
采前红蓝光连续光照光强对三种氮水平水培生菜AsA累积代谢的影响
2021-12-14 06:28刘家源刘文科
照明工程学报 2021年5期
刘家源,刘文科
(中国农业科学院农业环境与可持续发展研究所,农业农村部设施农业节能与废弃物处理重点实验室,北京 100081)
引言
氮素供应水平影响植株生长发育具有一定的浓度效应,氮素供应不足和过量均会抑制植株生长发育,氮素水平的高低会影响植物体内AsA的含量。研究表明,氮素营养对植物体内AsA含量没有影响或者使其增加,但是过量的氮会降低植物体内AsA的含量[1]。张玉彬等[2]研究发现,生长光强一致时,水培生菜的AsA等物质的含量随营养液氮水平的升高而增加。在作物生长过程中,协调好氮营养和光环境条件可以发挥两者对作物产量及品质提升的最佳耦合效应,进而实现作物优质高产[3]。
抗坏血酸(Ascorbic Acid, AsA),又称维生素C,是一种抗氧化剂,与其他抗氧化剂共同组成抗氧化系统,保护植物免受有氧代谢、光合作用和多种污染物造成的氧化损害[4]。AsA参与了一些植物激素如乙烯、赤霉素等物质合成代谢过程。内源性 AsA水平的高低会影响植物开花衰老相关的诱导,与开花时间、细胞的凋亡、病原体的信号转导和植物体衰老的发展都有联系[5]。光照是对AsA积累至关重要的环境因子[6]。AsA在叶片中的总浓度与光照有关。已有大量研究表明,在一定光强范围内,光强越大植物体内的AsA含量越高[7]。AsA循环再生过程中,APX、DHAR、MDHAR和GR作为抗氧化酶随着光照强度的增加而增加,从而提高AsA的含量以及植物在高光强下的抗氧化能力[8]。高光强还可通过促进光合产物生成,增加AsA合成前体,从而促进AsA的积累[9]。
连续光照通常是指改变植物原有的明暗交替的光周期规律,给植物提供连续24 h或超过24 h的光照[10]。已有研究表明,生菜体内AsA含量与供氮和光照条件密切相关,在采收前进行一定强度的短期连续光照可以显著提升水培叶菜的营养品质[11]。周晚来[12]研究发现,在生菜采收前,进行短期连续光照可以显著提高AsA等营养物质的含量。生菜是一种被人们广泛食用的全球性叶菜类蔬菜,在设施园艺中栽培广泛,也是人工光植物工厂广泛种植的代表性蔬菜[13]。本研究设想在生菜生长过程中,通过研究采前LED红蓝光连续光照光强对三种氮水平下生菜AsA含量和相关合成代谢的酶活性的影响,探究氮水平与光强对AsA含量和AsA相关代谢网络的变化机理以及对采前连续光照的响应,为植物工厂中高产优质蔬菜的生产和生产中光环境和氮营养的调控提供理论依据。
1 材料与方法
1.1 试验材料
本试验在中国农业科学院农业环境与可持续发展研究所植物工厂进行。试验采取水培方式,种植环境温度为25±1 ℃,相对湿度为65%±5%,CO2浓度为大气CO2浓度。试验以生菜(Lactucasativa)为试验材料,品种为“意大利耐抽薹”。
1.2 试验设计
将生菜种子播于海绵块中育苗,培养15 d后将长势一致的生菜苗随机移栽于长方形塑料栽培槽(长180 cm×宽60 cm×高6 cm)内,并于次日开始光照试验。试验期间采用营养液水培,营养液采用霍格兰配方。试验前期正常光照采用LED红蓝光面板灯进行光照处理,红蓝光比例为4∶1,如表1所示,前期设置2种光强(100、150 μmol·m-2·s-1,I100、I150)和3种氮水平(2.5、5、7.5 mmol/L,N2.5、N5、N7.5),通过调节营养液中硝酸钙的浓度来控制氮水平。试验首先在常规光周期(16/8 h)下栽培17 d,在第18 d开始进行连续光照处理,连续光照处理的光照强度统一调至为150 μmol·m-2·s-1。

表1 常规光周期下的试验参数
1.3 测定方法
分别于每个处理中随机选8株生菜作为重复样本,从茎基部切开,其中4株的地上部分将叶片与叶柄分离后,迅速用液氮冷冻,并用高通量组织研磨器在低温下把用液氮冷冻好的植物样品研磨成粉末,放至-80 ℃冰箱中留样备用。
AsA含量采用Gillespie[14]的方法测定。L-半乳糖酸-1,4-内酯脱氢酶(GalLDH)活性参照Li[15]的方法测定。AsA过氧化物酶(APX)活性参照Nakano[16]的方法测定。AsA还原酶(DHAR)、单脱氢AsA还原酶(MDHAR)和谷胱甘肽还原酶(GR)的活性参照Ma[17]的方法测定。
2 试验结果
2.1 采前LED连续光照光强对三种氮水平处理下生菜AsA含量的影响
由表2可得,连续光照前,N2.5I100下的AsA含量最低为0.68 mg·g-1,N7.5I150下抗坏血酸含量最高为1.37 mg·g-1。连续光照后,N5I100下的AsA含量增幅最低为12.79%,N2.5I100下的抗坏血酸增幅最高为39.71%。连续光照前后,同一光强下,AsA含量随着氮水平的升高而升高。同一氮水平下,I150下的抗坏血酸含量明显高于I100处理。连续光照前后,氮水平与光强处理下生菜叶柄的AsA含量均无显著差异。
由表3可知,连续光照前,光强与三种氮水平处理对生菜叶片的DHA含量具有显著影响。光照强度为150 μmol·m-2·s-1时,氮水平处理对生菜叶片具有显著影响,氮水平2.5 mmol/L处理下的DHA含量显著高于其他两个氮水平处理。光强与三种氮水平处理下生菜叶柄的DHA含量无显著差异。连续光照后,各处理下叶片和叶柄的DHA含量均无显著影响。可见,150 μmol·m-2·s-1的连续光照处理对水培生菜DHA的含量无显著影响。

表2 采前LED连续光照光强对三种氮水平处理下生菜AsA的影响
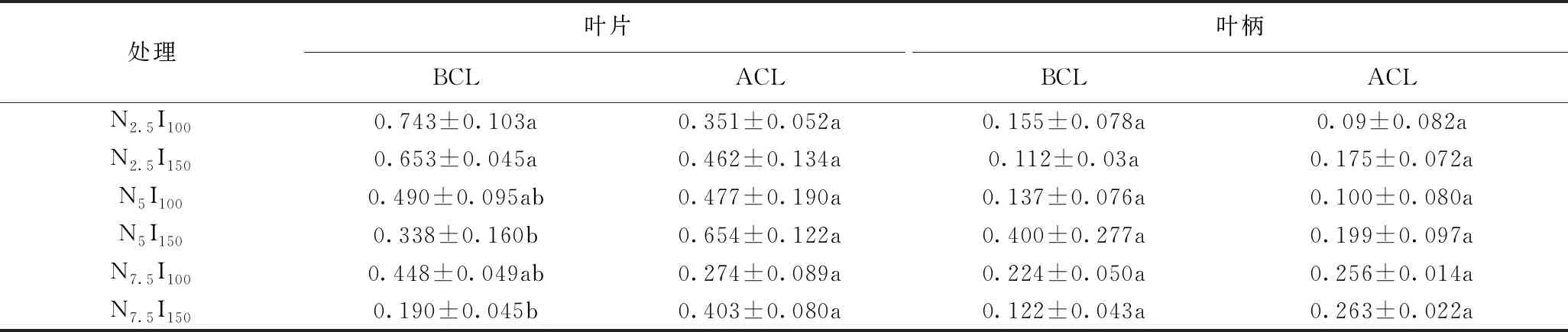
表3 采前LED连续光照光强对三种氮水平处理下生菜DHA的影响
2.2 采前连续光照光强对三种氮水平处理下生菜AsA代谢相关酶活性的影响
由表4可知,连续光照前,光强与三种氮水平处理对生菜叶片的GalLDH酶活性含量没有显著影响。N2.5I150处理下叶柄的GalLDH酶活性最大为0.209 U mg-1FW,光照强度为150 μmol·m-2·s-1时,生菜叶柄GalLDH酶活性随氮水平的升高呈现先降低后升高的趋势。连续光照后,光强与三种氮水平处理下生菜叶片和叶柄的GalLDH酶活性均无显著差异。可见,150 μmol·m-2·s-1的连续光照处理对水培生菜GalLDH酶活性无显著影响。
由表5可知,连续光照前,光强与三种氮水平处理对生菜叶片的APX酶活性含量具有显著影响。N2.5I150处理下叶片的APX酶活性最小为0.297 U mg-1FW,N7.5I150处理下叶片的APX酶活性最大达到0.468 U mg-1FW。同一生长光强处理下,氮水平2.5 mmol/L处理下生菜叶片的APX酶活性最低。连续光照后,光强为100 μmol·m-2·s-1时,氮水平对生菜叶片的APX酶活性无显著影响。光强为150 μmol·m-2·s-1时,生菜叶片的APX酶活性随氮水平的升高呈现先降低后升高的趋势。N7.5I150处理下的生菜叶片APX酶活性最大为0.784 U mg-1FW,N5I150处理下的生菜叶片APX酶活性最小为0.423 U mg-1FW。连续光照前后,各处理下的生菜叶柄的APX酶活性均无显著差异。说明150 μmol·m-2·s-1的连续光照光强及三种氮水平处理对生菜叶柄的APX酶活性无显著影响。

表4 采前LED连续光照光强对三种氮水平处理下生菜GalLDH酶活性的影响

表5 采前LED连续光照光强对三种氮水平处理下生菜APX酶活性的影响
由表6可知,连续光照前,光强与三种氮水平处理对生菜叶片的MDHAR酶活性含量没有显著影响。连续光照后,光强为150 μmol·m-2·s-1时,氮水平处理对生菜叶片的MDHAR酶活性无显著影响。光强为100 μmol·m-2·s-1时,氮水平5 mmol/L处理下生菜叶片的MDHAR酶活性显著高于其他两个氮水平处理下的。整体来看,N7.5I150处理下的生菜叶片MDHAR酶活性最大为3.56 U mg-1FW,N2.5I100处理下的生菜叶片APX酶活性最小为2.86 U mg-1FW。连续光照前后,各处理下的生菜叶柄的MDHAR酶活性均无显著差异。说明150 μmol·m-2·s-1的连续光照光强及三种氮水平处理对生菜叶柄的MDHAR酶活性无显著影响。

表6 采前LED连续光照光强对三种氮水平处理下生菜MDHAR酶活性的影响
由表7可知,连续光照前,光强与三种氮水平处理对生菜叶片的DHAR酶活性含量具有显著影响。N7.5I150处理下叶片的DHAR酶活性最小为1.35 U mg-1FW,N2.5I150处理下叶片的DHAR酶活性最大达到1.80 U mg-1FW。连续光照后,同一生长光强下,营养液氮水平对生菜叶片的DHAR酶活性无显著影响。同一氮水平下,150 μmol·m-2·s-1处理下的DHAR酶活性高于100 μmol·m-2·s-1处理。连续光照后,各处理下的生菜叶柄的DHAR酶活性均无显著差异。说明150 μmol·m-2·s-1的连续光照光强及三种氮水平处理对生菜叶柄的DHAR酶活性无显著影响。

表7 采前LED连续光照光强对三种氮水平处理下生菜DHAR酶活性的影响
由表8可知,连续光照前,光强与三种氮水平处理对生菜叶片和叶柄的GR酶活性含量具有显著影响。光强为150 μmol·m-2·s-1时,氮水平7.5 mmol/L处理下生菜叶片的GR酶活性高于其他两个氮水平处理。总体来看,连续光照前,N7.5I150处理下叶片的GR酶活性最大为0.570 U mg-1FW,N2.5I150处理下叶片的GR酶活性最小为0.367 U mg-1FW。N2.5I150处理下叶柄的GR酶活性最大为0.100 U mg-1FW,N5I100处理下叶柄的GR酶活性最小为0.053 U mg-1FW。连续光照后,各处理下的生菜叶片和叶柄的GR酶活性均无显著差异。
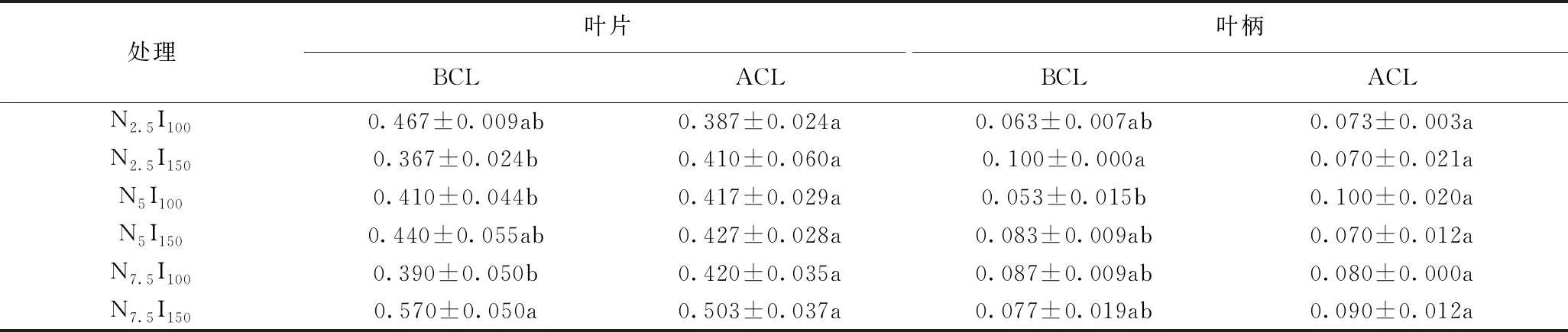
表8 采前LED连续光照光强对三种氮水平处理下生菜GR酶活性的影响
3 讨论
连续光照处理显著促进了三种氮水平水培生菜AsA的积累,AsA含量的增加主要受氮水平的影响,低氮水平时抗坏血酸含量的增幅较高,这与余意等[18]的研究结果一致。Zhou等[19]研究发现,水培生菜采收前经过72 h连续光照后,生菜叶片的AsA含量显著提高,与本试验结果一致。抗坏血酸合成代谢中的GalLDH合成酶、APX氧化酶和DHAR还原酶等的编码基因具有光诱导性, 在连续光照或强光照射处理下,相关基因的表达量增加[20], 从而使抗坏血酸合成加快。研究表明,植物体内的主要合成途径是L-半乳糖途径,Gal LDH是此途径中催化AsA合成的最后一个关键酶,在AsA的生物合成上起着至关重要的作用。GalLDH酶活性与AsA含量之间的关系会因植物种类、环境条件以及组织器官的不同而产生差异[21]。
一些研究表明,AsA含量的高低主要与APX、MDHAR、DHAR和GR等酶活性的高低有关[22]。本研究发现,连续光照前后,高氮高光强处理下生菜的AsA含量较高;连续光照后,高氮高光强处理下DHA的含量较低。连续光照前后,各处理间的叶片GalLDH酶活性无显著差异,说明在本试验中GalLDH酶对AsA的合成没有显著影响。由于APX是催化AsA氧化的酶,其活性的增加将消耗更多的AsA。然而,在许多研究[23]以及本研究中APX活性的升高往往伴随着AsA含量的增加。连续光照前后,叶片APX酶活性在高氮高光强处理下的活性更高,与AsA含量的变化趋势一致,说明氮水平和光强越高,APX酶活性越高,催化生成AsA的量越多。连续光照前,叶片MDHAR在高氮高光强处理下的活性略微升高,说明在本试验中MDHAR酶对AsA的合成没有显著影响。连续光照后,MDHAR酶活性在高氮高光强处理下的活性更高,与AsA含量的变化趋势一致,说明氮水平和光强越高,催化分解为DHA那部分的MDHA转化合成AsA的量越多。连续光照前,叶片DHAR在在高氮高光强处理下的活性降低,与AsA含量的变化趋势相反,说明在本试验中,连续光照前,DHAR酶对AsA的合成没有影响。连续光照后,DHAR酶活性在高氮高光强处理下的活性更高,与AsA含量的变化趋势一致。DHAR酶活性越高,催化分解为2,3-二酮古洛糖酸那部分DHA转化合成AsA的量越多。因此,连续光照前,氮水平与光强的影响可能是与参与AsA再生循环系统的MDHAR酶活性相关。连续光照后,氮水平与连续光照光强的影响主要是与参与AsA再生循环系统的MDHAR和DHAR酶活性相关。
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31
中老年保健(2022年6期)2022-08-19
小天使·四年级语数英综合(2022年6期)2022-06-21
中国机械工程(2022年8期)2022-05-09
作文·小学低年级(2021年4期)2021-11-02
作文小学中年级(2021年4期)2021-05-07
儿童时代·幸福宝宝(2021年1期)2021-03-29
农家参谋(2020年5期)2020-06-15
小资CHIC!ELEGANCE(2019年40期)2019-12-10
音乐教育与创作(2019年8期)2019-05-16
