
黑曲霉发酵脱脂麦胚过程中抗氧化活性变化研究
2022-02-07 13:09冯军伟刘世欣段兰兰侯银臣董必辉董得平王晓飞隋志方
中国酿造 2022年1期
冯军伟,刘世欣,段兰兰,侯银臣,董必辉,董得平,王晓飞,隋志方
(1.河南飞天农业开发股份有限公司,河南 鹤壁 456750;2.河南牧业经济学院 食品与生物工程学院,河南 郑州 450046;3.鹤壁市质量技术监督检验测试中心,河南 鹤壁 458000)
小麦是世界三大粮食作物之一,中国是世界上最大的小麦生产国[1-2]。麦胚占整个小麦籽粒的2%~3%,是小麦籽粒营养价值最高的部分,年蕴藏量约420~480万t,而利用率却不足2%,开发潜力巨大由于麦胚本身不易储藏和极易氧化的特性等原因,目前对小麦胚芽的开发利用还不够充分,利用率并不高,造成大量浪费[3-4]。麦胚营养丰富,被营养学家称为“人类天然的营养宝库”[5],富含蛋白质[6]、脂肪[7]、糖类物质[8]、纤维素、半纤维素、多种水溶性维生素、矿物质及一些微量的生理活性物质[9]。麦胚不仅营养丰富,还具有多种生理活性功能,如抗过敏[10]、抗氧化[11]、抗肿瘤[12]、降血压[13]、免疫调节[14]、抗炎[15]等。黑曲霉(Aspergillus niger)能产生丰富的酶系,如淀粉酶、纤维素酶、蛋白酶,是重要的发酵工业菌种[16],微生物发酵制备生物活性肽是目前研究的一个热点。赵朋辉等[17]以抗氧化活性为评价指标,优化了黑曲霉发酵麦胚工艺,并对醇沉组分的理化性质进行了研究;牛丽亚等[18]对黑曲霉发酵麦胚提取黄酮的工艺条件进行了研究;另外也有利用枯草芽孢杆菌、酵母等微生物对麦胚发酵的相关研究[19]。
本研究以脱脂麦胚为原料,利用黑曲霉(Aspergillus niger)对其发酵,采用动态活性追踪等方法研究了发酵过程中发酵产物体外抗氧化活性、蛋白酶活及发酵液中蛋白含量的动态变化。发酵液中蛋白酶活及蛋白含量的动态变化解释了发酵液抗氧化变化的原因,旨在为黑曲霉可控制发酵麦胚生产抗氧化活性肽提供了参考。
1 材料与方法
1.1 材料与试剂
脱脂麦胚:河南省鲲华生物技术有限公司;黑曲霉(Aspergillus niger):来源于河南工业大学菌株保藏中心;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基:美国Sigma公司;牛血清蛋白(bovine serum albumin,BSA):北京索莱宝科技有限公司;氧化铁、硫酸亚铁、考马斯亮蓝、水杨酸、福林酚、30%双氧水、酪氨酸(均为分析纯):洛阳昊华化学试剂有限公司;氯乙酸、天青I、邻苯三酚、氯化钠、三羟甲基氨基甲烷、铁氰化钾(均为分析纯):合肥千盛生物科技有限公司。
马铃薯葡萄糖琼脂(potato dextrose agar,PDA)培养基:200 g马铃薯切块,煮沸后取过滤液,添加20 g葡萄糖,15~20 g琼脂,添加蒸馏水至1 L,分装后,121 ℃灭菌20 min。PDA液体培养基不添加琼脂。
1.2 仪器与设备
LDZX-50KBS立式压力蒸汽灭菌锅:上海申安医疗器械厂;H1850R医用冷冻离心机:湘仪离心机仪器有限公司;SW-CJ-1S无菌工作台:苏州净化设备有限公司;FA224电子分析天平:上海舜宇恒平科学仪器有限公司;BlueStar B紫外可见分光光度计:北京莱伯泰科仪器股份有限公司;DHP-080电热恒温培养箱:郑州生元仪器有限公司;TS-2112B摇床:无锡玛瑞特科技有限公司。
1.3 实验方法
1.3.1 黑曲霉种子液制备
将保存的黑曲霉在PDA琼脂培养基上活化,然后挑取活化好的菌丝接种到装液量为150 mL的PDA液体培养基中进行扩培并制备发酵种子液,活化和扩培条件均为培养温度28 ℃、120 r/min,培养时间36 h。
1.3.2 黑曲霉发酵脱脂麦胚
脱脂麦胚按照料液比1∶9(g∶mL)的比例加入蒸馏水,121 ℃灭菌20 min。按照接种量10%接入黑曲霉种子液,28 ℃条件下,置于120 r/min的摇床上发酵。取出不同发酵时间点的发酵液,5 000 r/min离心10 min,取上清测定其体外抗氧化活性、蛋白含量、蛋白酶活等指标。
1.3.3 脱脂麦胚黑曲霉发酵液体外抗氧化活性
(1)DPPH·清除能力
参考HOU Y C等[20]的方法,略作修改。取发酵上清液0.2 mL,然后加入2.8 mL体积分数95%乙醇补足至3 mL,混匀后再加入3 mL DPPH溶液,室温避光反应30 min后,于波长517 nm处测定吸光度值(Ai)。空白对照为发酵上清液0.2 mL,加入5.8 mL体积分数95%乙醇补足6 mL,室温避光反应30 min,测定吸光度值(Aj)。空白组为3 mL甲醇溶液和3 mL DPPH溶液摇匀混合,室温避光反应30 min,测定吸光度值(A0),按下式计算发酵上清液对DPPH·的清除率:
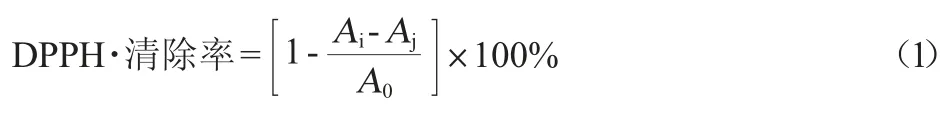
(2)O2-·清除能力
参照郝董林[21]超氧阴离子自由基清除率的测定方法,略作修改。取两支25 mL比色管,分别加入5.0 mL0.2 mmol/L天青I溶液和1.4 mL pH 9.0的Tris-HCl缓冲溶液后,向其中一支加入0.80 mL 30 mmol/L的邻苯三酚溶液,另一支不加。分别用水稀释至刻度,在室温条件下反应15 min,以水为参比,用1 cm比色皿在波长602 nm处测定体系的吸光度。设不加邻苯三酚时体系的吸光度值为A0,加入邻苯三酚后体系的吸光度值为A1,按上述步骤操作,在加入5.00 mL天青I溶液和pH 9.0 的缓冲溶液后,向其中一支比色管中加入样品,再分别加入0.80 mL邻苯三酚溶液,用水稀释至刻度定容后,摇匀。室温条件下反应9 min后,分别测定吸光度值,加样品的溶液的吸光度值记为A2。
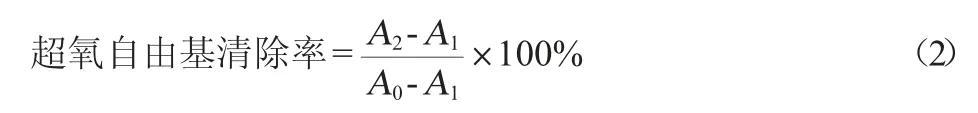
(3)·OH的清除能力测定
参照刘皓涵等[22]的方法,略作修改。取1.0 mL待测液与1.0 mL 9 mmol/L的FeSO4、1.0 mL 9 mmol/L的水杨酸-乙醇和1.0 mL 8.8 mmol/L的H2O2混匀后37 ℃静置反应10 min,在波长510 nm处测其吸光度值(A1),空白溶液不添加H2O2溶液(A0),参比溶液不添加H2O2(Ax0)。羟自由基清除能力计算公式如下:
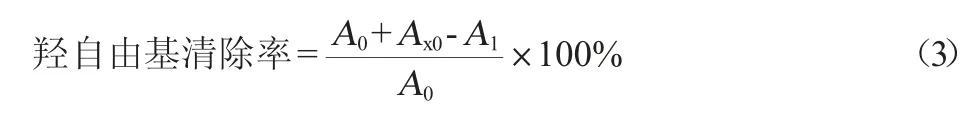
(4)总还原力的测定
分别精密吸取不同发酵时间的麦胚提取液0.1 mL置于试管中,然后加入0.9 mL体积分数95%乙醇补足至1 mL,同时精密吸取1.0 mL体积分数95%乙醇作为空白对照,然后依次加入2.5 mL磷酸缓冲液,再加入2.5 mL 1%铁氰化钾溶液,混合摇匀,封口后,置于水浴锅中50 ℃恒温水浴20 min。水浴结束后迅速冷却,再加入2.5 mL10%三氯乙酸溶液,混合摇匀,4 000 r/min离心10 min后精密量取2.5 mL上清液,依次加入2.5 mL蒸馏水、1.0 mL 0.1%三氯化铁溶液,混合摇匀,静置10 min后在波长700 nm处测定吸光度值(A700nm值),以A700nm值大小直接衡量其还原力大小[23],A700nm值越大,还原能力越大,表示其抗氧化活性越强。
1.3.4 蛋白含量的测定
采用Folin-酚显色法测定蛋白质含量[24]。将100 μg/mL标准牛血清白蛋白溶液以梯度浓度加入,依次加入0.4 mol/L碳酸钠溶液,Folin-酚试剂,迅速混合均匀,将其放置于40 ℃的水浴锅中加热20 min,取出后迅速冷却,采用紫外分光光度计进行比色,在波长650 nm处测得吸光度值A650nm。以牛血清白蛋白含量(x)为横坐标,吸光度值(y)为纵坐标,绘制牛血清白蛋白标准曲线,得到标准曲线回归方程为:y=0.030 9x+0.032 2,相关系数R2=0.957 5。
1.3.5 蛋白酶活力的测定
参考刘珊[25]的方法,略有改进。将100 μg/mL酪氨酸标准溶液配制成不同浓度梯度(10 μg/mL、20 μg/mL、30 μg/mL、40 μg/mL、50 μg/mL),各取1.00 mL,分别加入0.4 mol/L碳酸钠溶液5.00 mL,福林试剂使用溶液1.00 mL,置于40 ℃水浴中反应20 min,以不含酪氨酸的为空白管调零,在波长680 nm处分别测定其吸光度值,以酪氨酸质量浓度(x)为横坐标,吸光度值(y)为纵坐标,绘制酪氨酸标准曲线,得到标准曲线回归方程为:y=0.010 1x-0.003 7,相关系数R2=0.995 6。
蛋白酶活力的测定:先将2%酪蛋白溶液1 mL放入40 ℃水浴5 min后各加入1 mL样液,取一支作为空白管,加2 mL三氯乙酸,其他3管作为测试管各加入1 mL酪蛋白,摇匀,40 ℃保温10 min,取出试管,3支测试管中各加入2 mL三氯乙酸,空白管中加1 mL酪蛋白,静置10 min,过滤各取1 mL滤液,加入0.4 mol/L的碳酸钠5 mL、福林试剂1 mL,在40 ℃显色20 min,以空白管调零,在波长680 nm处测定OD680nm值。在40 ℃下每分钟水解酪蛋白产生1 μg酪氨酸,定义为1个蛋白酶活力单位(U/mL)。蛋白酶活计算公式如下:

式中:A为样品平行试验的平均OD680nm值;K为吸光常数;4为反应试剂的总体积,mL;10为酶解反应时间,min;n为酶液稀释总倍数。
2 结果与分析
2.1 发酵液体外抗氧化活性分析
2.1.1 发酵液清除DPPH·能力
由图1可知,脱脂麦胚本身具有一定的DPPH·清除能力,这主要是因为麦胚本身含有多酚、黄酮及一些肽类[26]。随着发酵时间的延长,0~36 h内无显著变化(P>0.05),此时主要是黑曲霉生长阶段,发酵36 h后,随发酵时间的延长其对DPPH·的清除能力呈先增强后减弱的趋势,当发酵时间达到84 h时,其清除DPPH·的能力最强,为75.68%,随后减弱,这可能是因为发酵过程中黑曲霉分泌的蛋白酶降解了脱脂麦胚中的蛋白,生成了小分子肽等抗氧化活性成分,从而造成发酵液清除DPPH·能力逐渐增加,随后可能是由于抗氧化活性成分的过度水解或者被氧化造成其对DPPH·的清除能力逐渐减弱。

图1 发酵过程中发酵液清除DPPH·能力变化Fig.1 Changes of DPPH·scavenging ability of fermentation broth during fermentation process
2.1.2 发酵液清除·OH能力
由图2可知,随着脱脂麦胚发酵时间在0~120 h的延长,发酵液对·OH的清除能力呈先增强后减弱的趋势,发酵36 h后,增加幅度明显增强。当发酵时间达到84 h时,发酵液的·OH清除能力达到峰值,为80.40%,随后呈现缓慢降低的趋势。
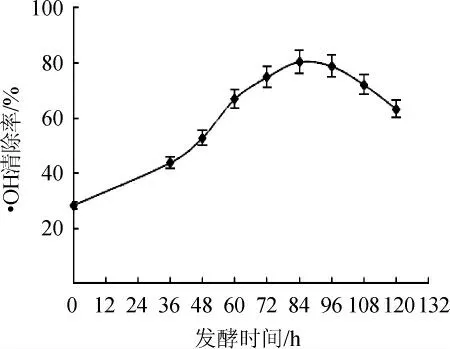
图2 发酵过程中发酵液清除·OH能力变化Fig.2 Changes of·OH scavenging ability of fermentation broth during fermentation process
2.1.3 发酵液清除O2-·能力
由图3可知,随着脱脂麦胚发酵时间在0~84 h的延长,发酵上清液对O2-·的清除能力逐渐缓慢上升,当发酵时间达到60 h时后上升幅度较大,当发酵时间为84 h时,其对O2-·的清除能力达到最大值,为40.7%,之后又随着发酵时间的延长逐渐降低。
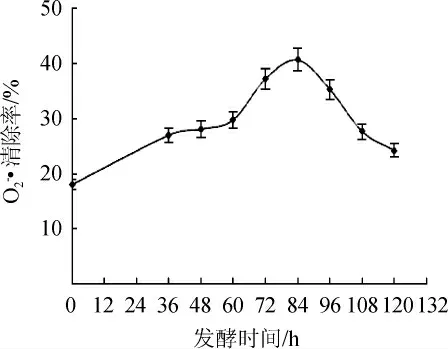
图3 发酵过程中发酵液清除O2-·能力变化Fig.3 Changes of O2-·scavenging ability of fermentation broth during fermentation process
2.1.4 发酵液总还原力
对不同发酵时间点的上清液总还原力进行测定,结果见图4。由图4可知,随着发酵时间的延长,黑曲霉发酵脱脂麦胚上清液的还原力与其他体外抗氧化活性呈现较为类似的趋势,缓慢上升后逐渐下降,当发酵时间为84 h时,发酵上清液总还原力达到最大值,吸光度值达到0.476 7。
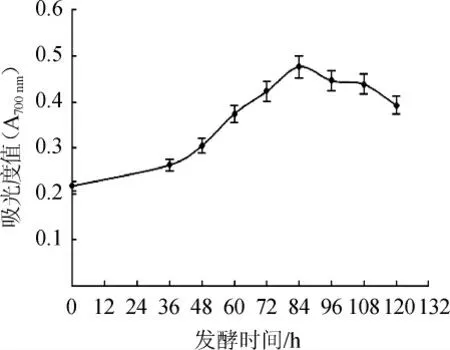
图4 发酵过程中发酵液总还原力变化Fig.4 Changes of total reducing power of fermentation broth during fermentation process
2.2 发酵上清液蛋白含量变化
检测不同时间点的脱脂麦胚发酵液蛋白含量,结果见图5。由图5可知,随着发酵时间的延长,脱脂麦胚发酵液中的蛋白含量呈现逐渐上升趋势,当发酵时间达到108 h时,蛋白含量达到最大值21.92 mg/mL,之后略有下降,但无显著变化(P>0.05)。麦胚本身有一定的水溶性蛋白和肽类,因此发酵初期其发酵液中含有一定的蛋白含量,随着发酵时间的延长黑曲霉发酵过程中会产生蛋白酶,这些蛋白酶造成非水溶性蛋白质的降解生成一些肽类、氨基酸和小分子的水溶性蛋白,因此发酵液中的蛋白质含量呈现上升趋势,这也说明发酵液的体外抗氧化活性的升高与发酵液中的蛋白质含量密切相关,这与枯草芽孢杆菌发酵花生粕制备抗氧化肽的研究内容相符合[27]。

图5 发酵过程中发酵液蛋白含量变化Fig.5 Changes of protein contents of fermentation broth during fermentation process
2.3 发酵液蛋白酶活力变化
检测不同时间点的脱脂麦胚发酵液蛋白酶活力变化,结果见图6。由图6可知,发酵时间在0~108 h范围内,黑曲霉发酵脱脂麦胚上清液蛋白酶活力变化呈现缓慢上升后保持稳定的趋势,当发酵时间为72 h时,发酵液蛋白酶活力达到最大值,为269.35 U/mL,之后保持平稳,发酵108 h后蛋白酶活呈现下降的趋势,这可能是因为酶活力损失造成。

图6 发酵过程中发酵液中蛋白酶活力变化Fig.6 Changes of protease activity of fermentation broth during fermentation process
3 结论
研究结果表明,随着脱脂麦胚发酵时间的延长,发酵液对DPPH·、·OH和O2-·的清除能力及其总还原力均呈现先增加后缓慢降低的趋势,当发酵时间达到84 h时,发酵液的体外抗氧化活性最佳,其清除DPPH·的能力为75.68%,清除·OH能力为80.40%,清除O2-·能力为40.7%,总还原力为0.4767(A700nm)。体外抗氧化活性的变化趋势与发酵液中蛋白含量的增加密不可分,随发酵时间的延长,脱脂麦胚发酵液中的蛋白含量呈现逐渐上升趋势,当发酵时间达到108 h时,发酵液蛋白含量达到最大值21.92 mg/mL,而发酵液蛋白含量的提高又是因为黑曲霉发酵脱脂麦胚过程中分泌蛋白酶所致,因此黑曲霉发酵脱脂麦胚过程中可以通过分泌的蛋白酶对麦胚中的蛋白进行降解,生成具有抗氧化活性的多肽。本研究为脱脂麦胚抗氧化产品的开发提供了一定的参考依据。
猜你喜欢
农业工程学报(2022年1期)2022-03-25
新乡医学院学报(2022年2期)2022-03-22
中国果业信息(2021年10期)2021-12-07
农技服务(2021年3期)2021-06-16
发明与创新·大科技(2021年2期)2021-04-20
中国生殖健康(2020年4期)2021-01-18
食品界(2019年8期)2019-10-07
船海工程(2017年3期)2017-07-05
江苏农业科学(2016年1期)2017-05-17
江苏农业科学(2016年1期)2017-05-17
