
不同年限毛竹-白及复合系统土壤微生物群落多样性特点
2022-03-05 06:12曹小青孙孟瑶王晶晶李培玺储炳银徐小牛
中国土壤与肥料 2022年1期
曹小青,王 亮,孙孟瑶,王晶晶,李培玺,储炳银,徐小牛
(安徽农业大学林学与园林学院,安徽 合肥 230036)
森林土壤微生物是土壤生态系统中最活跃的组分之一,在促进土壤养分循环、物质转化和能量交换中起着重要作用[1],具有分解土壤矿物质、促进腐殖质的分解和合成、保护植物根系免受病原菌的侵染和其他特性的能力[2]。同时,微生物本身还含有一定的N、P、K等,为地上植物生长提供必要的养分,直接或间接影响着植物的生长[3]。土壤微生物不仅受地上植物和土壤特性的影响,还通过自身特性与植物相互作用,形成与植物的交互反馈机制[4]。大量研究表明,土壤微生物群落功能多样性不仅能敏感地指示气候和土壤环境状况的变化,还可以反映土壤质量和森林健康状况[5-6],被认为是重要的生态指标。因此,研究森林土壤微生物群落多样性可以揭示森林生态系统“植物-土壤-微生物”三者之间的相互作用机制[7]。
毛竹(Phyllostachys pubescens)是禾本科(Gramineae)常绿乔木状单子叶植物。毛竹作为我国亚热带地区广泛分布的大径级竹种,栽培历史悠久,其经营包括材用、笋用和笋材兼用,也可生态林经营,具有重要的经济和生态价值,在脱贫致富、乡村振兴中发挥着重要作用。由于毛竹特殊的繁殖方式,长期以来多以单一经营为主,易受自然干扰及市场风险的冲击,严重影响其经营效益[8]。同时,毛竹林结构单一,物种多样性低,存在一定的生态风险。因此,为提高竹林的经营效益及其生态稳定性,近年来竹木混交以及竹药、竹草复合经营备受关注[9],已成为生态研究的热点。相关研究表明,实施竹木混合经营有利于改善土壤养分状况和提高森林生产力,有利于毛竹林的可持续发展和管理[10-11]。但是,关于毛竹与药用植物复合经营的研究相对较少[9,12]。
白及(Bletilla striata)属兰科(Orchidaceae)多年生宿根草本、国家二级保护植物,其假鳞茎是我国传统、珍稀名贵中药材,具有收敛止血、消肿生肌的功能,主要用于治疗咯血、吐血、外伤出血、疮疡肿毒、皮肤皲裂等[13]。白及种植3~4年后即可收获假鳞茎,生长5年以上假鳞茎药用价值更高。由于需求增加,白及市场价格不断攀升,野生资源因乱采滥挖以及天然生境的破坏而濒临灭绝。因此,急需开发高效、绿色的白及林下仿野生栽培技术,促进白及中药农业可持续发展[9,14]。
目前,有关林下套种白及的试验研究已有一些报道[9,15],但是毛竹-白及复合经营的研究甚少。为此,基于长期定位试验,对不同套种年限的毛竹-白及复合林分土壤微生物群落功能多样性进行分析,旨在探讨土壤微生物、植被与土壤质量之间的相互作用关系,为毛竹-白及复合模式的可持续经营提供依据。
1 材料与方法
1.1 样地概况与样品采集
试验样地位于安徽广德市邱村镇(119°23′37″E,31°01′49″N),属皖南山地和沿江平原的过渡带,以丘陵岗地为主,地势较平坦,竹资源丰富。属北亚热带湿润季风气候,四季分明,温和湿润,年平均气温为16.3℃,无霜期近300 d;多年平均降水量达1294 mm,主要分布在夏秋季节。地带性植被为亚热带常绿阔叶林,并分布有大面积毛竹林、杉木(Cunninghamia lanceolata)、马 尾 松(Pinus massoniana)和湿地松(P. elliottii)等人工林。地带性土壤为红黄壤,质地较黏,土层深厚,肥力一般。
该地自2010年开始毛竹林下种植中药材,以白及、黄精为主,目前种植面积约150 hm2。套种前,通过疏伐降低竹林密度,使郁闭度在0.5~0.6,再进行林地垦复,深度约30 cm,种植前每公顷施用腐熟有机肥22.5 t。选用1年生白及组培苗,按株行距20 cm×30 cm栽植,栽后每年除草2~3次,没有追肥和使用农药。于2019年5月白及生长前期在试验区内进行样地调查与样品采集,不同复合林分的栽培管理措施一致,其中毛竹林为天然林,除适度采伐竹材外无任何经营措施。选取立地条件相近、套种白及年限分别为1、3、4年的毛竹复合经营林分,分别设置面积为20 m×20 m的标准样地各3个,样地间距不小于100 m,采用“S”形5点混合取样法,用土钻(内径3 cm)采取白及行间0~10 cm的土样,过筛去除凋落物、细根和石砾等,混合均匀后分为两份,一份装入无菌袋,置于便携式冰盒冷藏带回实验室,在4℃冰箱保存,用于土壤微生物功能多样性的测定;另一份风干后过2 mm筛,用于土壤理化性质测定。
1.2 土壤理化性质和土壤微生物群落功能多样性的测定
土壤理化性质指标采用常规方法测定[16]。土壤含水量采用烘干法测定;土壤pH按照水浸提电位法(水土比为2.5∶1),采用Extech Ⅱ型pH计测定;土壤有机碳和全氮使用元素分析仪(EA3000,Vector,Italy)测定;铵态氮、硝态氮使用2.0 mol·L-1KCl浸提、全磷经湿式消煮,随后采用自动流动分析仪(Alliance-Futura,France)测定。
土壤微生物群落功能多样性采用31种碳源的Biolog-ECO微平板法测定[17-18],即称约10 g新鲜土样放入250 mL的三角瓶中,加入100 mL已灭菌的0.85% NaCl溶液,摇床振荡30 min(转速180 r·min-1),取上清液稀释1000倍,再置于4℃冰箱内静置2~3 min,取其上清液倒入无菌培养皿,利用移液枪向ECO微平板每孔内加入150 μL稀释液,接种好的Biolog-ECO板于28℃生化培养箱中黑暗培养7 d,每隔24 h在Biolog微平板自动读取仪上测其吸光度。Biolog-ECO板上的31种碳源依其化学基团的性质,分成6大类,即糖类(10种)、羧酸类(7种)、氨基酸类(6种)、聚合物类(4种)、多胺类(2种)、多酚酸类(2种)。
1.3 数据处理与统计分析
通过Biolog-ECO微平板测定的平均每孔颜色变化率(Average well color development,AWCD)可表示微生物群落的整体代谢活性[18],是反映土壤微生物碳源利用能力及其活性和多样性大小的一个重要指标:
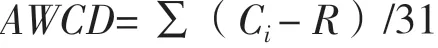
式中: Ci为第i个孔590 和750 nm的吸光度值,R为对照孔吸光度值,若Ci- R ≤ 0,其吸光度值计为0;31为ECO板供试碳源的种类数。
根据培养72 h的AWCD值变化,土壤微生物群落多样性指数按下式计算[18]:

式中:Pi=(Ci-R)/∑(Ci-R),表示第i孔的相对光密度值与整个微板相对光密度值总和的比值,ni表示第i孔的相对吸光度(Ci-R)。
利用Excel 2016整理数据,SPSS 19.0进行统计分析,采用单因素方差分析和LSD多重比较法对不同套种年限毛竹-白及复合林分土壤微生物碳源利用特征的差异显著性进行分析(P<0.05),Pearson相关系数检验各因子之间的相关性;采用Origin 2018 64bit绘图。
2 结果与分析
2.1 不同套种年限毛竹-白及复合林分的土壤理化性质
不同套种年限对复合林分土壤理化性质具有显著影响(表1)。套种4年的林分土壤含水量、全氮、全磷含量均显著高于套种1和3年的复合林分(P<0.05);套种3年的土壤有机碳、铵态氮显著低于套种1和4年的复合林分(P<0.05);套种1年的土壤pH显著低于套种3和4年的复合林分(P<0.01)。
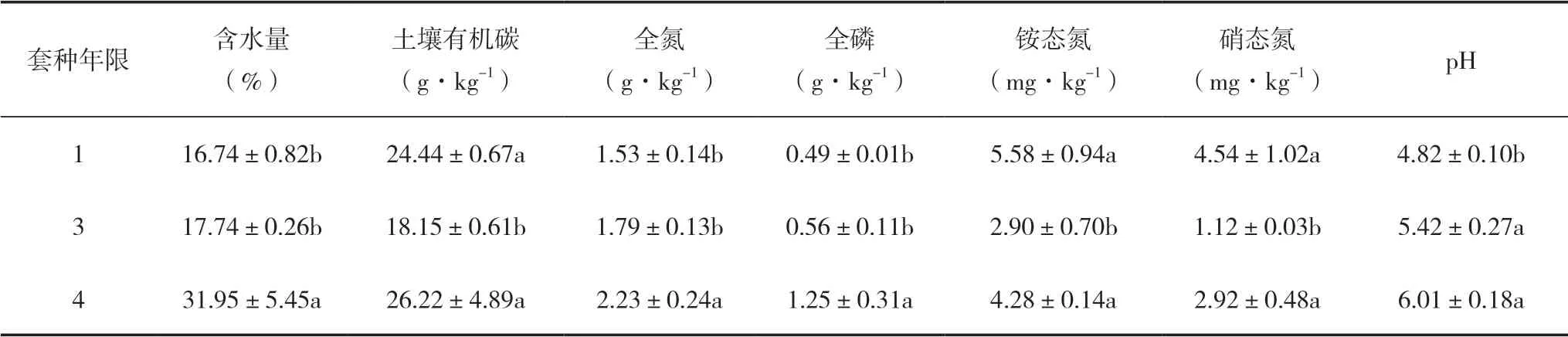
表1 毛竹-白及复合系统土壤理化性质
2.2 不同套种年限毛竹-白及复合林分土壤微生物群落碳源利用特征
试验结果显示,不同林分土壤微生物群落AWCD值随着培养时间的增加呈逐渐升高的趋势(图1)。在培养0~24 h内没有显著变化,随后呈直线上升,表明微生物代谢活性旺盛,碳源被大量利用。不同套种年限的土壤微生物利用不同碳源的AWCD值呈现4年>3年>1年的变化趋势。与套种1年的林分相比,套种3和4年的AWCD值均有显著差异(P<0.01);而套种1年的AWCD值无显著差异(P>0.05)。

图1 不同套种年限毛竹-白及复合林分土壤微生物群落平均颜色变化率
土壤微生物群落利用6大类碳源的AWCD值均随着培养时间的延长逐渐升高(图2),对6大类碳源的代谢能力差异显著(P<0.01)。在不同碳源利用水平上,不同套种年限林分差异较大(表2)。套种1年的土壤微生物群落碳源利用水平从高到低依次是羧酸类、氨基酸类、聚合物类、糖类、多胺类、多酚酸类,除糖类之外,均显著低于套种3和4年林分(P<0.05)。套种3和4年林分土壤微生物群落对碳源利用仅在多胺类存在显著差异(P<0.05),在其他碳源利用上两林分间无显著差异。

表2 培养72 h土壤微生物对碳源的利用程度
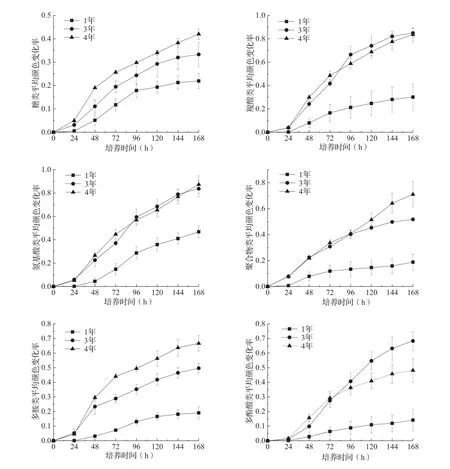
图2 不同套种年限毛竹-白及复合林分土壤微生物利用不同碳源的AWCD值
2.3 不同套种年限毛竹-白及复合林分的土壤微生物群落功能多样性
随着套种年限的增加,土壤微生物多样性、丰富度和优势度指数均表现为逐渐升高的趋势(表3)。不同套种年限的多样性指数和72 h AWCD值无显著差异(P>0.05);套种3和4年的土壤微生物丰富度和优势度指数显著高于套种1年林分(P<0.01)。可见,套种年限对土壤微生物群落物种丰富度和均匀度指数影响较大,但对微生物群落优势度指数影响较小。

表3 不同套种年限毛竹-白及复合模式土壤微生物群落多样性指数
主成分分析(PCA)结果显示,第一主成分(PC1)贡献率为52.39%,第二主成分(PC2)贡献率为14.94%,累计贡献率达67.33%。PC1和PC2作为微生物群落碳源利用变化的主要成分,可反映不同套种年限土壤微生物群落代谢能力的基本情况。不同套种年限的复合林分土壤微生物群落具有明显的分异(图3)。套种1年林分主要分布在PC1轴负方向,套种3年林分分布在PC2轴正方向,套种4年林分分布在PC1轴正方向;且不同套种年限林分的土壤样本分布较为分散,说明随着套种年限的增加,土壤微生物群落代谢功能变化明显。

图3 不同套种年限毛竹-白及复合模式土壤微生物群落多样性的主成分分析
由表4可知,与PC1具有较高相关性的碳源(| r | > 0.6)有18个,包括糖类3个、羧酸类6个、氨基酸类4个、聚合物类2个、多胺类2个和多酚酸类1个;与PC2具有较高相关性的碳源有5个,包括糖类2个、羧酸类1个和氨基酸类2个。表明6大类碳源对PC1和PC2均起分异作用,其中对PC1影响大。

表4 31种碳源在PC1和PC2上的载荷值(|r|>0.6)
2.4 土壤微生物多样性和土壤理化性质的相关性
从表5可以看出,土壤微生物McIntosh指数与土壤全磷、pH呈显著正相关(P<0.05);土壤微生物Shannon指数与土壤全磷呈显著正相关(P<0.05),与pH值呈极显著正相关(P<0.01);土壤微生物Simpson指数与土壤全磷呈显著正相关(P<0.05),与铵态氮和硝态氮呈显著负相关(P<0.05),与pH值呈极显著正相关(P<0.01)。这说明土壤微生物群落多样性指数与土壤理化性质密切相关、相互影响。

表5 土壤微生物群落多样性指数与土壤理化性质的相关系数
从表6中可以看出,6大类碳源中的糖类和多酚酸类与pH呈显著正相关(P<0.05);羧酸类和聚合物类与全磷呈显著正相关(P<0.05),与pH呈极显著正相关(P<0.01);氨基酸类与pH呈极显著正相关(P<0.01);多胺类与含水量呈显著正相关(P<0.05),与土壤有机碳、全氮、全磷以及pH均呈极显著正相关(P<0.01)。这说明土壤微生物群落与土壤理化性质关系紧密,尤其是多胺类与理化性质之间的关系显著。
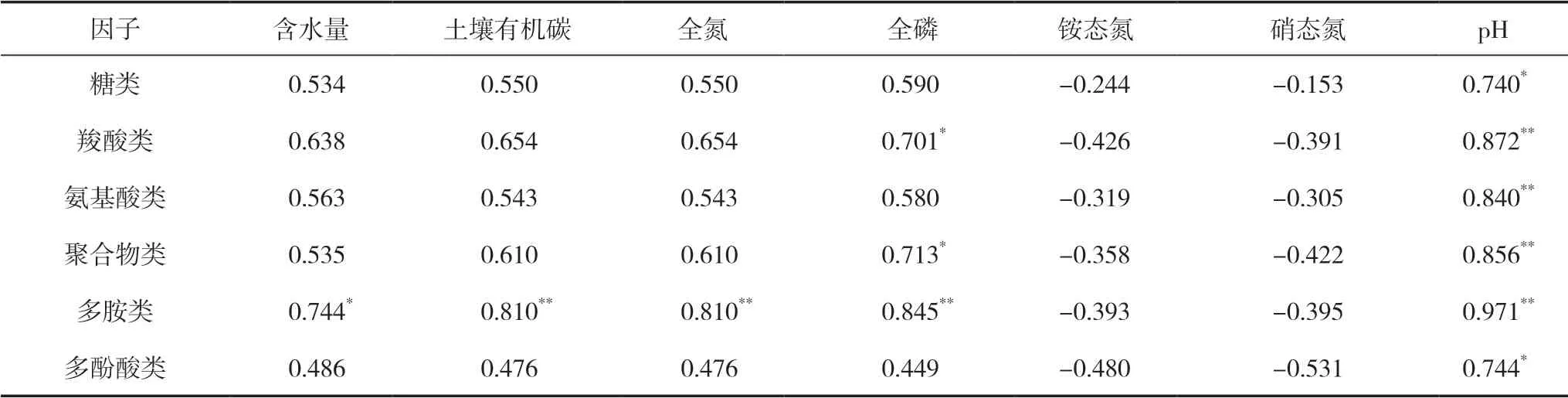
表6 土壤微生物碳源利用强度与土壤理化性质的相关系数
3 讨论与结论
3.1 套种年限对土壤微生物群落碳源利用特征和功能多样性的影响
土壤环境是土壤微生物组成和差异的主要因素[19],而森林凋落物、根系分泌物和土壤有机质是土壤微生物可利用碳源的主要来源[20-22]。有研究表明通过合理的间作套种以及不同树种混交,可提高土地利用率,改善土壤环境,增加土壤微生物多样性[23]。本研究发现,不同套种年限的毛竹-白及复合林分土壤微生物群落AWCD值变化存在明显差异,随着套种年限增加,土壤微生物碳源利用强度显著提高,套种4年林分土壤微生物代谢活性最强,而套种1年林分土壤相对较弱。这可能与白及生长发育有关,套种初期(1年)白及个体小,覆盖度低,随着年限增加,白及覆盖度不断提高,套种4年时达85%以上,土壤生态系统逐渐趋于稳定,为微生物群落发育提供了适宜条件;同时,白及地上部生物量随生长年限增加逐渐增大,凋落物量以及根系分泌物增多,为微生物群落提供了充裕的可利用碳源种类和数量,促进了微生物的生长发育[24-25]。因此,微生物群落多样性显著提高,代谢功能增强。王超群等[26]研究显示,随着林龄的增加,杉木林土壤微生物群落碳源利用率逐渐提高,这与本研究结果一致。
多样性指数反映了土壤微生物群落功能多样性变化,不同指数反映多样性不同侧面,Shannon指数和McIntosh指数主要反映群落物种个体数和分布均匀度,而Simpson指数反映群落中最常见类群的优势度[27-28]。本研究中Shannon指数、McIntosh指数和AWCD值均随套种年限增加显著提高,表明土壤微生物功能多样性随年限增加而提高。但Simpson指数在不同套种年限之间差异不显著,说明土壤微生物群落的优势种群并没有随套种年限增加而显著变化。由于土壤微生物群落结构及其多样性不仅受到立地环境的影响,而且与植被类型及其组成结构紧密相关[29],在立地环境基本一致的条件下,则主要受到植物群落结构的影响。Merilä等[30]研究得出,土壤微生物群落多样性与林下物种丰富度及多样性呈正相关关系。由于本研究的试验地立地条件较一致,因此,不同套种年限林分结构变化可能是导致土壤微生物群落功能多样性差异的主要因素。
3.2 套种年限对土壤理化性质的影响
根据不同类型碳源利用微生物随套种年限变化的差异(图2)可知,糖类碳源利用微生物受套种年限影响程度比较小,可能是毛竹-白及复合林分土壤的优势微生物类群。但是,随着套种年限增加,在套种3年后氨基酸类、羧酸类、糖类、聚合物类等碳源利用微生物成为优势类群。主成分分析结果直观反映了不同套种年限毛竹-白及复合林分土壤微生物群落利用碳源的种类和强度的整体差异[31]。套种年限的变化使得植被覆盖度发生变化,由此引发土壤环境改变,从而导致土壤微生物群落结构变化。本研究结果表明,表层土壤中有机碳、全磷、pH随着复合年限增加逐渐增加,与Chatterjee等[32]研究得出的土壤pH随着林龄的增加而降低的结果不一致。这可能是由于复合林分中白及生长逐年加快,对土壤养分需求较大,导致土壤中氮素的转化加速,有效氮增加,根系在吸收硝态氮时分泌出OH-来维系细胞膜内外电荷平衡,最终通过土壤中的OH-来协调土壤的酸碱度[33]。同时,白及生长加快,地上部生物量逐年增加,大量吸收土壤下层养分,地上部枯死后归还土壤增多,在一定程度上加速了土壤养分循环速率。相关分析结果表明,土壤微生物多样性指数与土壤全磷、pH、铵态氮和硝态氮具有显著相关性,土壤微生物碳源利用强度与土壤pH也具有显著相关性,与王淼等[34]研究结果相似。可见,土壤微生物碳源利用特征及多样性指数受到诸多因素的影响,加之Biolog-ECO微平板法的局限性[18],如要全面认识植物-土壤-微生物之间的互作机制,今后须结合高通量测序等技术,深入探讨竹药复合模式土壤微生物群落多样性的演变规律。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
中国农业科技导报(2022年7期)2022-09-27
江苏农业科学(2022年6期)2022-04-15
铀矿冶(2022年1期)2022-02-17
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
安徽农学通报(2017年6期)2017-04-11
绿色科技(2017年3期)2017-03-14
安徽农学通报(2014年18期)2014-10-23
