
骆驼乳清蛋白对热应激大鼠肝脏炎症及高迁移率族蛋白B1/Toll样受体4/核转录因子-κB信号通路的调节
2021-11-05 03:28乌恩吉雅杜冬华马雪妮哈斯苏荣
动物营养学报 2021年10期
乌恩吉雅 杜冬华,2 马雪妮 哈斯苏荣,3*
(1.内蒙古农业大学兽医学院,农业农村部动物疾病临床诊疗技术重点实验室,呼和浩特 010018;2.河北北方学院动物科技学院,张家口 075131;3.内蒙古骆驼研究院,阿拉善 750300)
热应激(heat stress, HS)是热带地区及高温季节最严重的应激源之一,现已成为危害全世界动物生产的重要因素,亦对动物福利造成了重大影响[1]。持续暴露于高温环境会导致动物各种应激反应,如体核温度升高、采食量减少、营养物质消化率和代谢改变等,进而增加对各种疾病的易感性[2]。当HS引起的体温升高和代谢过度时,活性氧(reactive oxygen species, ROS)的产生就会增加,进而激活炎症信号通路并诱导炎症细胞因子的分泌和激活凋亡[3]。此外,肝脏损伤是HS病例中最致命的并发症,也是导致患病动物死亡的直接原因[4]。因此,抑制炎症反应对于预防HS所致肝脏损伤并保护机体健康至关重要。
Toll样受体(Toll-like receptor,TLRs)家族成员是天然免疫和获得性免疫反应之间的主要联系,其中Toll样受体4(TLR4)可通过识别外源性或内源性配体介导免疫细胞活化、炎症细胞因子释放和组织损伤所需的主要受体[5]。高迁移率族蛋白1(high-mobility group box 1, HMGB1)属于HMG家族成员,是一种非组蛋白结合蛋白,在介导晚期炎症反应中发挥着关键作用[6]。在生理条件下,HMGB1位于细胞核与DNA结合,以稳定染色体结构,并调节转录和翻译。然而,在病理条件下,如发生肝脏损伤、细胞损伤及坏死时,可诱导HMGB1从细胞核移位到细胞外间隙激活核转录因子-κB(nuclear factor kappa-B,NF-κB)信号通路,通过诱导细胞因子释放和白细胞募集维持炎症微环境,最终引起组织损伤[6-9]。最近,Geng等[4]进一步证实HMGB1是TLR4的上游信号分子,腹腔注射HMGB1抗体可通过TLR4信号通路抑制HS诱导的大鼠肝脏炎症反应,从而缓解肝脏损伤。因此,靶向HMGB1/TLR4/NF-κB信号通路可能是预防HS所致肝脏炎症的有效途径。
骆驼乳清蛋白(camel whey protein, CWP)由α-乳白蛋白(α-LA)、乳铁蛋白(LF)、血清白蛋白(SA)、乳过氧化物酶(LPO)等组成,但缺乏易引起幼龄动物过敏反应的β-乳球蛋白(β-LG)[10]。据报道,骆驼乳清蛋白3(CWP3)可通过抑制促炎细胞因子分泌来促进糖尿病小鼠皮肤伤口的愈合,且其治疗效果优于牛乳[11-12]。膳食补充CWP亦可抑制NF-κB信号通路的激活,从而抑制HS小鼠淋巴细胞分泌促炎细胞因子[2]。本课题组前期在体外试验中也证实,经过模拟胃肠消化的CWP可通缓解氧化应激而抑制NF-κB信号通路,降低HS诱导的大鼠肝细胞炎症反应、凋亡和损伤[13]。上述研究提示,CWP在调节HS诱导的肝脏炎症反应方面具有潜在的独特作用。然而,迄今为止,CWP在动物模型中对HS所致肝脏炎症作用及机制的研究尚未见报道,对HMGB1/TLR4/NF-κB信号通路的调节作用亦有待探索。因此,本研究在前期研究的基础上,探讨了CWP对HS所致大鼠肝脏炎症的保护作用,并进一步探讨了CWP对炎症相关信号转导途径的影响。
1 材料与方法
1.1 药品和试剂
CWP由本实验室按先前报道的方法制备并保存[13];大鼠肿瘤坏死因子-α(TNF-α)、白细胞介素-1β(IL-1β)、白细胞介素-6(IL-6)、白细胞介素-8(IL-8)和白细胞介素-10(IL-10)含量酶联免疫吸附试验(ELISA)检测试剂盒购于厦门仑昌硕生物科技有限公司;Western及IP细胞裂解液购于上海碧云天生物技术公司;兔抗HMGB1抗体(货号:ab18256)、兔抗TLR4抗体(货号:ab217274)、兔抗β-actin抗体(货号:ab8227)购于艾博抗(上海)贸易有限公司;兔抗磷酸化核转录因子κB p65 (p-NF-κB p65)抗体(货号:3033)购于Cell Signaling Technology (CST)公司;辣根过氧化物酶(HRP)标记羊抗兔免疫球蛋白G(IgG)二抗购于天津三箭生物技术有限公司;乙二胺四乙酸(EDTA)抗原修复液(pH 8.0)、3,3-二氨基联苯胺四盐酸盐(DAB)显色试剂盒购于北京索莱宝生物科技有限公司;底物化学发光(ECL)化学发光检测试剂盒购于苏州宇恒生物科技有限公司;蛋白酶抑制剂、蛋白定量用Pierce BCA Protein Assay Kit购于赛默飞世尔科技(中国)有限公司;大鼠维持饲料购于某生物技术有限公司。
1.2 试验设计
6周龄SPF级雄性SD大鼠适应性饲养2周后随机分为6组:HS致肝脏损伤组(HS组)、CWP低剂量干预组(L组)、CWP中剂量干预组(M组)、CWP高剂量干预组(H组)、正常对照组(Control组)和CWP对照组(CWP组)。每组设置5个重复,每个重复6只,试验期22 d。L、M和H组大鼠每天灌服CWP(CWP溶于1 mL生理盐水,剂量分别为100、200和400 mg/kg BW),Control和HS组每天灌服1 mL生理盐水,CWP组每天灌服400 mg/kg BW的CWP,连续2周;之后,按本实验室先前建立的方法进行HS处理:除Control和CWP组外,其他各组大鼠于人工气候箱[温度为(40.0±0.2) ℃,相对湿度为60%~65%]内进行HS处理2 h(10:00—12:00),连续处理8 d。每次HS处理结束后,立即将大鼠移出人工气候箱并于初始环境饲养24 h。HS处理期间,于每次HS前1 h灌服上述剂量CWP或生理盐水。
1.3 免疫印迹(Western blotting)检测HS大鼠肝脏HMGB1、TLR4及p-NF-κB p65蛋白表达
取大鼠肝脏组织,用Western及IP细胞裂解液提取细胞总蛋白,按常规方法进行变性聚丙烯酰胺凝胶(SDS-PAGE)电泳后将蛋白转移至聚偏二氟乙烯(PVDF)膜。将PVDF膜用QuickBlockTMWestern封闭液室温封闭1 h后于稀释的HMGB1(1∶2 000)、TLR4(1∶1 000)、NF-κB p65(1∶1 000)、p-NF-κB p65(1∶1 000)及β-肌动蛋白(β-actin)(1∶10 000)抗体4 ℃孵育过夜。蛋白表达水平通过Quantity One ChemiDocXRS图像采集系统及Image-Pro Plus软件进行分析,结果用HMGB1、TLR4、p-NF-κB p65光密度值/β-actin光密度值表示。剩余大鼠肝脏组织分为2部分,一部分固定于4%多聚甲醛,另一部分于液氮冷冻后置-80 ℃保存,备用。
1.4 免疫组织化学法检测HS大鼠肝脏HMGB1、TLR4及p-NF-κB p65蛋白表达
4%多聚甲醛固定后的组织块经石蜡包埋并制成4 μm切片,每只大鼠制备3张连续切片,按免疫组化试剂盒说明书操作检测HMGB1、TLR4及p-NF-κB p65蛋白表达水平及细胞内定位。每张切片于400×显微镜下随机选择10个视野,结果分析采用积分综合计量法。利用Image J软件的IHC Profiler插件对染色强度进行自动评分:0(阴性)、1(弱阳性)、2(中度阳性)和3(强阳性)。此外,根据阳性细胞百分比进行计分:0(<5%)、1(5%~25%)、2(25%~50%)、3(50%~75%)和4(>75%)。然后,按文献[14]描述方法计算,免疫组化评分=阳性细胞数百分比分值×染色强度分数。
1.5 ELISA检测HS大鼠肝脏炎症细胞因子含量
取1.3中冻存的大鼠肝脏组织,按试剂盒说明说操作检测大鼠肝脏组织匀浆中TNF-α、IL-1β、IL-6、IL-8和IL-10含量。
1.6 数据统计分析
所有试验数据均以平均值±标准误表示,用SPSS 25.0软件的单因素方差分析(one-way ANOVA)和LSD法进行组间比较,结果以P<0.05表示差异显著,P<0.01表示差异极显著,P<0.001表示差异极其显著。
2 结果与分析
2.1 Western blotting法分析CWP干预对HS大鼠肝脏HMGB1/TLR4/NF-κB信号通路的影响
如图1所示,HMGB1/TLR4/NF-κB信号通路参与了HS所致大鼠肝脏损伤过程中的炎症级联反应调控。结果表明,HS大鼠肝脏中TLR4和p-NF-κB p65蛋白表达水平极其显著上调(P<0.001),相比于Control组分别上调了8.50和10.17倍,但HMGB1蛋白表达水平未发生显著变化(P>0.05)。CWP(200、400 mg/kg)可极显著和极其显著抑制p-NF-κB p65(P<0.01)和TLR4(P<0.001)蛋白表达水平。然而,100、200、400 mg/kg CWP同样没有显著影响HMGB1蛋白表达水平(P>0.05)。
2.2 免疫组织化学法分析CWP干预对HS大鼠肝脏HMGB1、TLR4及p-NF-κB p65蛋白表达的影响
如图2所示,免疫组化结果与2.1描述一致,不同的是,虽然HS对HMGB1蛋白表达水平无显著影响(P>0.05),但可使肝细胞核中HMGB1蛋白表达水平降低(图2-A,如红色箭头所示),胞浆中表达水平升高(图2-A,如黑色箭头所示)。有趣的是,虽然CWP对HMGB1蛋白免疫组化评分亦无显著影响(P>0.05,图2-B),但CWP可逆转HMGB1蛋白的核转位现象,且400 mg/kg CWP几乎完全恢复了HMGB1蛋白的细胞核定位(图2-A)。CWP(400 mg/kg)可极显著抑制TLR4蛋白表达(P<0.01,图2-C和图2-D)。此外,免疫组化评分结果显示,400 mg/kg CWP亦极其显著地恢复了p-NF-κB p65蛋白的胞浆表达(P<0.001),表明CWP可抑制其核转位(图2-D和图2-E,如绿色箭头所示)。这些结果进一步证实,CWP对HS大鼠肝脏HMGB1/TLR4/NF-κB信号通路具有调控作用,综合分析,400 mg/kg剂量效果较好。
2.3 CWP干预对HS大鼠肝脏炎症细胞因子含量的影响
炎症反应是介导HS所致肝脏损伤的主要因素之一。因此,本研究采用ELISA方法检测了各组大鼠肝脏TNF-α、IL-1β、IL-6、IL-8和IL-10含量,以确定HS前灌服CWP是否可抑制肝脏损伤过程中的炎症反应。由表1可见,HS组大鼠肝脏TNF-α、IL-1β、IL-6、IL-8含量均显著升高(P<0.05),而IL-10含量显著降低(P<0.05)。HS前灌服400 mg/kg CWP可显著降低TNF-α、IL-1β、IL-6、IL-8含量(P<0.05),显著提高IL-10含量(P<0.05)。此外,CWP表现出使正常大鼠肝脏中上述细胞因子含量升高趋势,但与Control组相比差异不显著(P>0.05)。
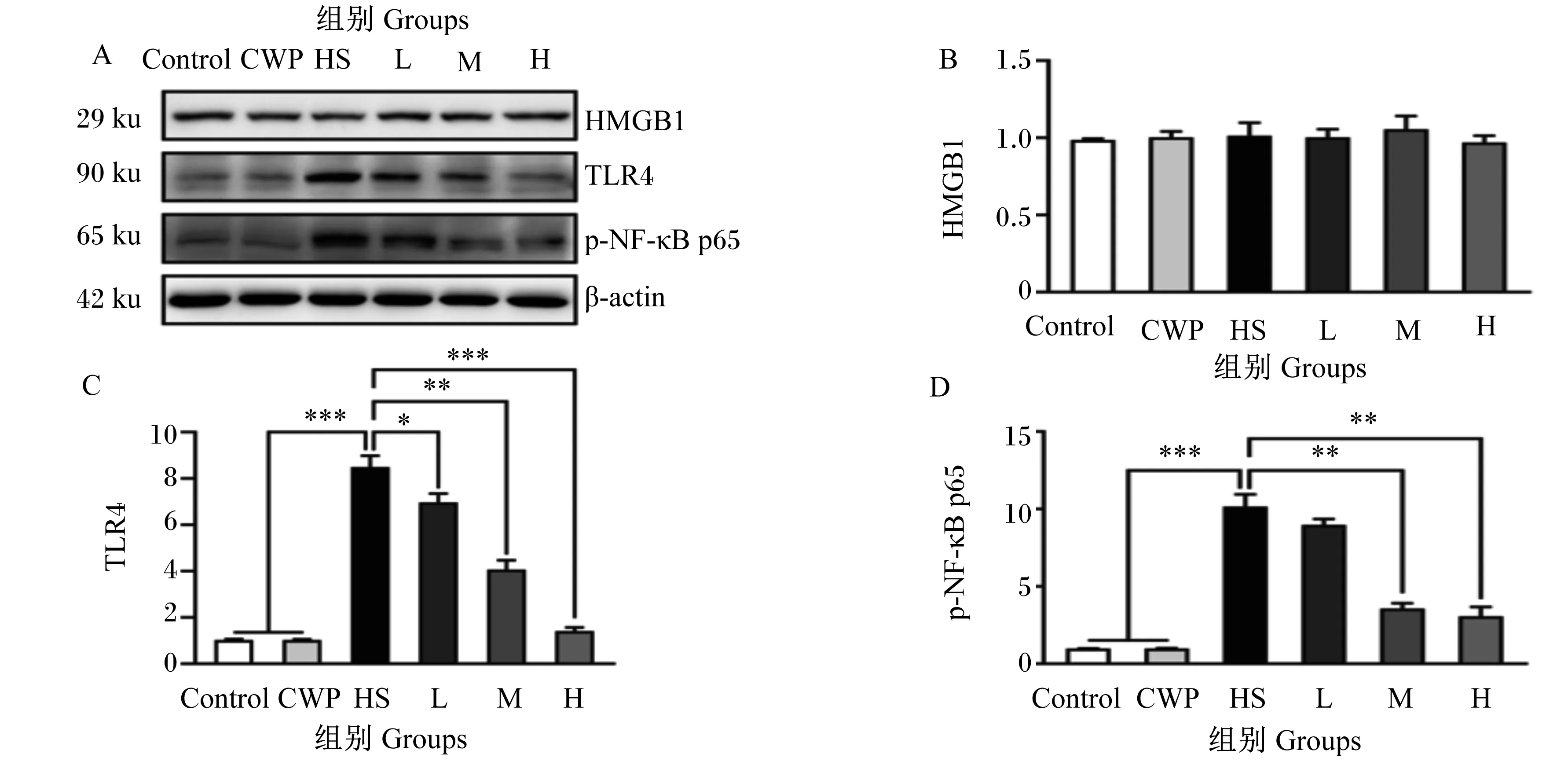
HMGB1:高迁移率族蛋白B1 hepatic high mobility group protein B1; TLR4 :Toll样受体4 Toll like receptor; 4p-NF-κB p65: 磷酸化核转录因子κB p65 phosphorylated nuclear factor-κB;β-actin:β-肌动蛋白。Control: 正常对照组 control group; CWP: 骆驼乳清蛋白组 CWP group; HS: 热应激致肝脏损伤组 HS group; L: 骆驼乳清蛋白低剂量干预组 L group; M: 骆驼乳清蛋白中剂量干预组 M group; H: 骆驼乳清蛋白高剂量干预组 H group。*表示差异显著(P<0.05),**表示差异极显著(P<0.01),***表示差异极其显著(P<0.001) * indicates significant difference (P<0.05), ** indicates highly significant difference (P<0.01), *** indicates extremely significant difference (P<0.001)。下图同 the same as below.
3 讨 论
HS已在全球范围内严重影响了动物健康和生产,其实质是动物机体产生的热量超过了其将热量散发到周围环境的能力。发生HS的动物会倾向于通过减少采食量来减少产热,由此对生长性能产生负面影响[1,4,15]。越来越多的证据表明,肝脏损伤是几乎所有HS病例中最常见且致命的并发症[16-18]。此外,尽管HS是由高温引起的,但即使恢复HS动物模型体温也不能阻止炎症反应,进而使肝脏损伤在HS恢复过程中才逐渐显现[19]。因此,HS所致肝脏损伤可能不是高温作用的直接结果,而是由后期持续的炎症反应所介导的[19]。已有研究证实,CWP具有显著的抗炎作用[11-12,20],且其水解产物具有抗肝癌细胞(HepG2)炎症反应活性[21],表明其具有抗HS致肝脏损伤潜能。另据报道,糖尿病小鼠补充CWP可通过抑制促炎细胞因子分泌来促进组织修复,从而加速皮肤伤口的愈合[11-12]。本课题组前期试验亦表明,CWP可通过抑制炎症信号通路缓解HS所致大鼠肝脏细胞损伤及凋亡,进一步证实了CWP抗HS致肝脏损伤的潜在医用价值,但其抗炎机制有待深入研究[13]。因此,本研究探讨了CWP对HS大鼠肝脏炎症的作用及可能机制。
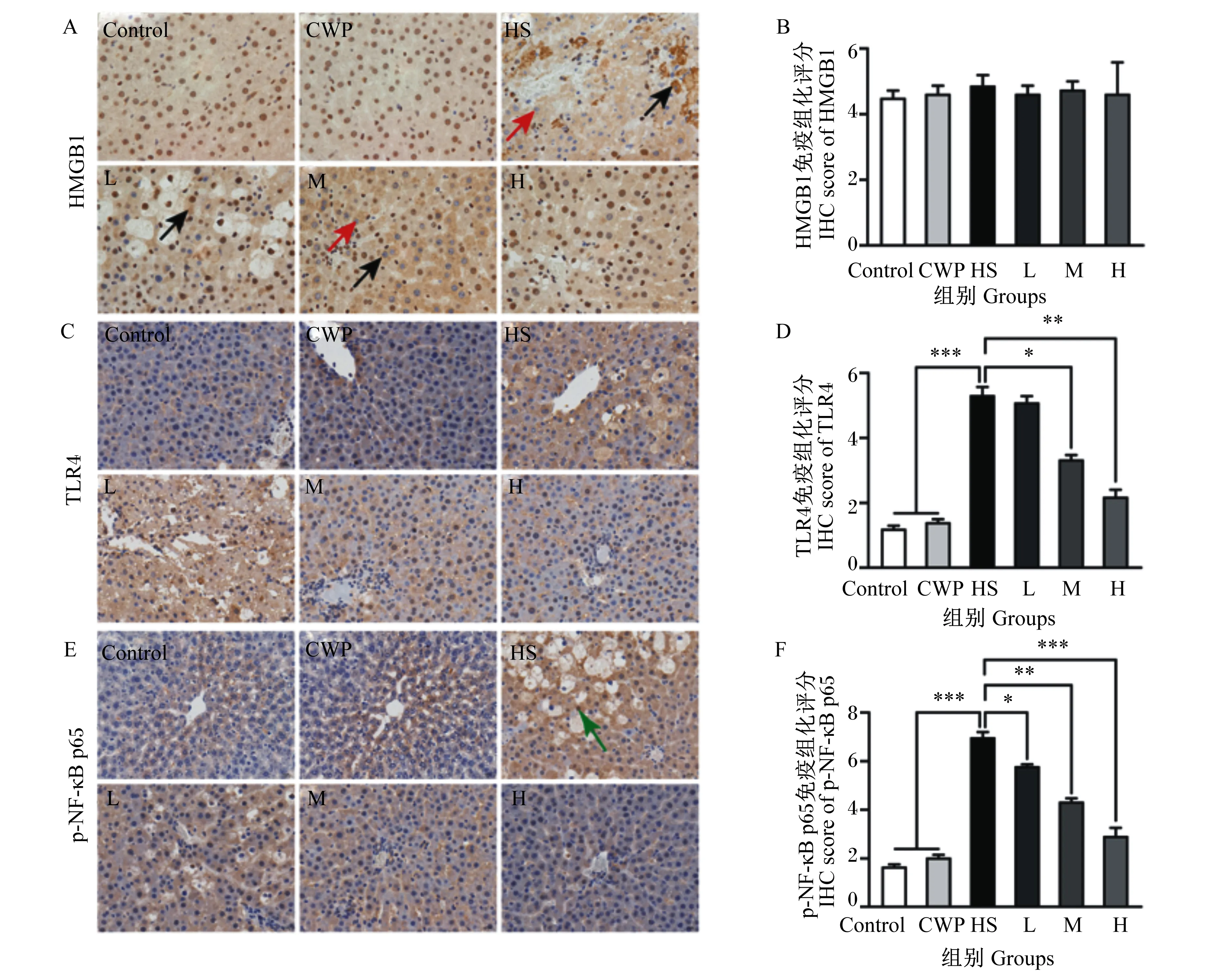
图A和图B:用免疫组化法分析HMGB1蛋白表达;图C和图D:用免疫组化法分析TLR4蛋白表达;图E和图F:用免疫组化法分析p-NF-κB p65蛋白表达。黑色箭头:目的蛋白在胞浆中表达增多;红色箭头:目的蛋白在胞核中表达降低;绿色箭头:目的蛋白在胞核中表达升高。
最近的研究表明,在无菌性感染相关的炎症反应中,HMGB1是一种重要的炎症介质。在生理条件下,HMGB1位于细胞核并与DNA结合,以稳定染色体结构、调节转录和翻译。然而,在病理条件下,如肝细胞损伤及坏死时,可诱导HMGB1从细胞核移位到细胞外间隙激活NF-κB信号通路,从而促进炎症细胞因子分泌[6-7]。另一项研究亦证实,当肝脏损伤发生时,HMGB1蛋白可被诱导表达并通过受损细胞被动释放至细胞外,或者从激活的免疫细胞中主动分泌[9]。被释放至胞外的HMGB1可通过TLR4激活NF-κB信号通路,诱导促炎细胞因子分泌,从而促进炎症发展[22]。因此,HMGB1/TLR4/NF-κB轴在介导各种应激因素诱发的炎症反应中发挥着重要作用。Yin等[23]研究显示,通过调节HMGB1/TLR4/NF-κB信号通路可改善炎症所致小鼠肝脏损伤。本研究发现,HS大鼠肝脏中TNF-α、IL-1β、IL-6、IL-8含量均显著升高,IL-10含量显著降低。相应地,HS大鼠肝脏中TLR4和p-NF-κB p65蛋白表达水平均高于正常对照组,HMGB1胞外表达量亦明显增多。这些数据提示,HS大鼠肝脏发生了炎症反应。有趣的是,HS前灌服CWP可以剂量依赖性方式逆转TLR4和p-NF-κB p65蛋白表达的改变,伴随着肝脏IL-1β、IL-6、IL-8含量的降低和IL-10含量的升高。这与Ramadan等[2]研究结果一致,膳食补充CWP可抑制NF-κB信号通路的激活,从而降低HS诱导的小鼠淋巴细胞促炎细胞因子(TNF-α、IL-1β、IL-6)和IL-4含量。另外,Western blotting结果显示CWP不影响HMGB1蛋白表达量,然而免疫组化结果显示其抑制了HMGB1的核转位,恢
复了其胞核内蛋白表达量,这与Geng等[4]研究结果一致。因此,我们推测CWP可能并不是直接与HMGB1相互作用,而是通过修复损伤细胞降低了HMGB1的释放,导致其不能通过TLR4激活NF-κB信号通路,最后抑制了细胞因子的分泌。因为已有研究表明,CWP可通过抑制促炎细胞因子分泌来促进组织修复,从而加速糖尿病小鼠皮肤伤口愈合[11-12]。虽然该假说尚需进一步证实,但本研究仍可证实CWP对HMGB1/TLR4/NF-κB信号通路具有调节作用,CWP可能是通过调节该途径抑制了HS诱导的大鼠肝脏炎症反应。
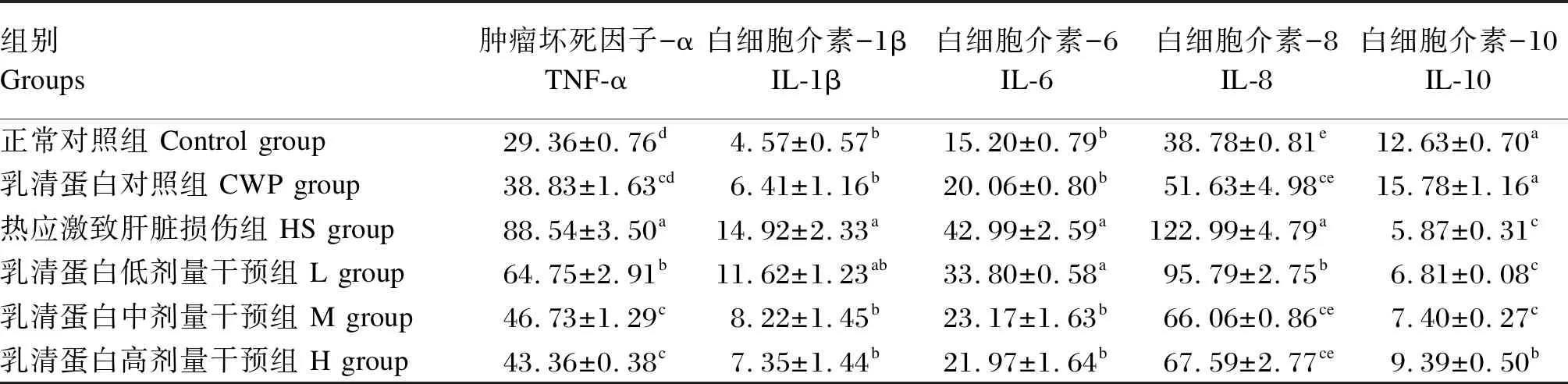
表1 CWP干预对HS大鼠肝脏细胞因子含量的影响
4 结 论
综上所述,本研究进一步证实了HMGB1在HS所致肝脏损伤中的作用,并阐明了HMGB1/TLR4/NF-κB信号通路在HS诱导肝脏炎症反应中的可能机制,提示HMGB1可能是干预HS所致肝脏损伤的潜在靶点。此外,本研究初步阐明了CWP对HS大鼠肝脏炎症及HMGB1/TLR4/NF-κB途径的调控作用。
猜你喜欢
中老年保健(2022年4期)2022-11-25
肝博士(2022年3期)2022-06-30
现代临床医学(2021年4期)2021-07-31
浙江医学(2020年9期)2020-07-01
当代水产(2019年9期)2019-10-08
浙江中西医结合杂志(2019年4期)2019-05-05
浙江医学(2019年2期)2019-01-23
安徽医科大学学报(2016年12期)2017-01-15
中西医结合心脑血管病杂志(2016年20期)2016-03-01
中国继续医学教育(2015年1期)2016-01-06
