
无瓣海桑叶斑病及其病原菌的生物学特性研究*
2022-02-02 09:06冯金艳陆建康张良军刘幸儿田龙艳
林业与环境科学 2022年5期
黄 华 冯金艳 陆建康 张良军刘幸儿 田龙艳
(1. 肇庆市封开县林业局森林病虫害防治检疫站,广东 肇庆 526500;2.广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广东 广州 510520)
拟盘多毛孢属Pestalotiopsis由Steyaert 命名,他将广义盘多毛孢属中分生孢子具5 个细胞、中间具有3 个深色细胞的种归入Pestalotiopsis属[1]。按照最新的分类系统拟盘多毛孢属属于子囊菌门,盘菌亚门,粪壳菌纲,炭角菌亚纲,黑盘孢目,被毛孢科[2]。这类真菌在热带和亚热带地区分布最为广泛,寄主专一性不高,适应能力强,可引起多种植物病害,是一类重要的植物病原菌[3-4]。
无瓣海桑Sonneratia apetala属于桃金娘目海桑科海桑属的红树树种[5]。天然分布于孟加拉国、印度等国家。由于无瓣海桑具有生长快、适应性强、能改善滩涂立地条件、加快滩涂绿化等特点,1985 年被首次引种到我国海南岛,在20 世纪90年代又从海南引种到广东、广西、福建等地[6]。目前已成为我国华南沿海生态保护与恢复中最重要的红树树种[7-8]。
无瓣海桑在我国虫害危害较为严重[9-10],病害危害较轻,有关无瓣海桑病害的研究尚鲜有报道[11]。前期研究发现无瓣海桑病叶组织分离到菌株WBHS-YB-2A,为拟盘多毛孢属新种Pestalotiopsis sonneratiae(未发表数据)。为了进一步确定无瓣海桑叶斑病的病原菌,本研究采用柯赫氏法则测定WBHS-YB-2A 的致病性、进行形态学观察和生物学特性等研究,以期明确无瓣海桑叶斑病及其病原菌的生物学特性,为无瓣海桑叶斑病的识别及防治提供研究基础。
1 材料与方法
1.1 试验材料
Pestalotiopsis sonneratiae: 菌 株WBHS-YB-2A,保藏于中国林业科学研究院,保藏号CFCC 57390。PDA 培养基:去皮土豆200 g·L-1,葡萄糖20 g·L-1,琼脂粉15 g·L-1,pH 调至7.0。察氏培养基:KH2PO42.0 g·L-1,MgSO41.0 g·L-1,KNO32.0 g·L-1,葡萄糖20.0 g·L-1,琼脂20.0 g·L-1,pH 调至7.0。
1.2 试验方法
在PDA 平板上活化菌株WBHS-YB-2A,转接后培养5 d,备用。取新鲜健康的叶片清洗2次,用70%的酒精进行表面消毒,无菌水清洗2次,晾干后置于铺有润湿滤纸的培养皿中,备用。在菌株WBHS-YB-2A 菌落边缘用打孔器取直径5 mm 菌饼,接种于叶片上,以空白PDA 培养基作对照,设置8 个重复。放置于光照培养箱中培养(25 ℃,12 h 光照培养;22 ℃,12 h 暗培养),每日观察发病情况,记录病斑大小与发病率。采用离体接种方法进行致性测定[12-13]。
1.3 形态学特征
将活化的菌株WBHS-YB-2A 接种于直径9 cm的PDA 平板上,(25±1)℃恒温培养10 d。每隔2 d 观察菌落培养形态,待产孢后,挑取分生孢子于显微镜下观察形态特征。
1.4 生物学特性分析
1.4.1 碳源试验 以察氏培养基为基础培养基,其中碳源葡萄糖以可溶性淀粉、玉米粉、麦芽糖、蔗糖、果糖等量替换,配制成含糖量相同碳源不同的培养基[14-15],灭菌后倒平板,接入5 mm 菌饼,以基础培养基为对照,每个处理3 个重复。置于(25±1)℃恒温培养箱中,每2 d 观察产孢、菌丝致密度等情况,同时测量菌落直径,记录生长速率。
1.4.2 氮源试验 以察氏培养基为基础培养基,其中的氮源KNO3以酵母浸粉、谷氨酸、尿素、蛋白胨等量替换,配制成含氮量相同氮源不同的培养基。接种、培养与数据统计与碳源试验处理一致。
1.4.3 温度试验 以PDA 培养基为基础培养基,将5 mm 菌饼接种到PDA 平板培养基中央,分别放入10、15、20、25、30 ℃的培养箱中培养,接种、培养与数据统计与碳源试验处理一致。
1.4.4 pH 试验 以PDA 培养基为基础培养基,用0.1 mol·L-1NaOH 和0.1 mol·L-1HCl,将培养基pH 分别调为4.0、5.0、6.0、7.0、8.0。按碳源试验的方法进行培养基的配制、接种、观察与测量。
1.5 数据分析
利用软件Microsoft Office Excel 2010 和SPSS 25.0 进行Duncan 氏新复极差差异显著性分析。
2 结果与分析
2.1 致病性
由图1 可知,接种3 d 后,无瓣海桑新鲜健康的叶片出现褐色点状病斑;6 d 后,表现为黑褐色圆形病斑;10 d 后病斑中心产生黑色的分生孢子堆,病斑可扩散至整个叶片。对照和老叶均未发病,接种感病症状与田间症状相似。对接种后发病的叶片进行分离,获得的菌株菌落形态、孢子形态、以及ITS 序列与菌株WBHS-YB-2A 一致。结果表明,菌株WBHS-YB-2A 对无瓣海桑有致病性,确定P. sonneratiae为无瓣海桑叶斑病的病原菌。

图1 室内离体接种症状Fig.1 Symptom of inoculation in vitro
2.2 无瓣海桑叶斑病危害症状
由P. sonneratiae引起的无瓣海桑叶斑病,在老叶和幼嫩的叶片均可发生,发病初期表现为褐色小点,病斑边缘有黄色的晕圈;随着病菌扩展,病斑逐渐扩大呈圆形或椭圆形或不规则,病斑外缘有黄色的晕圈,病斑外圈呈黑褐色,中心坏死褪色呈浅咖色,叶片不脱落。
2.3 无瓣海桑叶斑病菌形态特征
在PDA 上,无瓣海桑叶斑病菌菌落平展近圆形,产生白色絮状气生菌丝,菌丝发达致密,同时产生大量的分生孢子堆,呈墨色浓稠粘液,菌落同时伴有产生浅黄色至褐色沉淀(图2 A 和C)。在察氏培养基上,菌落不规则,菌落形态类似花瓣,菌丝较致密,菌落伴有产生浅棕色沉淀,未形成分生孢子堆(图2B)。分生孢子梗透明,光滑,柱状至壶状,环生。分生孢子梗不明显,常退化(图2D)。分生孢子纺锤形,直或稍弯曲,有4 隔膜,中间细胞3 个,棕色;顶端中间处附属物单2~3(图2E)。
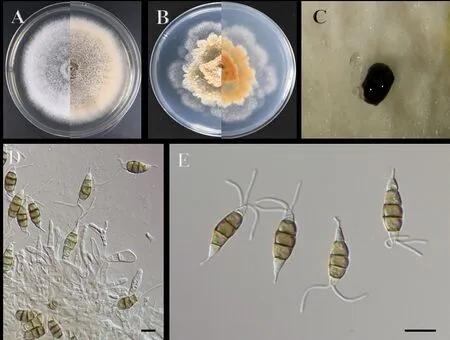
图2 菌株培养形态Fig.2 Morphology of the cultured strain
2.4 无瓣海桑叶斑病菌的生物特性
2.4.1 不同碳源对无瓣海桑叶斑病菌的影响 由表1 可知,在不同碳源条件下,无瓣海桑叶斑病菌均可生长。在不同碳源培养基上菌丝生长速率依次为:葡萄糖>麦芽糖=蔗糖>淀粉>玉米粉>果糖,前四者和后两者之间差异显著,不同碳源培养基上均产生黄色色素,菌丝均较致密,无明显差异;在缺碳培养基上菌丝稀疏,生长速度最慢。由此可知该病原菌的最佳碳源为葡萄糖。
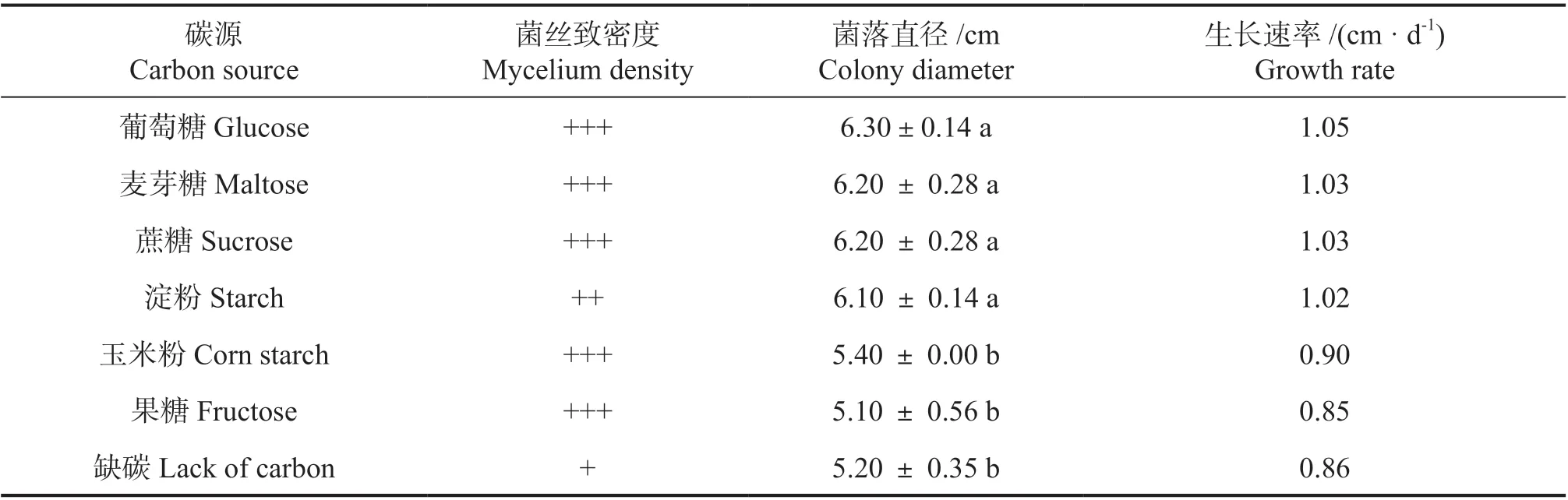
表1 不同碳源对菌株WBHS-YB-2A 的影响Table 1 Effects of different carbon sources on strain WBHS-YB-2A
2.4.2 不同氮源对无瓣海桑叶斑病菌的影响 由表2 可知,不同氮源培养基上菌株WBHS-YB-2A均可生长,但菌落形态、菌丝致密度差异较大,生长速率显著不同。其生长速率依次为:蛋白胨>酵母膏>谷氨酸>KNO3>尿素;以蛋白胨为氮源时,菌丝较致密,生长速度最快可达1.21 cm·d-1;以尿素为氮源时,菌丝致密,但生长最缓慢,生长速度仅为0.75 cm·d-1。
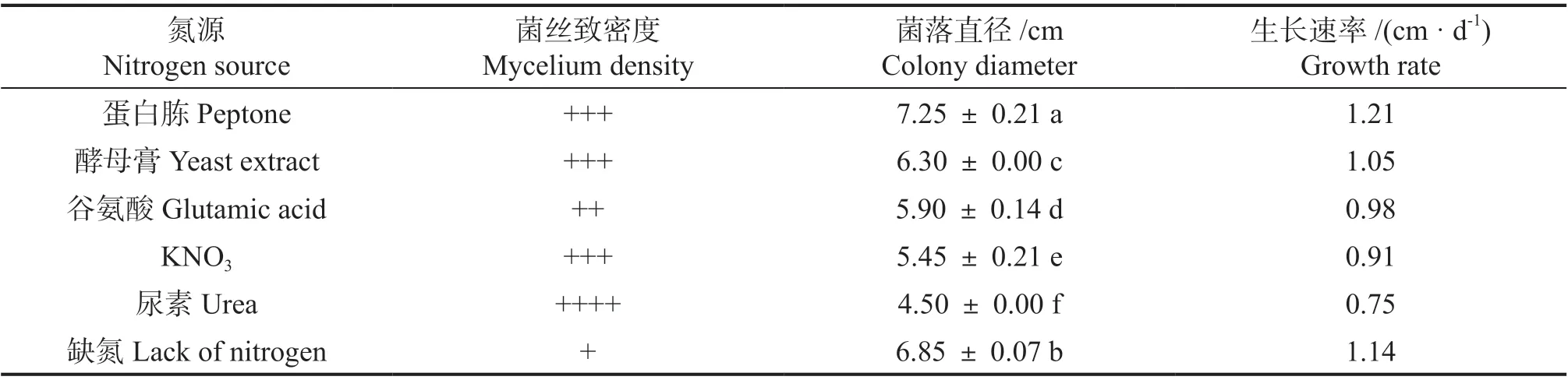
表2 不同氮源对菌株WBHS-YB-2A 的影响Table 2 Effects of different nitrogen sources on strain WBHS-YB-2A
2.4.3 不同温度对无瓣海桑叶斑病菌的影响 由表3 可知,不同温度对菌株菌株WBHS-YB-2A生长影响极显著(P<0.01)。菌丝生长速率依次为:25 ℃>20 ℃>30 ℃>15 ℃>10 ℃。当温度≥30 ℃或≤15 ℃时,菌丝生长速率显著下降,其最适宜生长温度为20~30 ℃。

表3 不同温度对菌株WBHS-YB-2A 的影响Table 3 Effect of different temperature on strain WBHS-YB-2A
2.4.4 不同pH 对无瓣海桑叶斑病菌的影响 由表4 可知,菌株WBHS-YB-2A 在不同pH 培养基上生长速率依次为6.0>5.0>7.0=4.0>8.0。该菌偏好于酸性的生长环境。
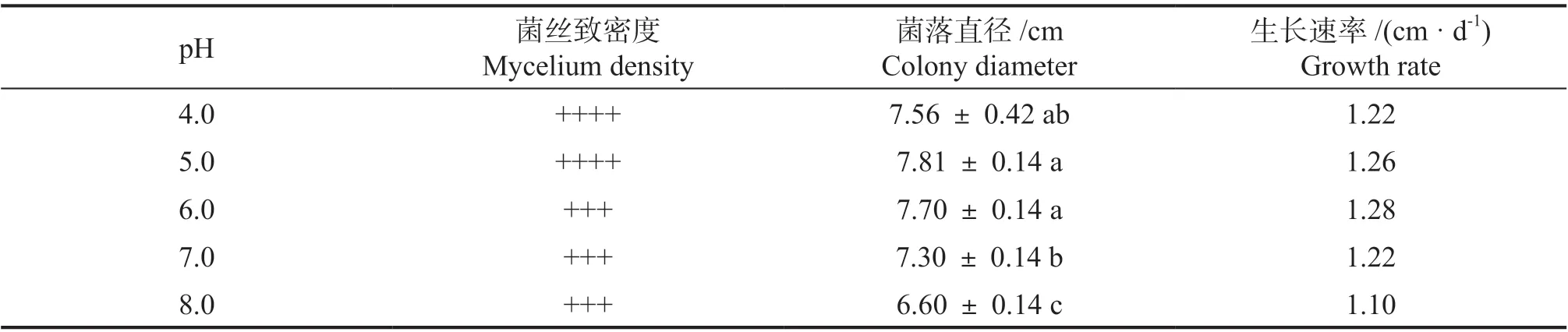
表4 不同pH 对供试菌株WBHS-YB-2A 的影响Table 4 Effect of different pH on strain WBHS-YB-2A
3 讨论与结论
拟盘多毛孢属的很多成员是重要的植物病原菌,能够造成农林作物减产和严重的经济损失。有报道表明,该类病原菌可在草莓、葡萄、苹果、红毛丹树、茶、莲雾等多种植物上引起病害.其中,叶斑病是其危害中最为常见的病害类型[16]。一般情况下,拟盘多毛孢的寄主专化性较弱,很多病原菌具有广泛的寄主[17]。在前期研究中,我们从无瓣海桑叶片和枝干病组织分离到的4 株拟盘多毛孢属菌株,形态学观察无明显差异,基于多片段构建的系统发育树分析,确定菌株WBHSYB-2A 为拟盘多毛孢属新种P. sonneratiae(未发表数据)。本文系统研究了P. sonneratiae菌株的致病性与生物学特性,发现叶片中分离得到的菌株WBHS-YB-2A 对新鲜幼嫩叶片具有较强的致病性,确定为无瓣海桑叶斑病的病原菌。同时,从无瓣海桑枝干上分离的P. sonneratiae菌株对叶片也具有致病性。但是,P. sonneratiae对无瓣海桑枝干是否具有致病性仍待进一步研究。
本研究针对无瓣海桑叶斑病菌的生物学特性进行了分析,明确了该菌对不同的碳源、氮源的利用情况,确定该菌最适碳源为葡萄糖、最适氮源为蛋白胨,在10~30 ℃均能生长,较适宜偏酸性环境条件,说明无瓣海桑叶斑病菌具有较强的适应能力,这与引起莲雾软腐病的异色拟盘多毛孢P.versicolor类似[18]。目前,国内外有关无瓣海桑叶斑病以及病原菌的报道较少,有关该类病害的致病机制、发生流行规律及其防治技术等尚不清楚,亟待开展相关研究。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
甘肃教育(2021年10期)2021-11-02
昆钢科技(2021年6期)2021-03-09
天津科技(2020年4期)2020-05-09
现代农业科技(2016年23期)2017-04-06
中国酿造(2016年12期)2016-03-01
中国酿造(2014年9期)2014-03-11
植物营养与肥料学报(2011年6期)2011-10-24
