
新疆动物源粪肠球菌的耐药性分析与耐药基因检测
2022-01-05 06:30陈万昭陈月月易海波徐琦琦吴慧敏夏利宁
中国农业大学学报 2022年2期
陈万昭 陈月月 易海波 秦 蕾 徐琦琦 吴慧敏 夏利宁*
(1.新疆农业大学 动物医学学院,乌鲁木齐 830052;2.新疆霍城县职业技术学校,新疆 霍城 835200;3.新疆昭苏县喀夏加尔镇畜牧兽医站,新疆 昭苏 835618)
粪肠球菌(Enterococcusfaecalis)为革兰氏阳性菌,在自然界中广泛存在,是人和部分动物肠道中的常在菌群之一,也是条件致病菌[1]。粪肠球菌可能会引发包括泌尿系统感染、感染性心内膜炎、脑膜炎和菌血症等多种严重疾病[2]。近年来,随着肠球菌对抗菌药物耐药性的增加,导致肠球菌感染治疗失败的病例屡见不鲜[3],粪肠球菌在人类临床肠球菌感染分离株中占有重要比例[2],亦成为全球范围流行的第三大院内感染病原菌[4]。粪肠球菌耐药机制复杂,具有显著的基因组可塑性,利用质粒、转座子和插入序列高效获取和转移可移动抗性元件[3],所以它可作为肠道中其他致病菌耐药基因的“蓄水池”,促进耐药性的传播[5]。
耐药粪肠球菌的传播不仅危害人类健康,还影响养殖场经济效益,一旦爆发会降低动物的生产性能,大幅增加养殖场的用药成本。目前关于我国动物源肠球菌耐药性的报道大多针对一种或两种动物,多种动物源报道较少,新疆动物源肠球菌耐药性的报道仅局限于新疆北部地区与阿克苏地区的猪源粪肠球菌[6-7]、西北部地区(伊犁和塔城等地)的动物原奶、手工乳酪中分离出的屎肠球菌[8],从药敏结果看耐药情况较为严重,而粪肠球菌在新疆多种动物肠道中的分布情况及耐药基因携带情况尚不明确。因此,本研究以新疆9个县区分离的鸡源、猪源、牛源、羊源、鸽源和骆驼源粪肠球菌为研究对象,进行耐药性调查以及相关耐药基因检测,为了解新疆多种动物源粪肠球菌的耐药现状及耐药基因携带情况,为临床用药提供科学依据,同时也为解析粪肠球菌的多重耐药机制提供数据基础。
1 材料与方法
1.1 材料
1.1.1菌株
2015年在石河子地区采集羊源肛拭子样品110份。2016年在乌鲁木齐市米东区采集鸡源肛拭子42份。2019年在昌吉地区采集猪源肛拭子90份,在阿勒泰福海县采集骆驼源肛拭子49份,在伊犁察布查尔锡伯自治县采集牛源肛拭子135份,在五家渠采集牛源肛拭子18份,在和田墨玉县采集鸽源肛拭子100份。2020年在阿勒泰布尔津县采集骆驼源肛拭子20份。采集4个年份的样品共计564份。标准质控粪肠球菌(ATCC29212)与金黄色葡萄球(ATCC29213)购自杭州天和微生物试剂有限公司。
1.1.2主要试剂和培养基
脑浸出液(BHI)肉汤培养基购自北京奥博星生物技术有限公司;MH肉汤、肠球菌选择培养基(胆汁七叶苷叠氮钠琼脂)购自青岛海博生物技术有限公司;尿道菌群定位显色培养基购自法国科玛嘉;1×TE缓冲液购自生工生物(上海)有限公司;2×TaqPCR Master Mix,D2 000 DNA Maker购自天根生化科技(北京)有限公司。
1.2 方法
1.2.1细菌分离鉴定
用灭菌棉签沾取动物肛门内新鲜粪便,放入装有1 mL MH肉汤的EP管中。吸取10 μL MH肉汤中的样品至1 mL 6.5%氯化钠BHI肉汤中,过夜培养后将增菌液划线接种于肠球菌选择培养基平板上,37 ℃培养16~20 h后,挑取白色圆滑、凸起、大小适中且菌落周围显黑色的单一菌落,置于1 mL BHI肉汤,过夜培养后将增菌液划线接种于尿道菌群定位显色培养基平板上,37 ℃培养16~20 h后挑取大小合适的蓝绿色单菌落于1 mL BHI肉汤,42 ℃培养16~20 h,初步鉴定为肠球菌。通过PCR方法对粪肠球菌持家基因进行扩增[9],采用煮沸裂解法提取分离株总DNA作为PCR模板,提取标准质控粪肠球菌(ATCC29212)的总DNA作为PCR阳性对照。扩增产物经加核酸染料的1.0%琼脂糖凝胶电泳检测后,将目的片段进行胶回收并送生工生物工程(上海)股份有限公司测序。测序结果用BLAST进行序列比对,相似性>96.0%的结果判定该菌株为粪肠球菌。
1.2.2菌株保存
确定为粪肠球菌的菌株接种在BHI肉汤隔夜培养后,加入灭菌甘油,制成20.0%甘油菌,-20 ℃冰箱保存。
1.2.3药物敏感性试验
根据美国临床和实验室标准协会(CLSI)推荐的动物源细菌抗菌药物敏感性试验琼脂稀释法执行标准进行试验操作[10],对分离的粪肠球菌进行11种抗菌药物(恩诺沙星、阿米卡星、庆大霉素、利福平、氟苯尼考、多西环素、四环素、红霉素、万古霉素、利奈唑胺和氨苄西林)的最小抑菌浓度测定。
1.2.4耐药基因的检测
根据所选药物相对应的耐药基因进行检测。参考文献[11-19]已有序列合成引物(表1),引物由生工生物(上海)有限公司合成。扩增产物经琼脂糖凝胶电泳验证后,将目的片段进行胶回收并送生工生物(上海)有限公司测序。测序结果经BLAST进行序列比对后确定基因型。
1.2.5数据分析
采用IBM SPSS Statistics 26.0进行数据分析。单因素分析采用卡方检验或Fisher精确检验,P<0.05为差异显著,P>0.05为差异不显著。
2 结果与分析
2.1 粪肠球菌的分离鉴定
由表2可知,共采集4个年份不同动物的样品共计564份,经粪肠球菌持家基因扩增分离鉴定(图1为部分结果)得到242株粪肠球菌,分离率为42.9%(242/564)。鸡源粪肠球菌的分离率(95.2%,40/42)显著高于其他动物源粪肠球菌(P<0.05),猪源粪肠球菌分离率为61.1%(55/90),牛源粪肠球菌(35.9%,55/153)、羊源粪肠球菌(34.5%,38/110)、鸽源粪肠球菌(34.0%,34/100)与骆驼源粪肠球菌(29.0%,20/69)的分离率差异不显著(P>0.05)。
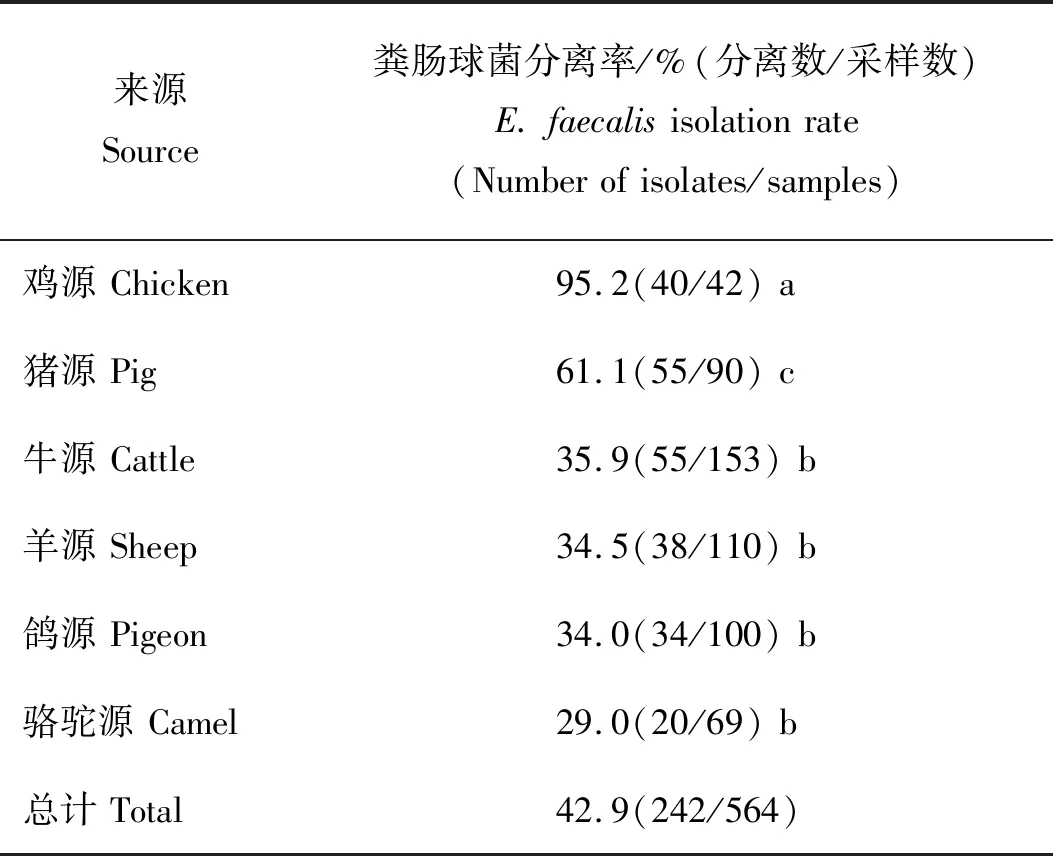
表2 动物源粪肠球菌的分离率Table 2 Isolation rate of E. faecalis from animal
2.2 粪肠球药敏试验分析
分离的242株粪肠球菌对红霉素、四环素、多西环素和利福平的耐药率最高,均大于80.0%,对万古霉素和氨苄西林高度敏感,无耐药菌株检出。此外,检出10株利奈唑胺耐药菌。
表3为6种动物源粪肠球菌对11种抗菌药物的耐药率。鸽源粪肠球菌、鸡源粪肠球菌、牛源粪肠球菌、骆驼源粪肠球对恩诺沙星耐药率在20.0%~41.2%,显著高于猪源粪肠球菌(5.5%)与羊源粪肠球菌(0)(P<0.05);鸽源粪肠球菌、猪源粪肠球菌与牛源粪肠球菌对阿米卡星的耐药率均大于80.0%,显著高于羊源粪肠球菌(7.9%)、鸡源粪肠球菌(27.5%)与骆驼源粪肠球菌(25.0%)(P<0.05);牛源粪肠球菌对庆大霉素的耐药率为96.4%,显著高于其他源粪肠球菌(P<0.05),其中羊源粪肠球菌的耐药率为7.9%,显著低于其他源粪肠球菌(P<0.05);猪源粪肠球菌、牛源粪肠球菌、鸡源粪肠球菌、骆驼源粪肠球菌对红霉素的耐药率均大于90.0%,显著高于羊源粪肠球菌(42.1%)与鸽源粪肠球菌(52.9%);四环素的耐药严重程度从高到低依次为猪源粪肠球菌(100.0%)、牛源粪肠球菌(96.4%)、鸡源粪肠球菌(92.5%)、骆驼源粪肠球菌(80.0%)、鸽源粪肠球菌(67.6%)、羊源粪肠球菌(34.2%);猪源粪肠球菌、牛源粪肠球菌、鸡源粪肠球菌、骆驼源粪肠球菌对多西环素的耐药率均大于80.0%,显著高于羊源粪肠球菌(47.4%)与鸽源粪肠球菌(55.9%)(P<0.05);羊源粪肠球菌(0)与鸡源粪肠球菌(10.0%)对氟苯尼考的耐药率显著低于其他源粪肠球菌(P<0.05)。利福平的耐药严重程度从高到低依次为鸽源粪肠球菌(100.0%)、牛源粪肠球菌(92.7%)、猪源粪肠球菌(87.3%)、骆驼源粪肠球菌(85.0%)、鸡源粪肠球菌(72.5%)、羊源粪肠球菌(52.6%)。通过上述分析不同动物源粪肠球菌对抗菌药物的耐药严重程度从高到低依次为牛源、猪源、鸽源、鸡源、骆驼源和羊源。
2.3 粪肠球菌多药耐药性分析
242株动物源粪肠球菌菌株多药耐药在0~9耐有分布,3耐及3耐以上占84.7%(205/242),以5耐和6耐为主,占53.7%(130/242)。猪源与骆驼源粪肠球菌多药耐药以5耐为主,分别占58.2%(32/55)和45.0%(9/20);鸡源粪肠球菌以5耐和6耐为主,占70.0%(28/40);牛源粪肠球菌以6耐为主,占56.4%(31/55);羊源粪肠球菌多药耐药以0~4耐为主,占97.4%(37/38);鸽源粪肠球菌以2耐和7耐为主,占44.1%(15/34)(表4)。
猪源粪肠球菌共有14种耐药谱型,在3~9耐有分布,以ERY-TET-DOX-RFP-AMK为主,占比为43.6%(24/55)。羊源粪肠球菌共12种耐药谱型,在1~5耐有分布,以ERY-TET-DOX-RFP主,占比为18.4%(7/38)。骆驼源粪肠球菌共10种耐药谱型,在1耐与3~7耐有分布,以TET-GEN-ERY-DOX-RFP为主,占比为40.0%(8/20)。牛源粪肠球菌共15种耐药谱型,在2耐与4~9耐有分布,以AMK-TET-GEN-DOX-RFP-ERY为主,占比为43.6%(24/55)。鸡源粪肠球菌共16种耐药谱型,在1~8耐有分布,以TET-DOX-GEN-ERY-RFP为主,占比为20.0%(8/40)。鸽源粪肠球菌共16种耐药谱型,在2~9耐有分布,以AMK-RFP与AMK-TET-RFP-ERY-DOX-GEN-ENR为主,各占14.7%(5/34)。
2.4 粪肠球菌耐药基因分析
242株动物源粪肠球菌除cfr和poxtA基因未检出外,其他8种耐药基因均有检出,tetM基因(78.1%)检出率最高,aph(3′)-Ⅲ、aac(6′)/aph(2″)和ermB基因的检出率也高达70.0%以上。
由表5可知不同动物源粪肠球菌中耐药基因检出结果,猪源粪肠球菌耐药基因携带率最高,ermB(100.0%)基因携带率显著高于其他动物源粪肠球菌(P<0.05)。猪源粪肠球菌检出8种耐药基因,ermA基因(5.5%)仅在猪源粪肠球菌中检出。猪源、鸡源、牛源和骆驼源粪肠球菌tetM基因的携带率均大于80.0%,显著高于鸽源(55.9%)与羊源(26.3%)粪肠球菌(P<0.05)。aac(6′)/aph(2″)基因的携带率从高到低依次为牛源(100.0%)、猪源(96.3%)、骆驼源(80.0%)、鸡源(75.0%)、鸽源(38.2%)、羊源(28.9%)。羊源、鸽源粪肠球菌对aph(3′)-Ⅲ基因的携带率显著低于其他源粪肠球菌(P<0.05)。
3 讨 论
粪肠球菌作为肠球菌属中临床分离率最高的菌种[20],耐药机制复杂,易产生耐药性,成为耐药基因的贮存库,获得性交叉耐药等问题给兽医及人医临床带来了巨大挑战[21]。本研究以新疆9个县区分离的6种动物源粪肠球菌为研究对象,分析不同动物源粪肠球菌的耐药特点及耐药基因携带情况,对于临床用药具有指导意义。
本研究中新疆9个县区粪肠球菌的整体分离率为42.9%,略高于魏莲花等[22]报道的中国西部地区人医临床粪肠球菌与李延山等[23]报道的东北地区不同动物源粪肠球菌的分离率。不同动物源以鸡源粪肠球菌分离率最高,达95.2%,高于李金磊等[24]报道的河南省部分地区鸡源粪肠球菌分离率(55.6%)。本研究中猪源粪肠分离率为61.1%,与王舒丰等[7]报道的阿克苏地区猪源粪肠球分离率基本一致。牛源、鸽源、骆驼源和羊源粪肠球菌的分离率差别不大,都在30.0%左右。结果显示,粪肠球菌作为肠道共生菌在不同动物中具有较高分离率,但在不同动物体内的比例具有一定差异性。
药敏试验结果表明,不同动物源粪肠球菌对四环素、多西环素和红霉素的高耐药率与姚晓慧等[25]报道的粪肠球菌的耐药结果基本一致,分析高耐药率产生的原因,可能是由于四环素类与大环内酯类的一些抗菌药物属于农业部批准的饲料药物添加剂,从而导致菌株对其耐药严重;此外,利福平、庆大霉素和阿米卡星,这3种药物的耐药率也偏高,其原因可能是这几种药物的性价比高,长期不规范使用导致耐药菌株的大量出现。猪源粪肠球菌对恩诺沙星的耐药率(5.5%)相较于其他几种常用药物耐药率较低。新疆不同动物源粪肠球菌对氟苯尼考(16.1%)与氨苄西林(0)的耐药率总体偏低,提示该药物可作为治疗不同动物细菌性疾病的备选药物。未检出耐万古霉素肠球菌(Vancomycin-resistantEnterococcus,VRE)。值得注意的是,检出10株利奈唑胺耐药菌株,其原因可能是与酰胺醇类药物交叉耐药所致,例如氟苯尼考。虽然不同动物源粪肠球菌对氟苯尼考耐药率较低,但也应严格控制该药物的使用,通过联合用药或轮换用药减少氟苯尼考耐药菌株的出现,从而避免利奈唑胺耐药菌株的爆发。
不同动物源粪肠球菌菌株多药耐药集中在5耐和6耐,耐药谱型以ERY-TET-DOX-RFP-AMK-GEN为主。不同动物源粪肠球菌的耐药谱型差别较大,其原因可能是,菌株来源于不同地区不同动物,用药习惯存在差异。牛源、猪源和骆驼源粪肠球菌耐药率高,耐药情况最为严重,其原因可能是因为这3种动物饲养周期长,使用抗菌药物几率更高,导致分离菌株耐药严重。鸽源与鸡源粪肠球菌耐药谱最广,耐药率较高,其原因可能是养殖户缺乏规范用药意识,不合理使用甚至滥用药物,导致多耐菌株和高耐菌株的出现。羊源粪肠球菌对被检药物的耐药率整体偏低,对多种药物敏感性良好,其原因可能是养殖场用药合理,采样的羊群为一月龄胡羊,多数未使用抗菌药物。不同养殖场可根据药敏试验结果与耐药谱型,在目前的用药模式上合理选用具有较高敏感性的抗菌药物,在提高治疗效果降低经济成本的同时,也可缓解交叉耐药情况的产生。
本研究耐药基因检测结果显示,新疆动物源粪肠球菌耐药基因携带率高,对被检测的5大类耐药基因均有检出。猪源粪肠球菌对被检耐药基因的携带率最高,检出的种类也最多,其次为牛源粪肠球菌,其原因可能是样品采集的猪场和牛场饲养动物密集,携带多种耐药基因的粪肠球菌易通过粪便、污水等进行传播。羊源耐药基因携带率最低与药敏试验结果相符。tetM(78.1%)、ermB(71.5%)基因的携带率与Molechan等[26]报道的南非鸡源肠球菌一致,但aac(6′)/aph(2″)(73.6%)、aph(3′)-Ⅲ(74.0%)基因的携带率却显著高于其报道,略低于国内吴安琪等[27]的报道。其原因可能是对于氨基糖苷类药物国内用药习惯与国外存在差异,相关部门应加大监察力度,规范该类药物合理使用。氟苯尼考的耐药率为16.1%,与酰胺醇类耐药基因fexA检出率(16.9%)基本一致。虽然噁唑烷酮类多药耐药基因cfr与poxtA基因未检出,但是optrA基因检出率高达9.1%,除羊源未检出外,其他动物源均有检出。少数携带该基因的菌株对利奈唑胺没有表现出耐药,但为中介值,结果印证Osei等[28]报道的药敏试验中介的菌株也可能扩增出相应耐药基因的结论。optrA基因不仅介导对酰胺醇类抗菌药物和利奈唑胺的耐药,还可介导对2014年被美国FDA批准上市的新一代噁唑烷酮类药物—泰地唑利的耐药,该药被批准用于临床治疗耐甲氧西林的金黄色葡萄球菌(Methicillin-resistantStaphylococcusaureus,MRSA)和耐甲氧西林的凝固酶阴性的葡萄球菌(Methicillin-resistant coagulase negativestaphylococci,MRCoNS)等多重耐药菌引起的感染[14]。因此,应该注意对该基因的持续监测,避免交叉耐药危害人类健康。
综上所述,粪肠球菌在新疆9个县区的6种动物粪样均中具有较高的检出率,且分离到的粪肠球菌对四环素、红霉素、多西环素和利福平的耐药率高,养殖过程中应减少这4种药物的使用,可根据药敏结果选用恩诺沙星、氟苯尼考和氨苄西林等敏感性高的药物来提高疗效。猪、牛和骆驼,饲养周期长的动物,耐药严重,耐药基因携带率高,应严格控制抗菌药物的使用,合理轮换用药。鸡、鸽在饲养过程中,应注意用药的规范化,避免滥用药物。在除羊外的5种动物中,均检出噁唑烷酮类耐药基因optrA,提示应该注意酰胺醇类药物的使用与相关耐药基因的持续监测,实验室与临床结合,避免交叉耐药的产生,保护公共卫生安全。
猜你喜欢
淡水渔业(2022年4期)2022-07-29
中国典型病例大全(2022年7期)2022-04-22
中国典型病例大全(2022年7期)2022-04-22
保健与生活(2019年2期)2019-08-01
特别健康·下半月(2019年7期)2019-07-29
山西农经(2018年1期)2018-05-14
植物保护(2018年1期)2018-05-14
大自然探索(2017年8期)2017-08-28
特别健康·下半月(2017年5期)2017-06-15
医学信息(2016年7期)2016-10-14
