
赣江发现溯河洄游型刀鲚(Coilia nasus)*
2021-09-23 11:45杨一帆高小平轩中亚陈修报李乐康刘洪波
湖泊科学 2021年5期
杨一帆,姜 涛,高小平,轩中亚,陈修报,李乐康,刘洪波,杨 健,
(1:南京农业大学无锡渔业学院,无锡 214081) (2:中国水产科学研究院长江中下游渔业生态环境评价与资源养护重点实验室,中国水产科学研究院淡水渔业研究中心,无锡 214081) (3:江西省九江市水产科学研究所,九江 332000)
刀鲚(Coilianasus),是我国的名贵鱼类,被认为是“长江三鲜”之首,长江生态大保护的“旗帜种”之一[1]. 每年春季,溯河洄游型长江刀鲚会从沿海经河口逆江洄游至长江中下游沿线湖泊繁殖[2-4]. 由于长期受到过捕和生境破坏等人为因素的影响,其资源量已急剧下降[1],开展准确追溯其现存分布区和产卵场的工作非常困难.
鄱阳湖为我国的第一大淡水湖,湖区联通赣江、抚河、信江、饶河和修水五大水系[5]. 近年来基于耳石微化学图谱的客观确认,中国水产科学研究院淡水渔业研究中心在鄱阳湖及其相关水系中刀鲚分布区和产卵场的调查中取得了一些新突破. 研究不仅发现长江刀鲚仍可生殖洄游回到距长江口近千千米的鄱阳湖(如湖口、庐山(原星子)、都昌、余干、永修等水域)中[6-7],而且首次发现庐山-都昌(特别是麻头池到刘家山湖区)水域是一个长江溯河洄游型刀鲚的产卵场(对应于受精卵的孵化场)[7-10]并实证通湖的信江亦有该洄游型刀鲚的分布且该水系内应该也存在刀鲚产卵场[7]. 近期,国家大力实施“长江生态大保护”及 “长江十年禁渔”(2019年2月1日起农业农村部停止发放长江刀鲚、凤鲚、中华绒螯蟹专项捕捞许可证,标志着这些品种已先期开始禁渔)等一系列国策,将对长江鱼类资源及其栖息地的恢复与保护起到举足轻重的作用. 进一步基于上述耳石微化学手段来实证受惠于捕捞强度下降、生态环境向好等新形势下鄱阳湖及其联通水系所出现的新分布区或产卵场[11],无疑将对评价上述国策所产生的实际效果做出独特的贡献.
赣江是鄱阳湖流域的第一大河[12],长约760余km[13],在南昌以下分西支(主支)、中支、北支、南支入鄱阳湖; 主支在永修吴城镇附近入湖. 该河流曾发生过由于过捕、水污染、水利工程的兴建等因素,导致分布区、特别是产卵场的消失,直至长江鲥鱼灭绝的教训[14]. 有关赣江刀鲚的研究尚很缺乏,仅刘雄军等[15]在2015年5-8月对鄱阳湖、赣江和抚河下游的调查中捕获鲚属鱼类206尾; 其中长颌鲚(曾命名为C.ectenesJordan et Seale,1905)2尾(是否为赣江个体不明),被认为是洄游型刀鲚,及近10年赣江流域未发现有该类刀鲚分布[13]的报道. 实际上,长江流域刀鲚存在5个生态表型; 而长颌鲚和短颌鲚(曾命名为C.brachygnathusKreyenberg et Pappenheim,1903)均既有溯河洄游,又有淡水定居两种表型; 上颌骨长度等传统的形态分析经验并不能确证两类刀鲚是否为溯河洄游的个体. 因此,引入能够有效反映刀鲚生境履历及洄游习性的方法,来破解上述难题非常重要. 渔业环境微化学凭借其独特的学科交叉及可“破译”积累在耳石上鱼类所经历环境元素背景特性的优势[16],已被证实为能够客观、准确地反演鱼类生境履历和洄游模式的重要方法之一[17-21].
2020年5月,笔者实验室在赣江采集到刀鲚(含长颌鲚和短颌鲚2种形态型[1])共21尾. 利用上述耳石微化学技术[17-19]及刀鲚5种生态表型的分型基准[1],笔者首次对上述形态型个体的生态表型归属进行了确认并分析了其是否有溯河洄游史特征. 本研究拟为确证现今赣江中是否存在溯河洄游型刀鲚及推测其可能的产卵场; 以及利用刀鲚做为“模式”鱼种[11],尝试评价在上述生态大保护及禁渔的国策下,通鄱阳湖河流水系洄游型鱼类资源的恢复响应状况提供技术支撑和理论依据.
1 材料和方法
1.1 材料
于2020年5月27日在赣江主支炉子窑江段(29°9′N,116°0′E,距长江口约860 km)进行科研渔业特许捕捞(图1). 用单层定置丝网作业,网目20 mm,网片高度 1.2 m,单张网长50 m,3张连一张共150 m. 作业网次为1次,时间为15:00至次日6:00,共15 h. 共采集到刀鲚标本21尾(表1). 本文中按照上颌骨形态以长颌鲚(上颌骨>头长)和短颌鲚(上颌骨<头长)的方式对两类个体加以表述. 其中长颌鲚共16尾,全长为(288±25)mm,体重为(61.97±19.68)g;短颌鲚共5尾,全长为(239±42)mm,体重为(29.69±21.26)g. 鱼体标本带回实验室后-20℃冰箱冷冻保存备用.

图1 本研究刀鲚的采样位置Fig.1 Sampling location of Coilia nasus in the present study
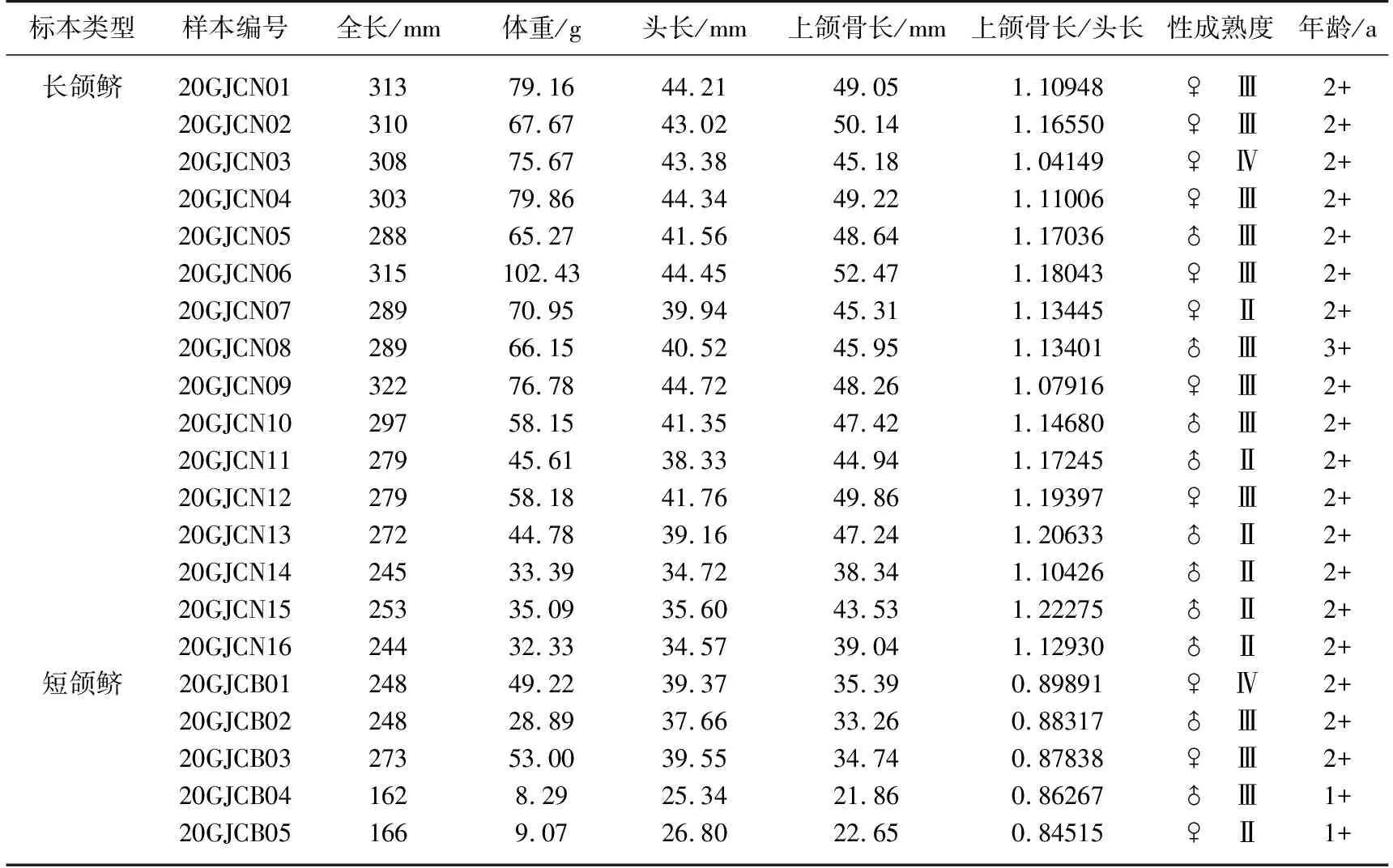
表1 赣江刀鲚标本的信息

续表1
刀鲚解冻后解剖摘取耳石. 考虑到相对于星耳石与微耳石,矢耳石体积最大,参考笔者实验室前期的方法选择左矢耳石作为研究材料,并进行前处理及微化学分析[6,20-21]. 用环氧树脂(Epofix,丹麦Struers公司)包埋耳石,将包埋耳石的树脂粘贴到玻璃片上,切割多余树脂,先用70 μm金刚砂轮(Discoplan-TS碾磨机,丹麦Struers公司)和35 μm金刚砂轮(Discoplan-TS 碾磨机,丹麦Struers公司)粗磨至暴露耳石,再用15 μm和10 μm砂纸精磨至暴露耳石核心,抛光(LaboPol-35磨抛机,丹麦Struers公司)以消除表面划痕,切割多余玻璃片. 用Milli-Q水超声清洗5 min,烘干后,用真空镀膜机(JEE420,日本电子株式会社)镀膜(36A,25s).
1.2 耳石微化学分析
基于碳酸钙(CaCO3)和钛酸锶(SrTiO3)的标准样品,使用X射线电子探针微区分析仪(简称电子探针) (EPMA,JXA-8100型,日本电子株式会社)分析耳石微化学. 即从耳石核心至耳石边缘沿最长径一条直线进行元素Sr和Ca的定量线含量分析[6,20-21],如果最长径上有划痕或者缺损,则选择次长径进行分析. 按惯例本研究计算出标准化的Sr/Ca×103比值(即本文表述的Sr/Ca比值). 随后对耳石样品进行Sr元素的面分布分析,并形成耳石过核心截面各部位Sr信号高低的16色直观图谱(黑色和红色分别反映最低和最高的浓度水平). 为方便展示以及按照惯例,根据日本电子JXA-8100操作说明在其Mapping Analysis界面下将所得Sr信号转换为相对于电子探针内标准的相对浓度(%). 上述EPMA分析过程具体的参数设定见表2.

表2 电子探针分析的条件
根据姜涛等[22]方法,对耳石矢状面先进行抛光以消除表面划痕,再用5%EDTA 酸蚀后用光学显微镜观察年轮,读取年龄.
1.3 数据处理
根据本研究室前期研究结果刀鲚耳石Sr/Ca比值及Sr含量高低图谱与其盐度生境对应关系为:淡水Sr/Ca比值≤3,低Sr蓝色图谱;河口半咸水3
利用Excel 2019计算淡水系数(freshwater coefficient,FC),即洄游性鱼类早期生活史阶段对淡水生境的依存程度[23],具体为定量线分析结果中自耳石核心开始连续对应淡水生境Sr/Ca比值≤3 的耳石径长和直到耳石最边缘点的耳石径长百分比[23-24]. 使用SPSS 22.0软件对不同生活史阶段耳石Sr/Ca比值的差异性进行非参数检验(P<0.05为显著水平,Mann-WhitneyU-test). 利用Liu等[25]的方法使用STARS(sequentialT-test analysis of regime shifts)来对定量线分析的结果进行趋势转换,设定置信度P为0.05、截断长度为5、Huber权重为1.
2 结果
2.1 赣江刀鲚耳石Sr/Ca 比及淡水系数
根据耳石读取年龄来看,长颌鲚有15尾为2+龄,有1尾为3+龄;短颌鲚有2尾为1+龄,其余3尾均为2+龄.
短颌鲚耳石定量线分析的Sr/Ca值及其趋势转换结果显示,所有5尾个体耳石从核心到边缘Sr/Ca比值的类型相同,均处于<3的低水平(图2).

图2 赣江5尾短颌鲚个体耳石Sr/Ca 比(黑窄线)和其趋势转换结果(灰宽线) (图中两条水平虚线分别对应Sr/Ca比值 为3和7. 竖直虚线对应年轮. S表示海水, B表示河口半咸水,F表示淡水)[6,11,20] Fig.2 Otolith Sr/Ca ratio transects (black narrow line) and their shifts (grey wide line) of 5 short-supermaxilla Coilia nasus from the Ganjiang River (The two horizontal dotted lines in the figure correspond to Sr/Ca ratios of 3 and 7, respectively. The vertical dotted lines correspond to annuli. S means sea water,B means brackish water,and F means fresh water)[6,11,20]
16尾长颌鲚的Sr/Ca值类型富有变化,基于数值趋势转换的结果,从核心至边缘可见显著的由低到高再降低的3个变化阶段(图 3). 表3和图3的结果显示,随个体的不同,其耳石Sr/Ca值第1阶段分别为从耳石核心沿矢状面向边缘延伸至710、740、930、1170、970、1170、750、1360、940、820、760、1120、970、650、880、680 μm半径长(即径长,下同)的范围. 其耳石区域的Sr/Ca 较低(<3). 20GJCN03、20GJCN08和20GJCN15样品在第1阶段进入第1年轮,其余样品在第2阶段进入第2年轮. 之后的第 2 阶段随个体的不同,分别为距核心向边缘方向的径长710~2010、740~1920、930~2020、1170~2040、970~2060、1170~2100、750~1790、1360~1900、940~1930、820~1840、760~2350、1120~2090、970~1960、650~1690、880~1760、680~1540 μm的范围,Sr/Ca 比显著升高(>3). 其中20GJCN07、20GJCN10、20GJCN11、20GJCN13该阶段的Sr/Ca比值甚至大于7. 除个体20GJCN14外,接下来的第3阶段,其耳石区域的Sr/Ca 比有显著降低(多数大于3). 除了样品20GJCN08和20GJCN11,其余样品在第2阶段末期和第3阶段前期进入第2年轮. 16尾长颌鲚耳石均显示 Sr/Ca 比变动的第1阶段与第2阶段、第2阶段与第3阶段间存在显著差异(P<0.05). 而只存在两个阶段的20GJCN14,第1阶段和第2阶段也存在显著差异(P<0.05).

表3 赣江刀鲚耳石Sr/Ca比值的变化

图3 赣江16尾长颌鲚个体耳石 Sr/Ca 比(黑窄线)和趋势转换结果(灰宽线) (图中两条水平虚线分别 对应Sr/Ca比值为3和7. 竖直虚线对应年轮. S表示海水,B表示河口半咸水,F表示淡水)[6,11,20] Fig.3 Otolith Sr/Ca ratio transects (black narrow line) and their shifts (grey wide line) of 16 long-supermaxilla Coilia nasus from the Ganjiang River (The two horizontal dotted lines in the figure correspond to Sr/Ca ratios of 3 and 7,respectively. The vertical dotted lines correspond to annuli. S means sea water, B means brackish water,and F means fresh water)[6,11,20]
此外,长颌鲚的FC为0.47±0.10,短颌鲚的FC均为1.00,长颌鲚与短颌鲚FC之间的差异极显著(P<0.01)(图4).

图4 赣江刀鲚(长颌鲚和短颌鲚形态型) 的淡水系数(不同字母代表差异极显著, P<0.01,one-way ANOVA)Fig.4 The freshwater coefficient of long- supermaxilla and short-supermaxilla Coilia nasus from the Ganjiang River (Those with different alphabet letters are significantly different at P<0.01,one-way ANOVA)
2.2 赣江刀鲚耳石Sr含量图谱
短颌鲚耳石从核心到边缘整个截面均呈现的是低Sr的蓝色图谱(图5). 长颌鲚相应截面颜色图谱富于变化(图6). 首先是耳石核心及附近显示出低Sr的蓝色区域. 与上述短颌鲚的图谱相似. 随之则出现了明显高Sr含量的绿色、黄色、甚至更高的红色图谱(如20GJCN10和20GJCN11). 除20GJCN14外,其余长颌鲚个体耳石的边缘部分还可变化回低Sr的蓝色图谱. 从两类刀鲚耳石整体图谱来看,后者的不同颜色分色图谱和前者单纯蓝色图谱的差别极为显著. 目视观察即可截然区分. 所有耳石经酸蚀后可清晰观察到年轮特征(图7).

图5 赣江短颌鲚耳石Sr含量的面分析图谱Fig.5 X-ray intensity maps of Sr content in the otolith of short-supermaxilla Coilia nasus from the Ganjiang River

图6 赣江长颌鲚耳石 Sr 含量的面分析图谱Fig.6 X-ray intensity maps of Sr content in the otoliths of long-supermaxilla Coilia nasus from the Ganjiang River

图7 刀鲚耳石相同过核心矢状面电子探针Sr面分布测定(A)及光学显微镜反射光观察(B)下的截面图 (a为从核心到边缘耳石生境Sr含量变化; b为从核心到边缘的年龄变化 (核心用箭头表示; 年轮用圆圈表示). a和b是耳石上的同一区域)Fig.7 The same sagittal section map through the core of otoliths for Coilia nasus from the Ganjiang River under Sr content measurement by electron probe analysis (A) and reflected light observation by optical microscope (B), repectively (The map a shows the variation of habitat Sr content from core to edge in otolith; the map b shows the change of annuli from core to edge in otolith (core: arrowhead; annuli: circled). The maps a and b show the same area in otolith)
3 讨论
淡水和海水的元素组成及背景含量有着显著的不同,如元素Sr在海水中的背景含量可高至淡水背景值的100倍[26]甚至5000倍[27]. 鱼类在经历这些不同盐度水域生活史过程时,耳石中所形成对应区域的Sr/Ca比值高低或Sr含量图谱颜色的转换会发生规律性的变化[11,28],从而可用于客观准确地重建其洄游的生境履历. 基于多年对海区、江河和湖区刀鲚耳石微化学特征及对应不同盐度生境的关联性研究,笔者研究室发现刀鲚耳石Sr/Ca比值及Sr含量高低图谱与其盐度生境一般存在以下的规律[6,11,20,29],即淡水(盐度<5,下同):Sr/Ca比值≤3,低Sr蓝色图谱;河口半咸水(5~25):3
从前述本研究的结果可见,所采得短颌鲚类群耳石全径长的Sr/Ca比值均<3,对应的图谱均呈低Sr的蓝色. 这些都呈现淡水生境履历的特征,且FC为1.00显示出其全生活史均在淡水中度过;因此,可以确定这些个体均为典型淡水定居型生态表型. 与其相对照,本研究所采长颌鲚类个体耳石的中部区域都有从核心到700~1360 μm不等径长,较大范围的低Sr/Ca比值(<3)的区域. 对应的图谱均呈低Sr的蓝色. 这表明上述个体均在淡水生境中孵化(对应其亲本在淡水生境中产卵),相应地早期生活史阶段需要在淡水中发育的状况. 需要注意的是,在这范围之后,耳石上则会转换出现从数百到数千微米间隔的Sr/Ca比值(>3)的区域,且对应的Sr含量图谱则变换成了Sr含量水平显著增高的绿色、黄色、甚至红色图谱. 根据之前的大量研究不难发现[6,11,20,29],这些海水的生境履历表明上述个体在发育到对应的生活史阶段已经入海,在半咸水或海水栖息地中继续发育和生长. 多数个体在第1冬龄之前离开淡水,进入河口半咸水,但20GJCN03、 20GJCN08和20GJCN15个体在第1冬龄之后离开淡水,进入河口半咸水. 离开淡水生境后,多数个体会在河口或者近岸半咸水水域栖息,而部分个体(如20GJCN07、20GJCN10、20GJCN11和20GJCN13)会继续向远岸高盐度海域迁徙. 及至第2冬龄后,除20GJCN08外均已启动生殖洄游. 与其他个体相比,20GJCN08作为本研究中唯一1尾3+龄个体,其于第3冬龄后启动生殖洄游,这种情况与袁传宓[30]所报道的刀鲚(长颌鲚)1~3冬龄时启动生殖洄游的情况相符. 由于这些个体又都在距离长江口近千千米的赣江水域被捕获,故这些长颌鲚均为典型的溯河洄游型刀鲚生态表型个体,同时该批鱼成熟度为II~IV期(恢复阶段至后成熟阶段)[31],因此可认为是为生殖洄游进入赣江的亲鱼. 从赣江所孵化起源的刀鲚(长颌鲚)仔幼鱼可能会对鄱阳湖、长江干流、长江口及相邻海区资源量的补充起到不可忽视的作用.
值得注意的是,20GJCN14刀鲚(长颌鲚)个体耳石边缘部分Sr/Ca比值结果显示依旧为>3的半咸水生境特征,其原因应该与此溯河洄游长颌鲚亲鱼迅速上溯[32]以及耳石上Sr/Ca比值沉积有一定的时滞有关[33]. 相似情况在本研究室前期长江安庆段(毗邻鄱阳湖)该相同生态表型刀鲚耳石微化学研究中也有发现[34]. 此外,20GJCN07个体耳石微化学结果显示耳石后端与翼叶部分Sr含量异常偏低. 类似情况在Tzeng等[35]所研究的鳗鲡耳石中也有报道. 其原因主要为矢耳石正常耳石晶体结构为文石,由于晶体结构发生变化,由文石变成了球文石,导致Sr/Ca比值异常偏低[35]. 因此,在今后的研究中有必要利用诸如拉曼光谱分析等技术进一步分析该个体耳石上异常区域的晶体结构,以确认其值异常的原因. 同时有鉴于此,笔者建议今后在进行基于耳石微化学分析的鱼类生境履历反演研究时,除了有耳石轴上的定量线分析外,有必要对耳石进行全剖面的面分布分析,以避免因晶体结构异常等原因而导致生境履历反演时发生严重误判.
本研究的耳石微化学结果首次直观地确认和实证了目前赣江有溯河洄游型刀鲚(溯河洄游长颌鲚生态表型)分布的现状. 值得注意的是,本研究所获长颌鲚绝大部分为溯河洄游型刀鲚. 其占总刀鲚标本尾数的比率达76%,不仅远高于淡水定居型短颌鲚的比率(24%);亦远高于迄今文献中所报道鄱阳湖及其通湖水系调查所获洄游型或全部长颌鲚数占总尾数的比率,如3.6%(鄱阳湖、赣江和抚河)、0.97%(鄱阳湖、赣江和抚河下游)、0.16%(鄱阳湖)[15]、15%(信江)[36]等. 该现象显示出,虽然赣江流域已近10年未发现有溯河洄游刀鲚[13],但得益于上述长江生态大保护,特别是从去年开始实施的长江禁渔的国策,赣江水域洄游型刀鲚资源量和栖息地有恢复向好的趋势. 由于这些个体均自长江口外海区沿江入湖而来;因此,本研究的结果也可从某种意义上反映出,作为刀鲚重要洄游通道、分布区甚至产卵场的长江干流及鄱阳湖[8,37],其中相应的资源量和栖息地应该同样具有恢复向好的趋势. 相关的刀鲚资源跟踪、评价和其栖息地验证研究急需尽快得到开展.
本研究所采赣江洄游型长颌鲚虽为2龄以上,但除20GJCN03个体为Ⅳ期外,无论雌、雄,其性成熟度均仅为Ⅱ~Ⅲ期;远未到产卵繁殖(Ⅴ期)的阶段. 这种情况与我们之前发现并定位的庐山市(原星子县)周边水域刀鲚产卵场情况完全不同(繁殖度Ⅲ~Ⅵ期)[8],而与信江刀鲚 (Ⅱ~Ⅲ)的情况相类似[7]. 笔者推测,这些个体溯江繁殖的目的地产卵场尚未到,即赣江中存在洄游型刀鲚未知的产卵场,但其相应的产卵场可能在采样点上游附近或更远的江段中合适水域,这在今后的工作中需要深入调查. 水利工程是影响洄游型鱼类洄游通道和产卵场最主要的人为因素之一[38]. 目前距本研究采样点最近的是在建的江西赣江丰城龙头山水电站枢纽[39-40](约130 km上游)及2019年底建成的江西赣江新干航电枢纽[41](约180 km上游). 这两座大型枢纽水库虽拟或建有仿生态鱼道[40,42],但其效果尚有待确认;因此,近期宜重点进行赣江这些枢纽以下江段洄游刀鲚分布及产卵场的定位调查,并开展这些枢纽及下游河道挖沙等人类经济开发活动对洄游刀鲚分布区、洄游通道、产卵场潜在影响的评价,以便对赣江水域该名贵鱼种资源及其栖息地环境进行有效保护. 同样需要注意的是,拟建的大型鄱阳湖水利枢纽工程建拦水闸位置选定在靠近该湖入长江水道的屏风山与长岭山间(其下游距长江仅27 km)[43]. 中国水产科学研究院淡水渔业研究中心迄今所发现的鄱阳湖内(如庐山/星子、都昌、永修、鄱阳、余干等)及其通湖赣江、信江溯河洄游型刀鲚的分布区或产卵场几乎全在闸址的上游[6-11,32,36]. 该水利枢纽工程的建设有调枯不控洪的原则和3月上中旬-8月31日泄水闸门全部敞开,江湖连通的水位调度方案. 这类措施对鄱阳湖洄游型刀鲚意义重大,即如果不能确保闸口适宜通江过鱼环境的话,自海溯江入湖的刀鲚将无法到达这些关键栖息地,则上述分布区或产卵场功能将受到破坏性影响. 因此,从赣江、鄱阳湖、长江、长江口及口外沿海等水域洄游刀鲚资源需联动保护的角度来看,鄱阳湖水利枢纽工程的建设需要高度重视闸址有效的江湖连通性.
4 结论
1) 耳石微化学结果首次发现并确证了目前赣江有溯河洄游型刀鲚分布,为把握在长江生态大保护及10年禁渔的国策下赣江水域洄游型刀鲚资源量和栖息地有恢复向好的趋势提供了客观理论依据.
2)溯河洄游型长颌鲚占总刀鲚标本尾数比率达76%,远高于迄今文献中所报道的鄱阳湖及其通湖水系的相关比率.
3)赣江中存在洄游型刀鲚未知的产卵场. 相应的产卵场应该位于采样点上游附近或更远的江段中合适水域.
4)宜尽快全面调查赣江下游全江段洄游型刀鲚的分布区及产卵场,并评价水利工程、河道挖沙等人类经济开发活动的潜在威胁和破坏,以对刀鲚资源及关键栖息地进行抢救性保护.
猜你喜欢
水利水电快报(2022年7期)2022-07-18
中国水利(2022年3期)2022-03-08
中华养生保健(2020年7期)2020-11-16
现代出版(2019年6期)2020-01-14
创作评谭(2018年5期)2018-11-07
老友(2018年7期)2018-08-08
养生保健指南(2017年12期)2017-12-07
中国诗歌(2017年12期)2017-11-15
科学生活(2017年9期)2017-10-16
