、黄热病毒、寨卡病毒上均有甲基转移酶的相关报导[4-5],但在鸭坦布苏病毒上却鲜有研究。本研究构建了鸭坦布苏病毒甲基转移酶基因的真核表达系统,并在哺乳动物细胞中高效表达MTase重组蛋白,旨在為进一步研究其功能、研发坦布苏病毒病的诊断、治疗方法奠定基础。
1 材料与方法
1.1 病毒、细胞和载体
坦布苏病毒分离株(JS804)、BHK-21细胞株、真核表达载体pCMV-Flag,均由笔者所在实验室保存。E.coli DH5α感受态细胞,购于TaKaRa公司。
1.2 主要试剂
限制性内切酶、Ex Taq DNA聚合酶、pMD19-T载体、T4 DNA连接酶、蛋白质相对分子质量标准等,购自宝生物工程(大连)有限公司;体液病毒RNA提取试剂盒、质粒小量提取试剂盒、PCR产物纯化试剂盒,购自Axygen生物科技有限公司;脂质体转染试剂lipofectamine 3000,购自Invitrogen公司;FITC标记的山羊抗小鼠IgG,购于碧云天公司;辣根过氧化物酶标记的山羊抗小鼠IgG,购于中杉金桥公司;DMEM细胞培养基、胎牛血清,购自Gibco公司。
1.3 引物设计与合成
根据GenBank中登录的坦布苏病毒JS804株(GenBank登录号JF895923) NS5基因序列,设计PCR 扩增引物,其中上游引物为MTase-F:5′-CAGGGACCCTCGAGATGGGAGGGGGAACT-3′,划线部分为引入的XhoⅠ酶切位点。下游引物为MTase-R:5′-CCAAGTCTTCCGCGGATTGTCTTGGTCAT-3′,划线部分为引入的SacⅡ酶切位点。以上引物由南京金斯瑞生物有限公司合成,预期扩增片段大小为900 bp。
1.4 DTMUV MTase基因的扩增
按照Axygen公司体液病毒RNA提取试剂盒要求提取DTMUV JS804株的总RNA,并按照反转录试剂盒操作说明书,合成反转录产物cDNA。以cDNA为模板,MTase-F和MTase-R为引物进行PCR扩增。扩增体系(25.0 μL体系):其中双蒸水15.0 μL,10×Ex PCR Buffer 2.5 μL,25 mmol/L MgCl2 1.5 μL,2.5 mmol/L dNTP 2.0 μL,上、下游引物(25.0 pmol/μL)各1.0 μL,cDNA产物1.5 μL,Ex Taq (5 U/μL) 0.5 μL。PCR反应程序:95 ℃预变性3 min;95 ℃变性15 s,56 ℃退火15 s,72 ℃延伸60 s,30 次循环;72 ℃总延伸5 min。PCR产物经1%琼脂糖凝胶电泳鉴定,鉴定正确后按照琼脂糖DNA凝胶回收试剂盒操作说明进行产物回收纯化。回收的片段按照pMD19-T载体说明书进行连接,构建重组克隆质粒,并转化至DH5α感受态细胞中,经LB固体培养基37 ℃倒置过夜培养后,挑取单菌落振荡培养3 h,并对菌液进行PCR鉴定。鉴定正确的菌液取500 μL送至南京生工生物科技有限公司进一步测序,鉴定正确后按照质粒提取试剂盒操作说明提取质粒。
1.5 重组质粒pCMV-Flag-MTase的构建
用XhoⅠ、SacⅡ对重组克隆质粒及pCMV-Flag载体双酶切,电泳切胶回收,将2种酶切后回收的片段按照适宜比例与T4 DNA 连接酶16 ℃连接过夜。连接产物转化至DH5α 感受态细胞中,涂布于卡那霉素的平板,37 ℃倒置过夜培养,挑取单菌落进行菌液PCR鉴定,鉴定正确的菌液扩大培养后提取质粒,用XhoⅠ、SacⅡ进行双酶切鉴定,酶切鉴定正确的克隆送至南京擎科生物科技有限公司测序,鉴定正确的重组质粒命名为pCMV-Flag-MTase。
1.6 BHK-21细胞的转染
按照质粒小量提取试剂盒要求提取pCMV-Flag-MTase,分光光度计测定其浓度。取处于对数生长期的BHK-21细胞,接种24孔细胞培养板,细胞铺至细胞板约90%时进行转染。按照lipofectamine 3000转染试剂操作说明书制备DNA-脂质体混合物,其中转染的DNA用量为2.5 μg。混匀后室温孵育20 min,将DNA-脂质体混合物均匀铺到细胞中。
1.7 间接免疫熒光鉴定
取质粒pCMV-Flag-MTase转染24 h后的BHK-21细胞,PBS洗涤3次,采用4%多聚甲醛固定细胞10 min;PBS洗涤3次,每次5 min,采用0.5% triton X-100对细胞透化处理10 min;PBS洗涤3次,5 min/次,加入1% BSA,室温封闭30 min;PBS洗涤3次,5 min/次,然后加入事先制备的坦布苏病毒阳性小鼠血清(1 ∶1 000),室温孵育1 h,PBS洗涤3次,5 min/次,最后加入FITC标记的山羊抗小鼠IgG(1 ∶400),室温孵育30 min,PBS洗涤3次,5 min/次,倒置荧光显微镜下观察,照相记录。
1.8 Western Blot鉴定
pCMV-Flag及pCMV-Flag-MTase分别转染BHK-21细胞,48 h后收集细胞,提取细胞总蛋白,测定蛋白浓度后经Western Blot检测该蛋白的表达情况。先进行SDS-PAGE电泳,再经转膜、封闭,然后以抗坦布苏病毒阳性小鼠血清体为一抗,辣根过氧化物酶标记的山羊抗小鼠IgG为二抗进行反应,ECL显影观察结果。
2 结果与分析
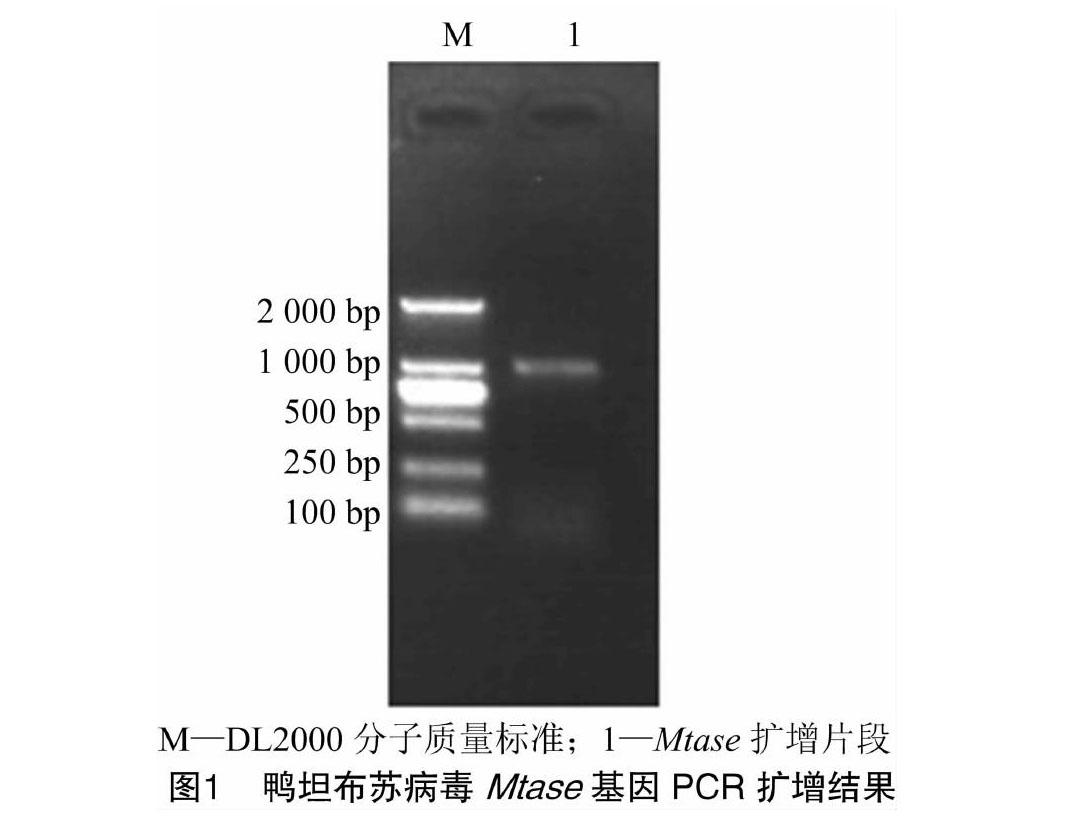
2.1 鸭坦布苏病毒MTase基因的PCR扩增
由图1可知,以含有鸭坦布苏病毒JS804株全长基因的cDNA为模板,采用所设计的特异性引物扩增MTase基因全长,结果扩增的片段大小约为900 bp,与预期片段大小相符。
2.2 重组质粒pCMV-Flag-MTase的PCR和双酶切鉴定
由图2可知,菌液PCR结果显示,挑取的单克隆均为阳性,可扩增得到与MTase大小一致的DNA片段。利用XhoⅠ、SacⅡ对重组质粒pCMV-Flag-MTase进行双酶切,结果显示出现2条特异性的条带,一条大小约为4 300 bp,为pCMV-Flag载体条带,另一条大小为900 bp,与MTase大小一致。测序结果显示,MTase基因序列与GenBank发布的序列同源性达99%。结果表明真核质粒pCMV-Flag-MTase构建成功。
2.3 重组质粒pCMV-Flag-MTase的转染效果
采用脂质体转染试剂lipofectamine 3000介导pCMV-Flag-MTase转染至BHK-21细胞,24 h固定间接免疫荧光检测表达情况。通过倒置荧光显微镜观察,由图3可知,正常未转染细胞无绿色荧光,而重组质粒转染细胞能观测到绿色荧光,荧光多数位于细胞质中,表明重组质粒pCMV-Flag-MTase能成功转染至BHK-21细胞中,其转染效率未受外源插入蛋白的影响。
2.4 DTMUV MTase在BHK-21细胞中的表达
由图4可知,经Western Blot检验,重组DTMUV MTase蛋白与坦布苏病毒阳性小鼠血清能发生特异性结合,条带分子量大小约30 ku,表明重组DTMUV MTase蛋白具有免疫反应活性。
3 讨论
鸭坦布苏病毒作为黄病毒科、黄病毒属、恩塔亚病毒群的重要成员,自2010年4月来,该病迅速传播到我国绝大部分养鸭地区,不仅给养鸭业带来了很大经济损失,而且极大地影响和制约了我国养鸭业乃至畜牧业的稳定健康发展[2]。因此,除了快速研制针对性的疫苗外,深入探索坦布苏病毒致病性、病毒蛋白质的结构功能及快速简便的临床检测方法,也成为现阶段防控坦布苏病毒病发生的重要技术手段。
NS5蛋白是坦布苏病毒基因组编码最大的蛋白质,蛋白质分子量可达100 ku,是黄病毒中最为保守的蛋白。而坦布苏病毒NS5蛋白的N端前900 bp为甲基转移酶活性的主要活性区,同时具有N-7和2′-O这2种甲基转移酶活性且对RNA序列有依赖性,参与病毒基因组RNA复制。有研究发现,当MTase的甲基化位点发生突变时,其病毒复制的能力将显著降低[6]。同时,MTase蛋白对宿主抗病毒反应也有拮抗作用,西尼罗病毒MTase蛋白的 2′-O甲基化功能区可以下调一种干扰素诱导产生蛋白的表达,进而使病毒逃避宿主的天然免疫反应,具体作用机制和生物学反应过程迄今尚不明确[7]。鉴于此,MTase作为DTMUV的主要蛋白酶,与其他非结构蛋白共同作为病毒基因组复制和转录的承载体,负责蛋白翻译后的切割、修饰等重要过程,在DTMUV病毒粒子成熟和复制过程中承担重要角色。又由于MTase在黄病毒属病毒中具有高度保守的氨基酸序列[7],因此可以作为研究抗坦布苏病毒相关药物和疫苗的关键靶标。目前,关于坦布苏病毒MTase蛋白相关研究较少,本研究通过克隆DTMUV MTase蛋白编码基因,并成功克隆至pCMV-Flag真核表达载体,通过间接免疫荧光和Western Blot试验检测,发现MTase蛋白表达成功。本研究获得的重组坦布苏病毒MTase蛋白可作为包被抗原,建立ELISA方法来检测血清中的抗体水平,亦可以用来筛选具备诊断功能/中和活性的单克隆抗体,此外,还为研究坦布苏病毒的感染致病机制提供了基础。
参考文献:
[1]吴 双,姜 勇,徐建生,等. 鸭坦布苏病毒、鸭肠炎病毒和番鸭细小病毒TaqMan三重实时荧光定量PCR检测方法的建立与临床应用[J]. 江苏农业学报,2020,36(3):626-633.
[2]杨志远,段会娟,王小蕾,等. 4株鸭坦布苏病毒的毒力、E基因序列和抗原差异性[J]. 中国农业科学,2019,52(23):4406-4414.
[3]Egloff M P,Benarroch D,Selisko B,et al. An RNA cap (nucleoside-2′-O-)-methyltransferase in the flavivirus RNA polymerase NS5:crystal structure and functional characterization[J]. The EMBO Journal,2002,21(11):2757-2768.
[4]Geiss B J,Thompson A A,Andrews A J,et al. Analysis of flavivirus NS5 methyltransferase cap binding[J]. Journal of Molecular Biology,2009,385(5):1643-1654.
[5]Zhou H,Wang F,Wang H,et al. The conformational changes of Zika virus methyltransferase upon converting SAM to SAH[J]. Oncotarget,2017,8(9):14830-14834.
[6]周希珍,赵 慧,高 岚,等. 虫媒黄病毒NS5蛋白的生物学功能研究进展[J]. 军事医学,2012,36(1):70-72.
[7]Dong H,Zhang B,Shi P Y. Flavivirus methyltransferase:a novel antiviral target[J]. Antiviral Research,2008,80(1):1-10.
